

Abstract
The development of navigational strategies to solve spatial problems appears to be dependent on an intact hippocampal formation. The circular water maze task requires the animal to use extramaze spatial cues to locate a pedestal positioned just below the surface of the water. Presently, we investigated the role of a recently discovered brain angiotensin receptor subtype (AT4) in the acquisition of this spatial learning task. The AT4 receptor subtype is activated by angiotensin IV (AngIV) rather than angiotensins II or III, as documented for the AT1 and AT2receptor subtypes, and is heavily distributed in the CA1–CA3 fields of the hippocampus. Chronic intracerebroventricular infusion of a newly synthesized AT4 agonist (Norleucine1-AngIV) via osmotic pump facilitated the rate of acquisition to solve this task, whereas treatment with an AT4 receptor antagonist (Divalinal) significantly interfered with the acquisition of successful search strategies. Animals prepared with bilateral knife cuts of the perforant path, a major afferent hippocampal fiber bundle originating in the entorhinal cortex, displayed deficits in solving this task. This performance deficit could be reversed with acute intracerebroventricular infusion of a second AT4 receptor agonist (Norleucinal). These results suggest that the brain AngIV–AT4 system plays a role in the formation of spatial search strategies and memories. Further, application of an AT4 receptor agonist compensated for spatial memory deficits in performance accompanying perforant path knife cuts. Possible mechanisms underlying this compensatory effect are discussed.
- spatial memory
- hippocampus
- perforant path knife cuts
- angiotensin IV analogs
- AT4 receptor
- circular water maze
Several classic roles have been ascribed to the brain renin angiotensin system, including blood pressure regulation, body fluid homeostasis, cyclicity of reproductive hormones and sexual behavior, and regulation of pituitary hormones (for review, see Johnston, 1990; Saavedra, 1992; Wright and Harding, 1992;Fitzsimons, 1998). These functions are mediated by the angiotensin receptor subtype AT1, with less involvement by the AT2 subtype. Both of these receptor subtypes are activated by the octapeptide angiotensin II (AngII) and the heptapeptide angiotensin III (AngIII) (for review, see Smith, 1996;Wright and Harding, 1997). Recently, our laboratory discovered and characterized a third angiotensin binding site (Harding et al., 1992;Swanson et al., 1992; Zhang et al., 1999), designated AT4(de Gasparo et al., 1995). This receptor subtype is activated by the hexapeptide angiotensin IV (AngIV) and is prominent in brain structures important to cognitive processing and sensorimotor functions, including neocortex, hippocampus, dentate gyrus, thalamus, and cerebellum (Miller-Wing et al., 1993; Wright et al., 1995). The AT4subtype is also present in the nucleus basalis magnocellularis (NBM) and medial septum in high densities (Møeller et al., 1996). In contrast, AT1 and AT2 subtypes are poorly represented in the neocortex, hippocampus, dentate gyrus, and cerebellum (Wright et al., 1995; Wright and Harding, 1997), although a recent immunohistochemical study has reported AT1-positive immunoreactivity in the dentate gyrus and subiculum, with lesser staining in the CA3 field and few stained cells observed in CA1 and CA2 fields (von Bohlen und Halbach and Albrecht, 1998). Intracerebroventricular injection of AngIV has been shown to stimulate c-fos expression in the CA1–CA3 fields of the hippocampus (Roberts et al., 1995) and to facilitate passive-avoidance conditioning in rats (Braszko et al., 1988; Wright et al., 1993). These results suggest that intracerebroventricular delivery of this peptide influences the hippocampus.
The notion that the hippocampus plays an important role in spatial memory processing is supported by the observation that damage to the hippocampus results in impaired ability to solve tasks that rely on spatial search strategies (Olton et al., 1978; Morris et al., 1990;Sutherland and McDonald, 1990). Thus, hippocampal damage has been correlated with disruption of spatial memory in a number of mammalian species, including rat (Morris et al., 1982; Sutherland et al., 1982,1983; Nadel, 1991; Jarrard, 1993) and human (Volpe and Hirst, 1983;Cummings et al., 1984; Zola-Morgan et al., 1986). Bilateral hippocampal lesions have also been shown to interfere with acquisition and/or retention of spatial memory in rats as measured by performance on the circular water maze task (Morris et al., 1982; Rudy and Sutherland, 1989; Stubley-Weatherly, 1996).
Our laboratory has synthesized and characterized several AT4-specific agonists, including Norleucine1-AngIV (Nle1-AngIV), Norleucinal (Sardinia et al., 1994; Wright et al., 1995), and AT4 receptor antagonists, including Divalinal (Krebs et al., 1996). The present investigation evaluated the efficacy of Nle1-AngIV to facilitate the acquisition of the circular water maze task and of Divalinal to disrupt acquisition. We also determined whether Norleucinal treatment compensated for deficits in spatial memory produced by damage to the perforant path (PP). Norleucinal was used rather than Nle1-AngIV because of its added resistance to degradation. The PP is the major afferent pathway to the hippocampus projecting from the entorhinal cortex to the dentate gyrus, passing near the dorsal hippocampal commissure (Hjorth-Simonsen and Jeune, 1972; Skelton and McNamara, 1992; Klug et al., 1998).
MATERIALS AND METHODS
Subjects
Male Sprague Dawley rats (270–350 gm, Charles River-derived) were adapted to a 12 hr light/dark cycle initiated at 7:00 A.M. in an AAALAC (Association for Assessment and Accreditation of Laboratory Animal Care) approved vivarium at a temperature of 21 ± 1°C. The animals were housed singly and were permitted access to water and Purina laboratory rat chow ad libitum; however, food was removed the night before surgery.
Apparatus
Motor test battery. All animals were tested for potential treatment-induced disturbance in motor ability using a motor test battery developed by Combs and D’Alecy (1987). The first test of this battery required that each animal be placed on a horizontal screen (62 × 54 cm, grid size of 0.6 × 0.6 cm) that was rotated into the vertical plane with a vertical drop from the screen to towels of 1.2 m. The duration in seconds that the animal was capable of holding onto the screen was recorded up to a maximum of 15 sec. The animal received one point for 0–5 sec, two points for 6–10 sec, and three points for 11–15 sec. After a 20 min rest period, the animal was next placed on a horizontal wooden dowel (3 cm in diameter × 62 cm long), also elevated 1.2 m above the towels. The time that the animal balanced on the rod was measured up to 15 sec. The same scoring procedure was used as described above. The third test consisted of timing the duration that the animal could cling to a horizontal hemp rope (1.0 cm diameter) using its forearms for up to a maximum of 5 sec. The vertical drop onto towels was 1.0 m. Points were awarded as follows: one point for 0–2 sec, two points for 3–4 sec, and three points for >4 sec. Each animal could achieve a maximum cumulative score across the three tasks of nine points.
Circular water maze. The circular water maze task developed by Morris (1981, 1984) was used to test spatial learning ability. The water maze consisted of a 1.6 m diameter × 0.6 m tall galvanized cylindrical tank painted black and filled to a depth of 30 cm with 26–28°C water. Three walls of the test room (40 cm from the edge of the tank) were prepared with visual cues consisting of circles on one wall, triangles on a second wall, and squares on a third wall. The fourth wall was 1.2 m from the edge of the tank, thus permitting space for one experimenter to stand. The position of this experimenter was constant after placement of the animal into the maze. A commercially available video tracking system and accompanying software package (Chromatrak; San Diego Instruments, San Diego, CA) was used to measure each animal’s latency and path distance to find the submerged pedestal.
The animals used in the first two experiments concerned with chronic treatment of Nle1-AngIV and Divalinal were prepared with osmotic pumps and, after 24 hr of recovery acquisition training was initiated consisting of five trials per day for 6 d. At the completion of this phase, one probe trial (description follows) was conducted, each osmotic pump was then removed under local anesthesia (Lidocaine; Elkins-Sinn, Cherry Hill, NJ), and the wound was closed with skin staples (model 8024–12; Davis and Geck, American Cyanamid, Wayne, NJ). After 1 d of recovery, 6 additional training days were conducted to determine whether treatment-induced deviations in acquisition could be overcome after termination of drug delivery. During these additional training days, the submerged pedestal was positioned in the opposite quadrant to the initial placement for each rat. At the completion of this phase, one additional probe trial was conducted.
Members of a third experiment were prepared with bilateral perforant path knife cuts and similarly tested; however, no osmotic pumps were implanted, rather acute intracerebroventricular injections were administered as described below. These animals received 8 d of acquisition training, five trials per day, with one probe trial conducted at the completion of testing on day 8.
Each trial entailed placing the animal into the water facing the wall of the pool at one of four locations [north (N), south (S), east (E), and west (W)] and tracking its swimming path and duration until it found the round submerged pedestal (12 cm diameter painted black, 3 cm below the surface). The pedestal was placed 30 cm from the edge of the tank equidistant from the edge of the quadrant within one of the four quadrants: NW, NE, SW, and SE. If the animal located and mounted the pedestal, it was permitted 30 sec on the pedestal before the next trial commenced. If the animal did not find the pedestal within 120 sec, it was placed directly on the pedestal and allowed a 30 sec rest period. The animal’s entry point was randomized on each trial, although the location of the pedestal was initially randomly assigned but remained fixed for each animal during acquisition training, and was repositioned as described above after pump removal for members of experiments 1 and 2. At the end of each test day, the animal was dried off with a towel and placed under a 100 W lamp for 10 min before being returned to its home cage. After the fifth trial of days 6 and 13, the pedestal was removed for members of the first two experiments, and a 2 min probe trial was completed. The animals of the third experiment were administered one probe trial at the conclusion of testing on day 8. During these probe trials, the time spent within the quadrant, as well as the number of crossings into and out of the quadrant where the pedestal had been located, were recorded.
Compounds. Table 1 provides information on the structures of the compounds, their molecular weights, peptide content as determined by HPLC analyses, AT4 receptor binding affinity, and the source, i.e., synthesized in our laboratory or provided by a commercial supplier. An automated peptide synthesizer (Coupler 250; DuPont Wilmington, DE) was used to prepare peptides not commercially available. Angiotensin IV is included for reference only. Peptide purity ranged from 90–100%, whereas acetate represented the major contributor to the decreased peptide content. Corrections were made for differences in peptide content and purity when the compounds were prepared for use.
Structure, molecular weight, peptide content, AT4 binding affinity, and source of each peptide examined
Design and procedures
Nle1-AngIV treatment. Twenty-four rats were randomly assigned to one of three treatment groups (n = 6 each) or an artificial CSF (aCSF) control group (n = 6). Each rat was anesthetized with ketamine hydrochloride (100 mg/kg, i.m.; Bristol-Myers, Syracuse, NY) and xylazine (2 mg/kg, i.m.; Haver-Mobey, Shawnee, KS) and prepared with a 7 d osmotic pump (model 2001; Alza Scientific Products, Palo Alto, CA) that infused intracerebroventricularly at the rate of 1 μl/hr. This was accomplished via a stereotaxically positioned length of hypodermic stainless steel tubing (23 ga, length of 3.2 cm) prepared with a 90° bend such that a 7 mm length of the tubing was inserted through a skull trephine hole, thus penetrating the roof of the lateral ventricle. Flat-skull coordinates used for placement of the trephine hole were 1.0 mm posterior (P) to bregma and 1.5 mm lateral (L) to midline (Paxinos and Watson, 1986). The stainless steel tubing was anchored to the cranium with stainless steel screws and dental cement. The pump was connected to the stainless steel tubing via PE-60 tubing (Clay Adams, Parsippany, NJ) and placed subcutaneously between the scapulas. These animals were given 1 d to recover and then testing was initiated, first on the motor function test battery and then daily in the circular water maze.
Members of groups 1, 2, and 3 received 0, 0.1, and 0.5 nmol/hr, respectively, the AT4 receptor agonist Nle1- AngIV. The 0 nmol/hr group represented an aCSF infusion control group. These doses of Nle1-AngIV were established in a preliminary investigation that noted a significant improvement in acquisition with intracerebroventricular administration of 0.1 and 0.5 nmol/hr compared with controls but no additional facilitation of acquisition when comparing the 0.5 and 1.0 nmol/hr doses. Members of group 4 received the pentapeptide AngII(4–8) (catalog #7037; Peninsula Laboratories, Belmont, CA) at a dose of 0.5 nmol/hr. Angiotensin II(4–8) has been shown to bind with low affinity at the AT4 receptor site (Sardinia et al., 1993) (Table 1). After the completion of day 6 of acquisition trials, each animal was tested for strength of conditioning using a probe trial. After a 2 hr rest period, each animal was further evaluated for sensorimotor deficits by using a visible pedestal (2 cm above the surface of the water). Both the entry points (N, S, E, W) and locations of the visible pedestal (NW, NE, SW, SE) were randomly assigned for each rat on each of five trials. In all other respects, these trials were conducted as described above. The osmotic pumps were removed on day 7, and acquisition trials were resumed beginning on day 8 for an additional 6 d, with the pedestal repositioned to the opposite quadrant for each animal. On day 13, an additional probe trial was conducted.
Divalinal treatment. Twenty-four rats were randomly assigned to a nontreated control group (n = 6) or one of three treatment groups (n = 6 each): 0, 0.5, or 5.0 nmol/hr Divalinal. These doses of Divalinal were determined in a preliminary study that noted no differences in inhibition of acquisition when comparing 5.0 and 10.0 nmol/hr. Each member of the treatment groups was prepared with a 7 d osmotic pump as described above. Training trials, probe trials, and the visible pedestal protocols were identical with the procedures described in the first experiment.
Perforant path knife cuts. Thirty-two rats were randomly divided between two major groups (16 rats each) and were prepared as follows during one surgical session under ketamine hydrochloride and xylazine. Members of the first group received bilateral PP knife cuts directed at the medial and lateral perforant path tracts that extend from the entorhinal cortex to the dentate gyrus, according to the protocol offered by Skelton and McNamara (1992). These knife cuts were accomplished in two steps using a stereotaxically held knife blade (width of 7 mm, thickness of 0.2 mm; Fine Science Tools, Foster City, CA) with flat-skull coordinates relative to bregma of P, 8.0; L, 3.5; and V, 6.0 mm from dura. From this starting point, the knife was moved medially 0.5 mm. The blade was removed and repositioned according to the following coordinates: P, 8.0; L, 4.8 and V, 6.5 mm from dura. From this starting point, the blade was again moved medially 0.5 mm and withdrawn. This procedure was then repeated in the opposite hemisphere. The animals of the second group served as surgical controls and received equivalent knife cuts of the occipital cortex directly superior to the location of the perforant path cuts made in members of the first group. These neocortex knife cuts were made according to the same posterior and lateral coordinates; however, the blade was lowered only 3 mm V to dura. Each animal was also prepared with an intracerebroventricular guide cannula (PE-60) stereotaxically positioned above the right lateral ventricle and fastened in place with skull screws and dental cement. This procedure is similar to that described above and has been reported previously in detail (Wright et al., 1985).
After 7 d of recovery, each animal was behaviorally tested for correct cannula placement by the intracerebroventricular injection of AngII (10 pmol in 2 μl of aCSF). This was accomplished by inserting a preloaded 30 ga stainless steel hypodermic tubing injector, prepared with a 24 ga stainless steel tubing sleeve, into the guide cannula such that it extended 2 mm beyond the tip of the guide, thus penetrating the roof of the lateral ventricle. Angiotensin II was then hand delivered via a 10 μl Hamilton syringe over a 30 sec period. The guide cannula was considered to be correctly placed if a burst of drinking was elicited within 5 min after AngII injection. After an additional 2 d of recovery, the animals were divided into subgroups (n = 8 rats each), which were treated with intracerebroventricular bolus injections of Norleucinal (1.0 nmol in 2.5 μl aCSF) or aCSF (2.5 μl) 5 min before testing for motor dysfunction using the motor function test battery. This was followed by 8 d of acquisition training in the circular water maze task, also preceded by the intracerebroventricular injection of Norleucinal or aCSF 5 min before training. At the completion of testing on the final day, a probe trial was conducted for each animal as described above.
Histology
Correct placement of the intracerebroventricular guide cannula for each animal used in the first two experiments was confirmed by the intracerebroventricular injection of 10–12 μl of fast green dye via the chronic cannula under equithesin anesthesia (3.5 ml/kg, i.p.; Jensen-Salsbury Laboratory, Kansas City, MO), followed by brain extraction and visual confirmation of dye within the brain ventricles. Each osmotic pump was also checked to see whether its contents had been exhausted.
After behavioral testing, each animal used in the third experiment was deeply anesthetized with equithesin and intracardially perfused with PBS, followed by 10% paraformaldehyde. The brains were removed and stored in 10% formaline and, 48 hr before sectioning, the brains were transferred to a 10% formaline–20% sucrose solution. Each brain was sectioned at 14 μm in the horizontal plane using a cryostat (Jung frigocut 2800E; Leica Instruments, Nussloch, Germany). Every third section through the knife cut was mounted on electrostatic microscope slides (Fisher Scientific, Pittsburgh, PA) and stained via a modified hematoxylin and eosin staining technique for verification of damage. The modified staining procedure consisted of the following sequence: (1) Gill-2 hematoxylin (2 min); (2) dH2O (10 dips); (3) acid rinse [2 ml of 0.2% HCl–1 dH2O (10 dips)]; (4) dH20 (10 dips); (5) bluing reagent [2 ml of 30% NH4(OH)–1 dH20 (1 min)]; (6) dH2O × 2 (10 dips each); (7) 95% EtOH × 2 (10 dips each); (8) eosin-Y (1 min); (9) 95% EtOH (10 dips); (10) 95% EtOH (10 dips); (11) 100% EtOH × 3 (10 dips each); and (12) xylene × 3 (10 dips each).
Knife cut damage was quantified by a computerized scanning device (Envisions Scanner, Trans env. 24 pro; International Business Machines, White Plains, NY) connected to an IBM personal computer formatted with appropriate software (Adobe Photoshop, Adobe Systems, San Jose, CA). After each initial scan, the area of damage was calculated via an additional computer package (SigmaScan Image; Statistical Program for the Social Sciences, Chicago, IL) that provided lesion size, as well as overall structural region size, in square millimeters. Reconstruction of the lesion along the dorsoventral plane permitted calculation of lesion volume in cubic millimeters. The knife cut damage was then converted to percent of total perforant path volume for each hemisphere. Hjorth-Simonsen and Jeune (1972) andPaxinos and Watson (1986) were consulted regarding identification of structures damaged.
Data analyses
The data sets concerned with the total score for each animal on the motor function test battery were evaluated using a one-way ANOVA. Significant effects were further evaluated using Newman–Keuls post hoc tests with a level of significance set at p = 0.01.
The mean latency and path distance to find the submerged pedestal during each daily block of five trials was calculated for each animal for each day of acquisition. Because we anticipated group differences only during early training trials in the first experiment, these data were submitted to a priori established separate one-way ANOVAs for days 1, 6, and 13 of testing. Given the use of an AT4 receptor antagonist in experiment 2, we predicted significant interference with acquisition during the entire 6 d treatment period. Therefore, these data were submitted to a groups × days ANOVA, with repeated measures on the second factor. Once again,a priori established one-way ANOVAs were applied to each data set concerned with latency and path distance on days 1, 6, and 13 of acquisition. We reasoned that, by the final day of acquisition, performance would be reasonably stable, thus permitting meaningful group comparisons regarding asymptotic levels. Similarly, one-way ANOVAs were used to determine whether there were differences among groups during probe trials and during the visible pedestal protocol. In the third experiment, we expected group differences to persist over all 8 d of acquisition training. Therefore, these data were submitted to a groups × days ANOVA, with repeated measures on the second factor. A one-way ANOVA was used to test for group differences during probe trials.
RESULTS
Histological findings
All animals of the first two experiments were found to have correctly placed intracerebroventricular guide cannulas, as evidenced by the presence of dye within the lateral ventricles. Each osmotic pump was also found to be either empty or nearly empty. The animals of the third experiment that received PP knife cuts and were treated with Norleucinal exhibited an overall mean ± SEM of 66.7 ± 3.2% damage to this pathway (left hemisphere, 67.0%; right hemisphere, 66.3%). Those animals prepared with PP knife cuts and treated with aCSF revealed an overall mean ± SEM of 64.4 ± 4.2% damage (left hemisphere, 65.3%; right hemisphere, 63.5%). Representative photomicrographs are presented from one animal of the PP path knife cut group treated with Norleucinal (Fig.1A) and one animal from the neocortex knife cut group treated with Norleucinal (Fig.1B). These knife cuts were discrete and transsected the PP at the anteroposterior level of the dorsal hippocampal commissure and subiculum, thus isolating the entorhinal cortex from more anterior brain structures. In all rats, the knife cuts passed through the forceps major of the corpus callosum, and the dorsoventral extent of damage generally spanned the full range of entorhinal cortices. There was no discernable damage noted in the dentate gyrus or CA1–CA3 fields of the hippocampus. The knife cut control rats evidenced damage to the neocortex overlying the PP–entorhinal cortex region; however, these latter structures remained intact. Members of the neocortex knife cut control groups that received Norleucinal or aCSF revealed no loss of PP fibers.
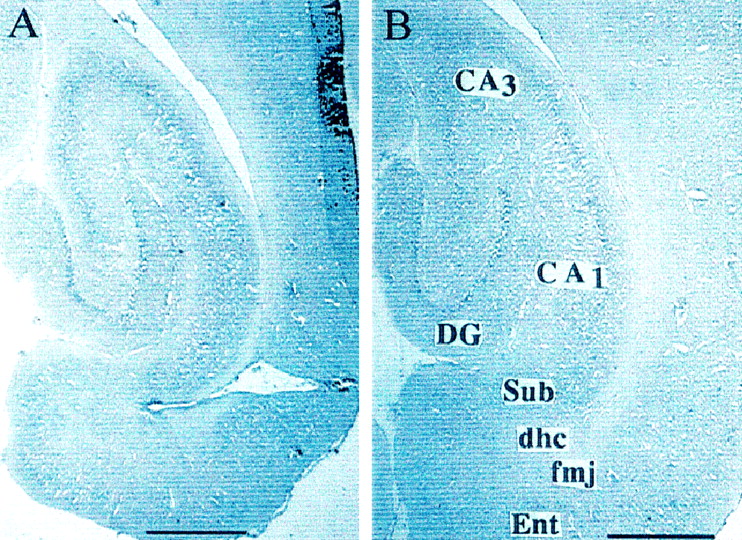
Representative photomicrographs of perforant path knife cuts (A) and control knife cuts to the neocortex (B) from animals treated with Norleucinal. Horizontal sections of the right hemisphere were taken at the level of 4.1 mm ventral to bregma according to Paxinos and Watson (1986). These knife cuts transsected the perforant pathway at the anteroposterior level of the dorsal hippocampus commissure (dhc), forceps major of the corpus collosum (fmj), and subiculum (Sub). These cuts isolated the entorhinal cortex (Ent) from the dentate gyrus (DG) and other anterior brain structures, such as the CA1–CA3 fields. Scale bar, 1 mm.
Motor and sensory functions
There were no differences in cumulative scores across the three motor tasks comparing the groups of the first experiment (F(3,20) = 2.78; p > 0.05). The mean ± SEM scores for the groups treated with 0.5, 0.1, and 0 of nmol Nle1-AngIV, and 0.5 nmol of pentapeptide were 8.4 ± 0.2, 8.2 ± 0.3, 7.3 ± 0.4, and 8.4 ± 0.2, respectively. There were also no differences among the groups used in the second experiment (F(3,20) = 1.84;p > 0.10). The mean ± SEM scores for the groups treated with 5.0, 0.5, and 0 nmol/hr Divalinal and nontreated controls were 7.0 ± 0.2, 6.3 ± 0.3, 6.3 ± 0.3, and 7.0 ± 0.2, respectively. Comparable scores from the third experiment were as follows: PP cut group, 6.9 ± 0.5; cortex control knife cut group, 7.4 ± 0.4. These groups did not differ (F(1,30) = 1.12; p > 0.10). Thus, there was no evidence of motor dysfunction among any of the animals used in these experiments.
Similar results were noted for the visible pedestal trials. Specifically, no differences among the groups were measured in the first experiment with respect to latency to find the visible platform (F(3,20) = 1.10; p > 0.10) or distance swam to locate the pedestal (F(3,20) = 0.87; p > 0.10). Similar findings were noted for the groups of the second and third experiments, i.e., no differences concerning latency or distance swam to find the visible pedestal.
Circular water maze
Nle1-AngIV treatment
Figure 2A presents the mean ± SEM latency to find the submerged pedestal for each group during the initial 6 d of testing with the osmotic pumps in place and during the subsequent 6 d after pump removal. Figure2B presents the mean ± SEM path distances swam by members of each group. Overall, the results indicate that those animals treated with Nle1-AngIV performed better on the initial 2 d of acquisition, with respect to latency and distance swam to find the pedestal, than members of the control group infused with aCSF or those rats infused with the pentapeptide. Figure3 provides representative examples of performance by an animal treated with 0.5 nmol/hr Nle1-AngIV on days 1 and 6 of acquisition compared with an animal infused with 0.5 nmol/hr pentapeptide. The Nle1-AngIV-treated animal evidenced a superior search strategy on day 1 compared with the rat infused with pentapeptide. Generally, members of the Nle1-AngIV-treated groups appeared to make use of extramaze spatial cues to locate the pedestal earlier in their acquisition trials compared with members of the pentapeptide- and aCSF-infused groups. Thus, their search patterns progressed to more productive strategies sooner than members of the other two groups. However, by day 6 of acquisition, the performances of these animals were equivalent. After pump removal, members of all four groups performed nearly equivalently, although the pentapeptide-treated rats evidenced slightly poorer performance than members of the other groups on days 9–11.

Mean ± SEM group changes in latencies (A) and swim distances (B) to locate the submerged pedestal in a circular water maze task during 6 acquisition days with osmotic pumps in place, followed by 6 additional acquisition days with the pumps removed. All treatments were intracerebroventricularly delivered via osmotic pump at the indicated doses in a volume of 1 μl aCSF/hr. Those animals treated with 0.5 nmol of Nle1-AngIV performed better during days 1 and 2 of acquisition with respect to latency to find the pedestal (p < 0.005) and swim distance (p < 0.05) than members of the control group (0.0 nmol of Nle1-AngIV) or those rats infused with AngII(4–8) (Penta). Pentapeptide binds at the AT4 receptor with low affinity. These groups did not differ during days 3–6 of acquisition. After pump removal, the location of the submerged pedestal was shifted to the opposite quadrant for each animal. Although those animals treated with pentapeptide revealed longer swim distances to find the pedestal on days 9–11, by days 12 and 13, the groups did not differ. Each group consisted of six rats surgically prepared with a 7 d osmotic pump and were given 1 d to recover before the initiation of acquisition trials. Five trials were administered per day with entry points randomly assigned (N, S, E, W), although the location of the submerged pedestal was fixed for each rat.

Representative examples of search patterns in the circular water maze during a 2 min trial by one member of the group treated intracerebroventricularly with 0.5 nmol/hr Nle1-AngIV during days 1 (A) and 6 (B) of acquisition training and one member of the group treated with 0.5 nmol/hr pentapeptide during days 1 (C) and 6 (D) of acquisition. Latency (Lat) in seconds to find the submerged pedestal and distance swam (Dist) in meters are indicated for each animal. Those rats treated with Nle1-AngIV displayed a superior search strategy compared with animals treated with AngII(4–8) (Pentapeptide) on day 1 of acquisition, as evidenced by significantly shorter latencies (p < 0.05) and swim distances (p < 0.05) to find the submerged pedestal. By day 6, all animals had acquired efficient search strategies and did not differ. Each group consisted of six rats prepared with 7 d osmotic pumps that infused at a rate of 1 μl/hr aCSF.
The statistical analyses to support these conclusions indicated a significant groups effect concerning latency to find the pedestal on day 1 (F(3,20) = 6.08; p < 0.005). Post hoc analyses revealed that those rats infused with 0.5 nmol/hr Nle1-AngIV revealed significantly shorter latencies to find the pedestal than the aCSF-infused rats or those rats treated with pentapeptide. The animals infused with 0.1 nmol/hr Nle1-AngIV displayed shorter latencies than the aCSF-infused animals but were not different from the pentapeptide group or those rats infused with 0.5 nmol of Nle1-AngIV. A similar pattern emerged for path distance, with an overall difference among groups on day 1 of acquisition (F(3,20) = 3.18; p< 0.05). Both groups treated with Nle1-AngIV evidenced shorter path distances than those animals infused with aCSF or pentapeptide. By day 6 of acquisition, no differences among groups were seen concerning latency or path distance to locate the pedestal. This was also true at day 13 after pump removal.
The results of probe trials conducted on day 6 indicated no differences among the groups concerning time spent in the target quadrant (F(3,20) = 1.58; p > 0.10) or number of entries into the target quadrant (F(3,20) = 0.96; p > 0.10).
Divalinal treatment
Figure 4A presents the mean latencies to find the pedestal for each group of the second experiment. On the first day of training, the groups did not differ; however, across the subsequent 5 d of acquisition, there were differences among the groups. Those animals that were continuously infused with aCSF, or nontreated controls, revealed steady improvement in performance, whereas members of the two groups treated with Divalinal performed very poorly and required significantly longer latencies to find the pedestal. The groups × days interaction was also significant and indicated that the groups treated with Divalinal were different from the groups infused with aCSF and the control group on days 4–6. Post hoc analyses indicated that the rate of improvement in acquisition performance was substantially greater for those animals that received aCSF and the noninfused controls compared with the Divalinal-treated groups. Figure5 offers representative examples of performance by an animal from each of the four groups on day 6 of acquisition. Those rats treated with 5.0 or 0.5 nmol/hr Divalinal (Fig.5A,B, respectively) evidenced much poorer search strategies than those animals infused with aCSF (Fig.5C) or the noninfused controls (Fig. 5D). Several members of the 5.0 nmol/hr Divalinal group displayed a persistent tendency to swim near the walls of the maze (positive thigmotaxis) during training trials. This was seldom observed in the control animals or those infused with aCSF.

Mean ± SEM group changes in latency (A) and swim distance (B) to find the submerged pedestal in a circular water maze task during 6 d of acquisition training with osmotic pumps in place and 6 additional acquisition days with the pumps removed. All treatments were intracerebroventricularly delivered via osmotic pump at the indicated doses in a volume of 1 μl/hr aCSF. Those animals treated with 5.0 and 0.5 nmol/hr Divalinal (Dival) revealed significant deficits in performance compared with members of the control group (0 nmol Dival) and nontreated controls on days 4–6 (p < 0.05). After pump removal, the location of the submerged pedestal was shifted to the opposite quadrant for each animal. By day 13 of training, there were no differences in latencies to find the pedestal among the groups; however, those rats that had been treated with 5.0 nmol of Divalinal continued to reveal significantly longer swim distances to find the pedestal than members of the other three groups (p < 0.05). Each group consisted of six rats surgically prepared with 7 d osmotic pumps and were provided 1 d of recovery before initiation of acquisition training.

Representative examples of search patterns in the circular water maze by one member of the group treated with 5.0 (A) and one treated with 0.5 (B) nmol/hr Divalinal during day 6 of acquisition; also, one member from the control group (C) infused with aCSF (0 nmol Divalinal) and a member of the nontreated control group (D). All treatments were intracerebroventricularly delivered via osmotic pump in a volume of 1 μl/hr aCSF. Those animals treated with Divalinal performed poorly compared with members of the groups infused with aCSF or nontreated controls. Specifically, the search pattern strategies of the Divalinal-treated rats were not as sophisticated as the control animals and often included positive thigmotaxis (persistent swimming near the walls of the maze), as evidenced by the animal from the group treated with 5.0 nmol/hr Divalinal (A). Each group consisted of six rats prepared with 7 d osmotic pumps that infused at a rate of 1 μl/hr aCSF. Members of the fourth group served as nontreated controls.
The statistical analyses to support these conclusions indicated a significant difference among the groups (F(3,20)= 3.08; p < 0.05), a days effect over the initial 6 d of acquisition (F(5,100) = 37.16;p < 0.0001), and a significant groups × days interaction (F(15,100) = 4.18; p< 0.0001). Thus, by day 6, members of the two groups treated with Divalinal revealed significantly slower latencies to find the pedestal than those rats infused with aCSF and the noninfused control group (F(3,20) = 3.93; p < 0.05). By day 13, 7 d after pump removal (Fig. 4A), the groups did not differ with respect to latencies to find the pedestal (F(3,20) = 1.66; p > 0.20).
Figure 4B displays the mean ± SEM distance swam to find the submerged pedestal, and there were differences among the groups. On the first day of training, the groups did not differ; however, by the sixth day of acquisition training, those animals that received Divalinal performed poorly compared with those rats that received aCSF or the noninfused controls (F(3,20) = 7.22; p < 0.005). There was improved performance over days (F(5,15) = 31.10; p < 0.0001), and the groups × days interaction was also significant (F(15,100) = 6.29; p < 0.0001) and indicated that the noninfused control group and those animals that received aCSF were superior to both Divalinal groups on days 4–6. Finally, the Divalinal groups did not differ from each other, nor did the aCSF and noninfused control groups differ. By day 13 of training (Fig. 4B), there remained differences among the groups concerning distance swam (F(3,20) = 3.03;p < 0.05). Those rats treated with the 5.0 nmol/hr dose of Divalinal displayed significantly longer swim distances to find the pedestal than members of the other groups.
Results from the probe trials conducted at the conclusion of acquisition training on day 6 are presented in Figure6. There were differences among groups concerning time spent within the target quadrant (F(3,20) = 3.25; p < 0.05) (Figure 6A). Post hoc analyses indicated that those animals treated with the 5.0 and 0.5 nmol doses of Divalinal revealed significantly less time in the target quadrant (37.1 ± 2.6 and 35.2 ± 2.8 sec, respectively) than those infused with aCSF or the control group (45.8 ± 3.7 and 46.8 ± 2.1 sec, respectively). Other comparisons were not different. Figure6B displays the number of entries into the target quadrant by members of each group. There were no differences among the groups on this measure.
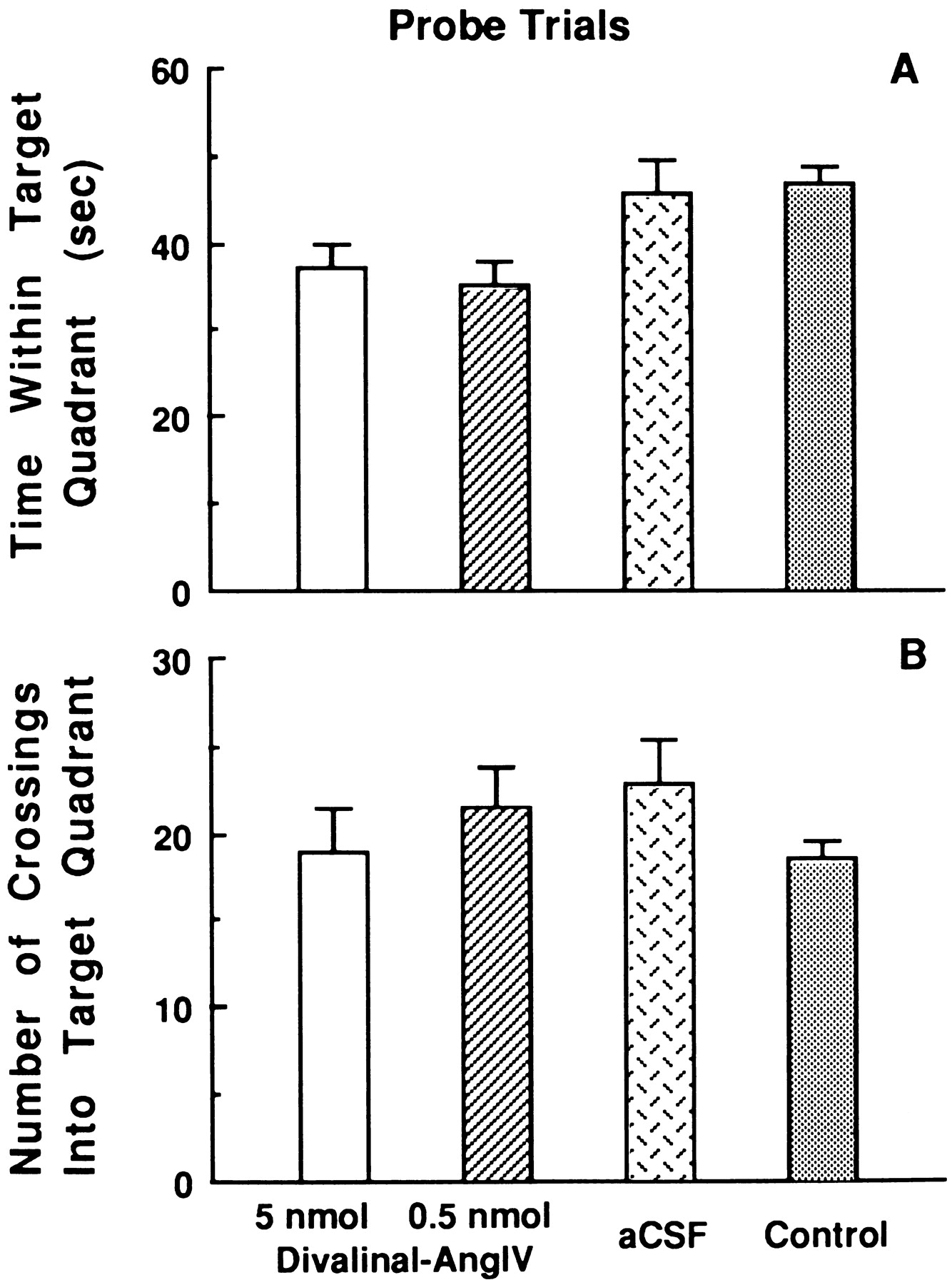
Mean ± SEM group changes and time spent within the correct (target) quadrant (A) and the number of entries into the quadrant (B) during one probe trial conducted at the conclusion of training trials on day 6 of acquisition for animals continuously intracerebroventricularly treated with 5.0, 0.5, or 0 nmol/hr Divalinal via osmotic pump for 6 d and nontreated controls. Those animals treated with 5.0 and 0.5 nmol/hr doses of Divalinal indicated significantly less time spent within the target quadrant compared with those rats that received intracerebroventricular infusion of aCSF or the nontreated control animals (p < 0.05). The groups did not differ with respect to number of entries into the target quadrant.
Perforant path knife cuts
Those rats that received bilateral PP cuts and were subsequently treated with Norleucinal (PP/Norl) revealed significantly shorter latencies (Fig. 7A) and swim path distances (Fig. 7B) than rats prepared with bilateral PP cuts and infused with aCSF (PP/aCSF). In turn, members of the PP/Norl group revealed significantly slower acquisition curves than the group that received bilateral knife cuts to the neocortex, followed by infusion of aCSF (Cor/aCSF), but were equivalent with neocortex knife cut rats that received Norleucinal (Cor/Norl). A 4(groups) × 8(acquisition days) ANOVA of these data revealed a groups effect (F(3,28) = 11.16; p < 0.001), a days effect (F(7,196) = 74.7; p< 0.001), and an interaction effect (F(21,196)= 2.42; p < 0.001). Post hoc analyses indicated that rats in the PP/aCSF group displayed the longest latencies to find the pedestal compared with the other groups on days 2–8. The Cor/aCSF group revealed the shortest latencies on days 2–5 and 7. Members of the PP/Norl and Cor/Norl groups did not differ in their patterns of acquisition. By day 8, the PP/Norl, Cor/aCSF, and Cor/Norl groups did not differ (10.9 ± 1.6, 16.5 ± 3.8, and 18.5 ± 3.4 sec, respectively), whereas the PP/aCSF group indicated significantly longer latencies to find the submerged pedestal (46.8 ± 6.4 sec) (F(3,28) = 14.77;p < 0.001).

Mean ± SEM group changes in latencies (A) and swim distances (B) to find the submerged pedestal in the circular water maze tasks during 8 d of acquisition training by two groups of animals surgically prepared with bilateral PP knife cuts and intracerebroventricular guide cannulas and treated intracerebroventricularly with bolus injections, 2 μl of aCSF or 1.0 nmol of Norleucinal in 2 μl of aCSF, 5 min before the initiation of training trials on each day of acquisition. Two additional groups of animals served as controls and received bilateral knife cuts to the neocortex immediately superior to the PP and were also treated with aCSF or Norleucinal 5 min before training trials each day. Those animals prepared with PP knife cuts and treated with Norleucinal displayed an acquisition curve not different from the control groups prepared with neocortex knife cuts and treated with Norleucinal or aCSF. In contrast, those rats that received PP knife cuts and were infused with aCSF displayed significant impairment in acquisition of the spatial memory task with respect to latencies (p < 0.001) and swim distances (p < 0.001) to find the submerged pedestal. These differences became evident by day 3 of acquisition training and persisted during subsequent days. Each group consisted of eight rats.
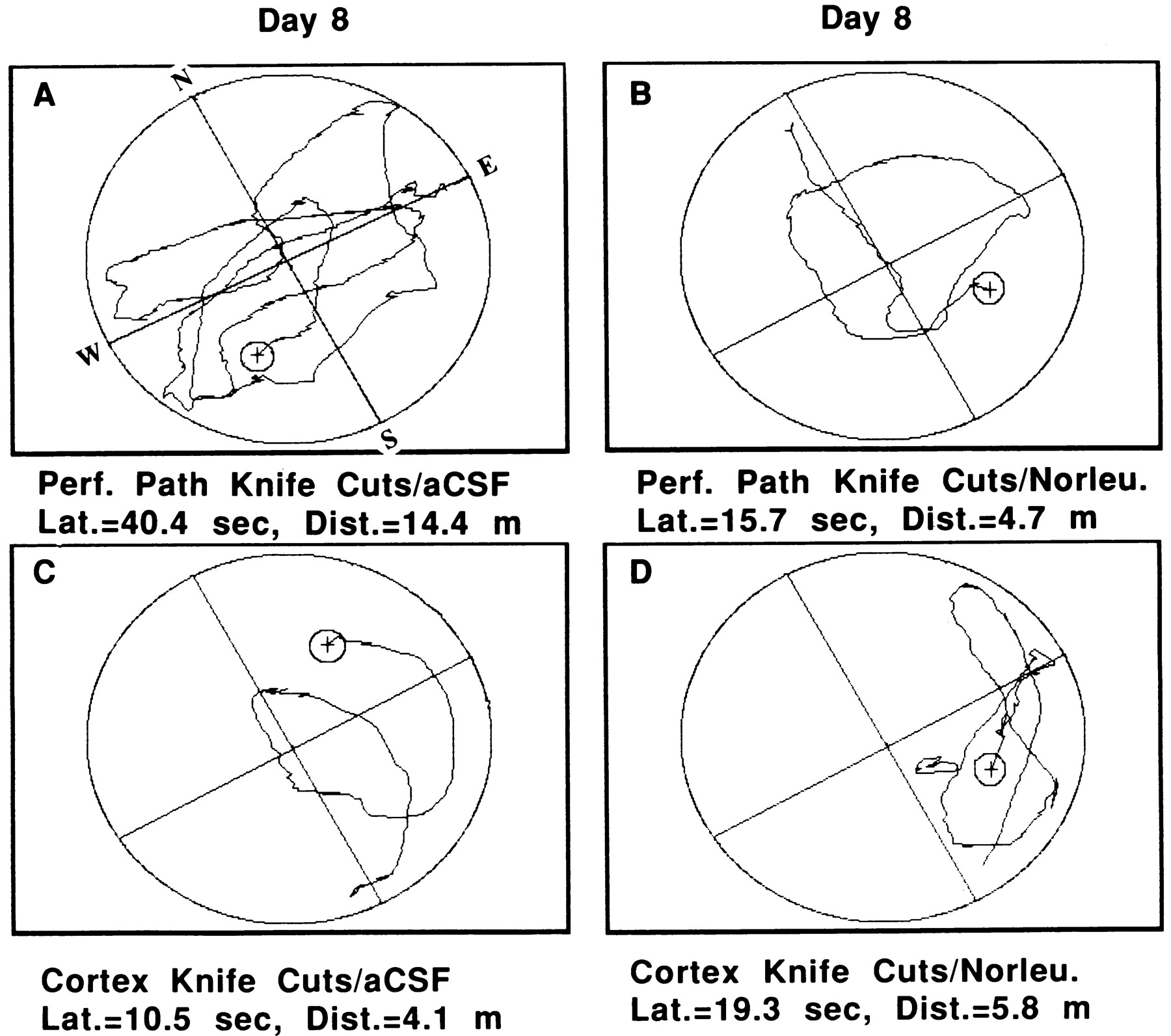
Figure 8 provides representative examples of performance by an animal from each of these four groups on day 8 of acquisition. Those rats prepared with PP knife cuts and treated with aCSF (Fig. 8A) displayed much poorer search strategies than PP knife cut rats treated with Norleucinal (Fig.8B) or neocortex knife cut rats treated with aCSF or Norleucinal (Fig. 8C,D, respectively).
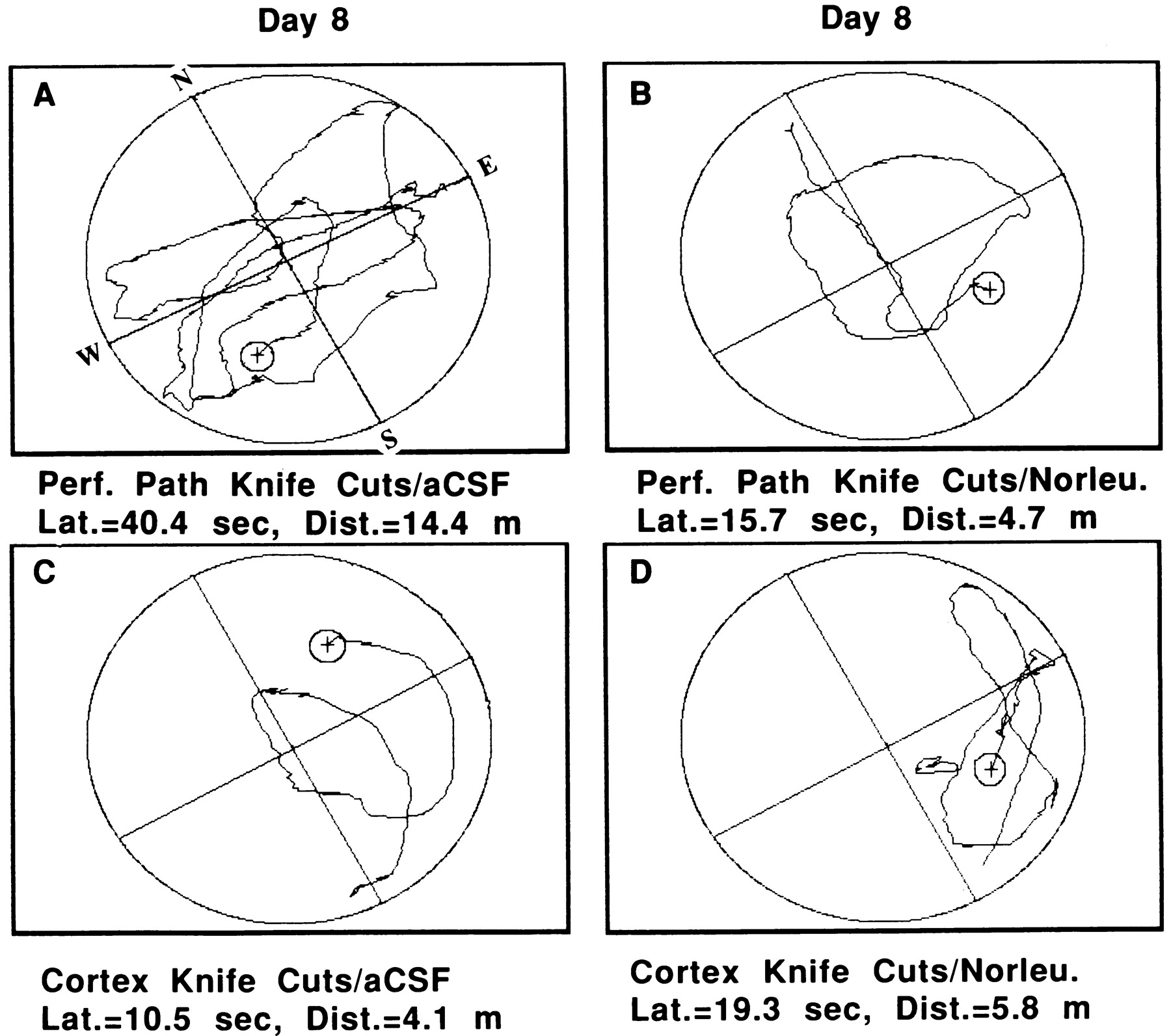
Representative examples of search patterns in the circular water maze by two members of the group prepared with bilateral PP knife cuts. One animal received intracerebroventricular administration of aCSF (A), and one member was treated with Norleucinal (B). For comparison purposes, two rats from the group prepared with bilateral neocortex knife cuts are also presented. One of these animals was from the group that was administered aCSF (C) and one that received Norleucinal (D). The animal that was prepared with PP knife cuts and was subsequently treated with Norleucinal displayed a search pattern that was equivalent with those by animals prepared with neocortex knife cuts and administered aCSF or Norleucinal. In contrast, those animals prepared with PP knife cuts and administered aCSF displayed significantly impaired search strategies (p < 0.001). Each group consisted of eight rats.
Path distance analyses indicated similar group differences as reported above. There was a groups effect (F(3,28) = 8.33; p < 0.001), a days effect (F(7,196) = 63.53; p < 0.001), and an interaction effect (F(21,196) = 64.79;p < 0.001). Post hoc analyses indicated that, by days 5–8 of acquisition, the Cor/aCSF, PP/Norl, and Cor/Norl groups did not differ. Members of all three of these groups displayed significantly shorter swim distances than members of the PP/aCSF group (F(3,28) = 10.08;p < 0.001). By day 8, these mean values were 4.8 ± 0.8, 5.1 ± 1.2, 6.0 ± 0.9, and 14.3 ± 2.3 m for the Cor/aCSF, PP/Norl, Cor/Norl, and PP/aCSF groups, respectively. Probe trials conducted on day 8 indicated no differences among groups concerning time spent in the target quadrant (F(3,28) = 1.36; p > 0.10) or the number of entries into the target quadrant (F(3,28) = 2.36; p > 0.05).
DISCUSSION
The hippocampus has been shown to play an important role in spatial learning and memory (Morris, 1981; McNaughton et al., 1986;Whishaw, 1987; Whishaw and Maaswinkel, 1998; Nadel 1991; Bures et al., 1997). Within the hippocampus, the CA1 field (Sutherland et al., 1983; McNaughton et al., 1989; Morris et al., 1990; Jarrard, 1993) and, to a lesser extent the CA3field (Sutherland et al., 1983; Stubley-Weatherly et al., 1996), are involved in mediating these cortical functions. Related to these observations NMDA receptors are present in high densities on several hippocampal cell types, including CA1–CA3 pyramidal cells. The application of NMDA receptor antagonists has been shown to block the acquisition of spatial learning in rodents (Morris et al., 1986; Davis et al., 1992). The AT4 receptor is heavily distributed within the hippocampus, as well as neocortex, cerebellum, and forebrain cholinergic structures. Of particular relevance to cognitive processing is the possibility that AT4 receptors reside on both pyramidal and granular cells of the hippocampus and medial septal cholinergic neurons that innervate the neocortex and hippocampus. Support for this notion is offered by the observation that AT4 agonists induce fos expression in these cells (Roberts et al., 1995), and iontophoretic application of AT4agonists drives these cells (Albrecht et al., 1997a,b). The linkage to cholinergic neurons is supported by the autoradiographic demonstration that AT4 receptors colocalize with acetylcholinesterase in the spinal cord and the medial forebrain in monkeys (Møeller et al., 1996). Further, recent data from our laboratory indicate that SN56 cells, a cholinergic nucleus basalis–neuroblastoma hybrid (Hammond et al., 1996), possess particularly abundant AT4 receptors (Bmax = 1.74 ± 0.04 pmol/mg protein;Kd = 2.07 ± 0.14 nM; mean ± SEM;n = 3). This localization suggests that AT4receptors may modulate neurotransmission of glutaminergic and cholinergic synapses (glutamate is the neurotransmitter used by both hippocampal pyramidal and granule cells). Thus, activation of AT4 receptors may produce changes in neurotransmitter release, postsynaptic receptors, receptor–intracellular signal coupling, or a combination thereof.
The present investigation initially determined that chronic intracerebroventricular delivery of Nle1-AngIV facilitated acquisition of the circular water maze task of spatial memory during the initial 2 d of training. In contrast, chronic intracerebroventricular delivery of Divalinal significantly interfered with normal acquisition of this task during days 4–6 of training. This Divalinal-induced impairment was not evident during 6 additional days of training after pump removal, suggesting that this deficit in acquisition was reversible. These differences in acquisition could not be attributed to motor and/or sensory impairment as measured by performances on a motor test battery and visible platform protocols. Further, acute intracerebroventricular treatment with Norleucinal restored normal acquisition of a spatial memory task in animals prepared with bilateral perforant path knife cuts.
One potential mechanism underlying these AT4 agonist effects relates to their ability to enhance cerebral blood flow. Increases in cerebral blood flow have been positively correlated with cognition. Angiotensin IV (Haberl et al., 1991; Kramár et al., 1997) and Norleucinal (Kramár et al., 1998) have been shown to increase cerebral blood flow by vasodilation of arterioles. These increments in blood flow appear to be nitric oxide-dependent. Thus, the presently noted improvement in memory could be mediated via elevations in cerebral blood flow. Along these lines, de la Torre (1994) has suggested that Alzheimer’s disease may, in part, be caused by distortion of brain capillaries that prevent normal blood flow, thus producing ischemia with consequential damage to CA1 field cells and other ischemic sensitive brain structures. Thus, increased blood flow to intact hippocampal neurons, as well as other neurons, may facilitate performance. A complimentary hypothesis by Sato and Sato (1995) proposes that cholinergic fibers possessing cell bodies within the NBM and the medial septum normally release acetylcholine within the hippocampus and neocortex, which in turn produces vasodilation and accompanying elevations in cerebral blood flow. Application of an AT4 agonist could potentially stimulate cholinergic neurons in the NBM and the medial septum, where there are high densities of AT4 receptors, reinstating normal acetylcholine release within the hippocampus and neocortex and, in turn, incrementing blood flow.
A second possible explanation relates to numerous studies that have demonstrated structural changes in the architecture of synaptic connections concomitant with the development of learning and memory (for review, see Agnihotri et al., 1998). These changes may be mediated by adhesive molecules that determine cell-to-cell and cell-to-extracellular matrix interactions in the brain. Recent data from our laboratory indicate that the AT4 receptor plays a potentially pivotal role in the restructuring of the extracellular matrix in numerous tissues, including the brain (M. S. Cummings, J. M. Hanesworth, S. E. Hunter, and J. W. Harding, unpublished observations). Specifically, AT4receptors mediate the expression of matrix metalloproteinases (MMPs), their inhibitors, and members of the plasminogen-plasmin cascade, which are responsible for MMP activation. These observations raise the possibility that the effect of AT4 activation may, in part, result from synaptic remodeling.
The majority of strategies concerned with the development of a pharmacological treatment for cognitive dysfunction have been stimulated by our current knowledge of long-term potentiation (LTP) processes. This LTP process by which synaptic strength is augmented in an activity-dependent manner has been correlated to cognitive processes (Lynch et al., 1983; Malenka et al., 1988; Izquierdo, 1993; Rison and Stanton, 1995; Wayner et al., 1995). The interplay of acetylcholine with LTP appears to be at the level of intracellular calcium and/or pyramidal cell depolarization. Working via muscarinic receptors, acetylcholine both increases intracellular calcium via IP3-dependent mechanisms and depolarizes by attenuating potassium efflux, which acts to prime the cells for LTP. The importance of this process is illustrated by the effectiveness of muscarinic blockers as amnesiacs. Interestingly, AT4 agonists can compensate for these muscarinic receptor antagonist-induced deficits (Pederson et al., 1998), suggesting that either acetylcholine release is augmented so that the effect of competitive muscarinic antagonists are overcome or, more likely, these agonists act independently to elevate intracellular calcium and “prime” the system to respond more effectively to glutamate. This notion is supported by the observation that AT4 agonists increase intracellular calcium in cardiac myocytes, another excitable cell (B. K. Slinker, J. W. Harding, and S. Simasko, unpublished observations). Consistent with this idea are recent results indicating that AT4 agonists facilitate LTP both in vitro (E. A. Kramár, unpublished observations) andin vivo (Ikeda et al., 1998). Thus, one pharmacological approach concerns enhancing cholinergic neurotransmission. Prototype drugs include acetylcholinesterase inhibitors, such as Cognex (Parke-Davis, Morris Plains, NJ; Warner-Lambert, Morris Plains, NJ), Aricept (Eisai, Tokyo, Japan; Pfizer, Groton, CT), Excelon (Novartis, Summit, NJ), Mentane (Hoechst Marion Roussel, Frankfurt am Main, Germany), and Metrifonate (Bayer, Wuppertal, Germany), which block acetylcholine removal from the synaptic cleft. A related strategy utilizes acetylcholine analogs possessing increased metabolic stability such as Memric (SmithKline Beecham, Upper Merion, PA), Xanomeline (Eli Lilly, Indianapolis, IN), and CI979 (Warner-Lambert) (for review, seeMarx, 1996).
Our laboratory is presently developing nonpeptidic agonists that bind with high affinity to brain-specific AT4 receptor subtypes. Results from the present investigation provide an initial demonstration of the potentially important role of the brain AT4 receptor system in normal cognitive function and encourage the possible use of AT4 receptor agonists to facilitate cognitive performance in compromised individuals.
Footnotes
This research was supported by the Edward E. and Lucille I. Laing Endowment for Alzheimer’s Research and Washington State University. We thank Dr. Ronald Skelton for his expert advice concerning the perforant pathway knife cut procedure. We also thank Ruth Day for her excellent secretarial help in preparing this manuscript.
Correspondence should be addressed to John W. Wright, Department of Psychology, Washington State University, Pullman, WA 99164-4820.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}