Article Text

Abstract
Background and aims Inflammatory bowel disease (IBD), comprising Crohn´s disease and ulcerative colitis, is characterised by chronic relapsing inflammation of the gut. Increased proteasome activity, associated with the expression of immunoproteasomes, was found to enhance proinflammatory signalling and thus promotes inflammation in patients with IBD. The aim of this study was to explore whether modulation of the proteasomal activity is a suitable therapeutic approach to limit inflammation in colitis.
Methods This concept was assessed in two different experimental set-ups. Development of dextran sodium sulfate (DSS)-induced colitis was tested (1) in lmp7−/− mice lacking the immunoproteasome subunit LMP7 and (2) in wild-type (WT) mice treated with the proteasome inhibitor bortezomib.
Results Compared with WT mice, lmp7−/− mice develop significantly attenuated colitis due to reduced nuclear factor-κB (NF-κB) signalling in the absence of LMP7. Further, treatment with bortezomib revealed dose-dependent amelioration of DSS-induced inflammation. In both approaches modulation of the proteasome activity limited the secretion of proinflammatory cytokines and chemokines. Consequently, infiltration of the colon by neutrophils and expansion of inflammatory T helper 1 (Th1) and Th17 T cells was diminished and thus prevented excessive tissue damage.
Conclusions It was demonstrated that modulation of the proteasome activity is effective in attenuating experimental colitis. The results reveal that reduction of the proteasome activity either by partial inhibition with bortezomib or by specifically targeting the immunoproteasome subunit LMP7 is a suitable treatment of intestinal inflammation.
- Basic sciences
- bortezomib
- experimental colitis
- IBD basic research
- immunology
- inflammation
- inflammatory bowel disease
- LMP7
- proteasome
Statistics from Altmetric.com
- Basic sciences
- bortezomib
- experimental colitis
- IBD basic research
- immunology
- inflammation
- inflammatory bowel disease
- LMP7
- proteasome
Significance of this study
What is already known about this subject?
A dysregulated activation of NF-κB, which drives the expression of many proinflammatory mediators, commonly occurs in IBD.
Proteasomes isolated from the inflamed mucosa of patients with IBD have a high content of immunosubunits, especially LMP7.
Enhanced proteasome activity, associated with the expression of immunosubunits, is involved in the sustained NF-κB-driven inflammation in IBD.
The anti-inflammatory capacity of proteasome inhibitors such as bortezomib and MG132 has been tested previously. The outcome of these studies has been controversial depending on the model that was used.
What are the new findings?
Mice deficient in the immunosubunit LMP7 reveal attenuated DSS-induced colitis due to reduced expression of many proinflammatory mediators regulated by NF-κB.
Partial inhibition of the proteasome activity by treatment with bortezomib attenuates DSS-induced colitis.
Treatment of DSS-induced colitis with non-selective proteasome inhibitors such as bortezomib, which equally target constitutive subunits and immunosubunits, causes dose-dependent adverse effects including increased mortality.
Adverse effects were not observed in LMP7-deficient mice. Thus, selective inhibition of LMP7 is a promising strategy to limit inflammation in colitis and should be preferred over non-selective inhibition of proteasomes.
How might it impact on clinical practice in the foreseeable future?
The results of this study suggest that selective inhibition of proteasomes by specifically targeting LMP7 is a promising approach for the treatment of IBD.
Introduction
Inflammatory bowel disease (IBD) comprises inflammatory disorders of the gut with largely unknown aetiology. IBD can be classified into Crohn´s disease (CD) or ulcerative colitis (UC). Both subtypes are characterised by chronic relapsing inflammation with various intertwined effector mechanisms, of which the initial trigger remains unclear.1 2 The expression of many proinflammatory mediators in IBD is controlled by nuclear factor-κB (NF-κB).3 4 The proteasome, a multimeric protease complex, mediates NF-κB activation by degradation of the inhibitors of κB (IκBs) and processing of the NF-κB subunit p105 to p50 which forms a heterodimer with p65 (RelA).5–9 Two subsets of catalytic subunits can be integrated into 20S proteasomes: either the constitutive subunits β1, β2 and β5 or their interferon γ (IFNγ)-inducible counterparts, the immunosubunits LMP2 (β1i), MECL-1 (β2i) and LMP7 (β5i).10 11 We recently identified the proteasome as a key molecule allowing the distinction between UC and CD according to its subunit composition. While patients with CD exhibit high amounts of immunosubunits in the inflamed gut, patients with UC mostly express constitutive subunits, with the exception of LMP7 which is highly abundant in both disease types.12 The proteasomes isolated from both patients with CD and those with UC, revealed accelerated cleavage of IκBα and p105 as compared with non-inflamed mucosal tissue. However, the proteasomes of patients with CD, containing higher amounts of immunosubunits compared with those of patients with UC, were superior in promoting NF-κB activation. This demonstrates that increased proteasomal activity, associated with the expression of immunosubunits, is involved in sustained progressive inflammation in IBD.13
Thus, we tested whether modulation of the proteasome activity is suitable to reduce intestinal inflammation. First, the efficacy of targeting the proteasome was evaluated following induction of colitis in LMP7-deficient mice. We chose LMP7 as target because it is highly abundant in both patients with CD and those with UC.12 13 In addition, LMP7 is known to promote the integration of the other immunosubunits, resulting in an overall reduced abundance of these subunits in lmp7−/− mice.14 15 Further, we evaluated whether bortezomib, a non-selective proteasome inhibitor, which equally targets constitutive subunits and immunosubunits, is able to limit inflammation. The anti-inflammatory capacity of non-selective proteasome inhibitors such as bortezomib and MG132 has been previously assessed in various models of inflammation.16–19 However, the results have been controversial depending on the model that was used.
Here, we analysed the dose-dependent effects of bortezomib in the treatment of DSS-induced colitis. We observed a wide range of effects ranging from no amelioration at low doses, to attenuation of inflammation at intermediate doses, to increased mortality at high doses. In contrast, DSS-induced colitis was strongly attenuated in lmp7−/− compared with wild-type (WT) mice, but adverse effects were not observed. Taken together, these results suggest that modulation of the proteasome activity is a suitable treatment to dampen inflammation in colitis. Moreover, our findings suggest that targeting of LMP7 in particular is a promising approach for the treatment of IBD.
Materials and methods
Experimental animals
Mice were kept under specific pathogen-free (SPF) conditions. C57Bl/6 mice (WT) were purchased from Charles River Laboratories (Berlin, Germany). Breeding pairs of lmp7−/− mice on the C57Bl/6 background, originally described by Fehling et al,20 were kindly provided by Professor H J Schild (Johannes Guthenberg University, Mainz, Germany). All experiments were performed according to the German animal protection law.
Induction of experimental colitis by dextran sodium sulfate (DSS)
Age- (9–11 weeks) and sex-matched (female) WT and lmp7−/− mice were used. DSS 3% (w/v) (35 000–50 000 kDa, MP Biomedicals, Heidelberg, Germany) was administered via the drinking water for 5 days.
Inflammation scoring
Formalin-fixed colon sections were stained with H&E. The inflammation scoring was performed blinded by a pathologist and is a combination of inflammatory cell infiltration and tissue damage. Points for infiltration were given as follows: 0, no infiltration; 1, increased number of inflammatory cells in the lamina propia; 2, inflammatory cells extending into the submucosa; 3, transmural inflammatory infiltrates; and for tissue damage: 0, no mucosal damage; 1, discrete epithelial lesions; 2, erosions or focal ulcerations; 3, severe mucosal damage with extensive ulceration extending into the bowel wall.
Cell cultures
Murine embryonic fibroblasts (MEFs) were derived from 13- to 14-day-old embryos of WT or lmp7−/− mice. Embryos were decapitated, inner organs removed and the residual, minced tissue was digested in 1× trypsin-EDTA solution (Gibco, Darmstadt, Germany) at 37°C. The isolated cells were seeded in Dulbecco's modified Eagle's medium (DMEM) containing 10% fetal calf serum (FCS), 1 mM l-glutamine (PAA, Cölbe, Germany), penicillin/streptomycin (PAA) and 1 mM sodium pyruvate (Biochrom AG, Berlin, Germany) (D10 medium) at 37°C and 5% CO2.
Nuclear translocation of p65 in MEFs by automated microscopy
WT and lmp7−/− MEFs were seeded in 96-well plates in D10 medium complemented with 50 U/ml IFNγ (Active Bioscience, Hamburg, Germany) and cultured for 48 h at 37°C, 5% CO2. Afterwards cells were stimulated with 40 ng/ml tumour necrosis factor α (TNFα; Active Bioscience) and fixed with 4% paraformaldehyde. Cells were permeabilised with 0.2% Triton X-100 in phosphate-buffered saline (PBS) and blocked with 3% bovine serum albumin (BSA) in PBS. Staining was performed with anti-p65 polyclonal antibodies (Abs) (sc-109, Santa Cruz), followed by Cy3-conjugated secondary Abs (Dianova, Hamburg, Germany) and Hoechst 33342. Four pictures per well were acquired using the automated microscopy system Scan^R (Olympus, Hamburg, Germany). The percentage of cells with significant co-localisation of the p65-Cy3 and the Hoechst staining was determined by using the Scan^R picture analysis software (Olympus).
Gene array analysis
Microarray experiments were performed as dual-colour hybridisations. Colon samples were briefly incubated in 4 M guanidinium isothiocyanate and total RNA was extracted with TRIzol (Invitrogen, Carlsbad, CA, USA). Quality control and quantification of total RNA was assessed using an Agilent 2100 bioanalyser (Agilent Technologies, Santa Clara, CA, USA). RNA labelling was performed with the Low RNA Input Fluorescent Linear Amplification Kit (Agilent Technologies). A 1.25 μg aliquot of each labelled cRNA was fragmented and hybridised to whole mouse genome 44k microarrays (Agilent Technologies). After scanning of microarrays with a DNA microarray laser scanner (Agilent Technologies), raw microarray image data were analysed with the Image Analysis software G2567AA (Version A.9.5.1, Agilent). The extracted files were further analysed with the Rosetta Resolver Biosoftware, Build 6.1 (Rosetta Biosoftware, Cambridge, MA, USA). Ratio profiles comprising single hybridisations were combined in an error-weighted fashion to create ratio experiments. A twofold change in expression cut-off for ratio experiments was applied together with anticorrelation of ratio profiles with a p value cut-off of 0.01.
Measurement of cytokine and chemokine secretion by ex vivo colon culture
Colon explants (0.05 g) were cultured in RPMI 1640 supplemented with gentamycin (50 μg/ml), penicillin/streptomycin (PAA) and 10% FCS. The supernatants were harvested after 24 h at 37°C, 5% CO2. Cytokine and chemokine concentrations were measured using the Bio-Plex Bead array technology according to the manufacturer´s instructions (Bio-Rad Laboratories, Munich, Germany).
Generation of bone marrow chimeras
Recipient mice were irradiated twice with 5 Gy. One day after irradiation recipient mice were reconstituted with 5×106 bone marrow cells isolated from the respective donor mice and given intravenously. Mice received antibiotics via the drinking water (0.1 mg/ml ciprofloxacin, 0.2 mg/ml neomycin) for 4 weeks. Colitis was induced 10 weeks after reconstitution.
Immunofluorescence microscopy
Formalin-fixed colon sections were incubated in LAB solution (Polysciences Inc, Eppelheim, Germany) for 15 min at room temperature. The tissue sections were blocked with 5% horse serum, 0.1% Tween-20 in PBS, followed by staining with antimyeloperoxidase-specific Abs (A0398, Dako, Glostrup, Denmark) and Cy3-conjugated secondary Abs (Dianova). The nuclear staining was performed with Hoechst 33342. Images were acquired using a Leica DMRB microscope and ProgRes CapturePro 2.7 software (Jenoptik, Jena, Germany).
Bortezomib treatment
Bortezomib (LC Laboratories, Woburn, MA, USA) (0.5, 0.35 0.2 or 0.1 mg/kg, or vehicle only (PBS)) was administered intraperitoneally starting on the first day of DSS exposure for a period of 10 days.
Isolation and flow cytometric analysis (FACS) of lamina propria mononuclear cells (LPMCs)
Colon samples were shaken in RPMI 1640 medium supplemented with 10% FCS, 1 mM l-glutamine, penicillin/streptomycin (colon medium) for 30 min at 37°C. Intraepithelial mononuclear cells were removed by rigorous shaking in PBS containing 2% FCS. LPMCs were isolated by digestion in RPMI 1640 supplemented with 0.4 mg/ml collagenase D and collagenase VIII (Sigma Aldrich, Munich, Germany) under constant stirring (120 rpm) for 1 h at 37°C. The cell suspension was filtered through a 120 μm iron mesh, washed, resuspended in 40% Percoll, which was then layered on 70% Percoll, both in RPMI 1640, and centrifuged for 20 min at 2000 rpm. The LPMCs were recovered from the interphase and resuspended in colon medium
LPMCs were stained with anti-MHC-II-PE (clone TIB120), anti-CD11c-FITC (clone N418), anti-CD11b-PE-Cy7 (eBioscience, San Diego, CA, USA) and anti-Ly6c/Ly6G-Pacific Blue (Gr-1) (eBioscience) for 30 min on ice.
For intracellular cytokine staining, the LPMCs were either left untreated or were stimulated with 4 μg/ml anti-CD3 and 8 μg/ml anti-CD28 for 6 h at 37°C, 5% CO2 supplemented with brefeldin A (10 μg/ml) (Sigma Aldrich) after 1 h. The cells were stained with anti-CD4-Pacific Blue (eBioscience) and anti-CD8-PerCP (clone H35-17.2) for 30 min on ice. After fixation with 4% paraformaldehyde in PBS and permeabilisation with 0.5% (w/v) saponin buffer, intracellular staining was performed with anti-IL-17-PE (eBioscience) and anti-IFNγ-PE-Cy7 (eBioscience) on ice for 30 min. The cells were analysed on an LSRII flow cytometer (BD Biosciences, Heidelberg, Germany) using the FACS Diva software (BD Biosciences).
Statistical analysis
Statistical analysis was performed using Student t test. Differences of p<0.05 were considered to be significant. Differences in survival rates were evaluated by the log rank test (Mantel Cox). All calculations were performed using GraphPad Prism 5.0.
Results
Attenuated DSS-induced colitis in lmp7−/− mice
To assess the impact of the immunosubunit LMP7 on the development of colitis, we exposed WT and lmp7−/− mice to DSS. Compared with DSS-treated WT mice, we observed significantly less weight loss in lmp7−/− mice (figure 1A). While WT mice slowly recovered from the weight loss, lmp7−/− mice regained normal weight already 12 days after DSS exposure (figure 1A). The reduction in colon length, a marker of intestinal inflammation, was also less pronounced in lmp7−/− mice (figure 1B). Histologically, WT mice showed strong infiltration of the lamina propria and submucosa following DSS exposure, associated with severe crypt damage. In contrast, the infiltrates in lmp7−/− mice were much smaller and less tissue damage was observed (figure 1D). Accordingly, the inflammation scores in lmp7−/− mice were lower throughout the course of colitis (figure 1C). Thus, the absence of LMP7 strongly attenuates DSS-induced colitis and allows rapid healing.

Dextran sodium sulfate (DSS)-induced colitis is attenuated in lmp7−/− mice. Change of weight was monitored either in untreated controls or in DSS-treated mice over a period of 18 days (n=12). One representative of three independent experiments is shown (A). The colon length in wild-type (WT) and lmp7−/− mice was measured at the indicated time points. Shown are mean values ± SEM of four mice per group, and one of three representative experiments (B). The inflammation scoring according to histological analysis of H&E-stained, paraffin-embedded colon sections was performed blinded in four mice per group and time point. Shown are mean values ± SEM, and one of two representative experiments is shown (C). Representative images of H&E-stained colon sections of control or DSS-treated mice (day 8), which were used for the inflammation scoring, are shown under ×10 and ×40 magnification, respectively (D). *p<0.05; **p<0.01; ***p<0.001.
Expression of proinflammatory mediators is strongly reduced in lmp7−/− mice
A microarray analysis was performed to determine which inflammatory mediators were affected by the lack of LMP7. We observed reduced expression of various proinflammatory mediators, cytokines and chemokines in DSS-treated lmp7−/− mice (figure 2A; data set available at http://www.ncbi.nlm.nih.gov/geo;accession-number=GSE18163). In particular the classical proinflammatory cytokines interleukin 1 α/β (IL-1α/β), IL-6 and TNFα were highly expressed in DSS-treated WT mice, but only a slight or even no induction was found in lmp7−/− mice. Further, the T helper 1 (Th1)/Th17 cytokines IFNγ and IL-17 were induced in WT but not lmp7−/− mice (figure 2A). In contrast, the Th2 cytokines IL-4, IL-5 and IL-13 were not differentially regulated in either group of mice (figure 2A).

Reduced expression of proinflammatory cytokines and chemokines in dextran sodium sulfate (DSS)-treated lmp7−/− mice. Microarray analysis was performed by comparison of naïve control mice with DSS-treated mice (4 and 8 days after treatment). RNA samples of three mice per group and time point were pooled for the analysis. Genes upregulated in DSS-treated mice versus the naïve controls are shown in red, downregulated genes in green and genes not differentially regulated in black (A). The cytokine (B) and chemokine secretion (C) by colon explants of wild-type (WT) and lmp7−/− mice were analysed using the Bioplex bead array technology. Shown are mean values ± SEM of four mice per group and time point, and one of two representative experiments is shown. *p<0.05; **p<0.01; ***p<0.001. IFNγ, interferon γ; IL, interleukin; MCP, monocyte chemotactic protein; MIP, macrophage inflammatory protein; nd, not detectable; RANTES, regulated upon activation, normal T cell expressed and secreted; TNFα, tumour necrosis factor α.
With regard to chemokine expression, profound differences were observed for the neutrophil-attracting chemokines KC, macrophage inflammatory protein-2 (MIP-2) and RANTES (regulated upon activation, normal T cell expressed and secreted), which were highly upregulated in WT but not lmp7−/− mice (figure 2A). On the protein level, we found reduced secretion of selected cytokines and chemokines by colon explants of DSS-treated lmp7−/− mice, confirming the mRNA expression data (figure 2B,C). The diminished secretion of IL-6 by lmp7−/− colon explants was also observed when tissues were stimulated with the Toll-like receptor (TLR) ligands lipopolysaccharide (LPS), lipoteichoic acid (LTA) or flagellin (Supplementary figure 1A). However, the uptake of fluorescein isothiocyanate (FITC)–dextran was similar between WT and lmp7−/− mice, suggesting that the lack of LMP7 has no direct effect on the epithelial barrier function itself (Supplementary Figure 1B). This finding indicates that NF-κB signalling in response to bacterial components sensed through TLRs is affected in lmp7−/− mice.
Lack of LMP7 affects inflammatory signalling in mucosal immune cells
Following leakage of bacterial components through the epithelial barrier, inflammation can be induced through TLR on intestinal epithelial cells (IECs) or by mucosal immune cells. To assess, in which of these cell populations the lack of LMP7 affects the proinflammatory signalling, we generated bone marrow chimeras of WT and lmp7−/− mice. lmp7−/− recipient mice reconstituted with WT bone marrow developed exacerbated colitis compared with lmp7−/− mice complemented with lmp7−/− bone marrow (figure 3A), which was accompanied by reduced colon length and increased levels of proinflammatory cytokines (figure 3B,C). Reciprocally, reconstitution of WT mice with lmp7−/− bone marrow attenuated DSS-induced weight loss, shortening of the colon and cytokine secretion compared with WT recipients receiving WT bone marrow (figure 3D–F). Thus, the lack of LMP7 predominantly affects the induction of the proinflammatory programme within mucosal immune cells, but plays a minor role in IECs.

Lack of LMP7 affects the induction of inflammation by haematopoietic cells The bone marrow chimeras were treated with 3% dextran sodium sulfate (DSS) for 5 days 10 weeks after their reconstitution. The change of weight was monitored in controls (n=4) and DSS-treated mice (n=11) over a period of 18 days (A, D). The colon length of the respective bone marrow chimeras was assessed in four mice per group and time point, and mean values ± SEM are shown (B, E). The cytokine concentrations secreted by colon explants of the diverse chimeric mice were analysed using the Bioplex bead array technology. Shown are mean values ± SEM of four mice per group and time point (C, F). *p<0.05; **p<0.01; ***p<0.001. BM, bone marrow; WT, wild type.
Reduced activation of NF-κB in the absence of LMP7
Many of the proinflammatory mediators, cytokines and chemokines strongly induced in WT but not lmp7−/− mice are regulated by NF-κB (figure 2). To verify that LMP7 promotes NF-κB activation, lmp7−/− and WT MEFs were stimulated with TNFα, and the nuclear translocation of p65, a hallmark of NF-κB activation, was followed by immunofluorescence microscopy. Upon TNFα stimulation, WT MEFs reacted by nuclear translocation of p65 for a period of 60 min. Thereafter the number of cells with translocated p65 declined, resembling a normal circuit of NF-κB activation.21 In contrast, only a few lmp7−/− MEFs reacted by detectable nuclear translocation of p65 in response to TNFα, demonstrating that the NF-κB activation is impaired in the absence of LMP7 (figure 4A,B). This is in line with our previous findings that proteasomes containing LMP7 drive NF-κB activation in inflamed tissues of patients with IBD.13

Reduced nuclear factor-κB (NF-κB) activation in LMP7-deficient cells. Wild-type (WT) and lmp7−/− murine embryonic fibroblasts (MEFs) pretreated with interferon γ (IFNγ; 50 U/ml) for 48 h were stimulated with tumour necrosis factor α (TNFα; 40 ng/ml). Intracellular staining for nuclear DNA (blue) and for the NF-κB subunit p65 (red) was performed at the indicated time points, and fluorescence images were acquired with an automated microscope. Representative fluorescence images of selected time points and three independent experiments are shown (A). The percentage of cells with significant nuclear translocation of p65 was determined in three replicate wells and four images per well in each experiment. Shown are mean values ± SEM of three independent experiments (B). *p<0.05; **p<0.01; ***p<0.001.
Reduced neutrophil influx in the colon of DSS-treated lmp7−/− mice
In addition to the differential expression of neutrophil-attracting chemokines, a group of genes which are predominantly expressed by neutrophils were highly upregulated in DSS-treated WT but not lmp7−/− mice, indicating a reduced neutrophil influx in the absence of LMP7. These genes include S100A8/S100A9 (calgranulin A/B), which are commonly used as markers for disease activity in patients with IBD22 (figure 5A). Histologically, reduced numbers of neutrophils were observed within the infiltrates in the submucosa and lamina propria oflmp7−/− mice (figure 5B). The flow cytometric analysis revealed that the frequency of neutrophils (Gr-1high MHC IIneg cells) within the LPMC was decreased by ∼50% in lmp7−/− compared with WT mice (figure 5C). Due to the overall reduced infiltration of the colon by immune cells in lmp7−/− mice, this difference is even more pronounced when the total numbers of neutrophils in the LPMCs were calculated (figure 5D). Hence, the influx of neutrophils into the colon was significantly reduced in lmp7−/− mice. As some of the differentially regulated chemokines act not only on neutrophils (figure 2C), but also on other immune cells, the number of infiltrating macrophages (CD11bhigh MHC IIhigh cells) was also reduced in DSS-treated lmp7−/− mice. Interestingly, no effect on dendritic cells (CD11chigh MHC IIhigh cells) was observed (Supplementary figure 2A).

Lack of LMP7 limits neutrophil influx in dextran sodium sulfate (DSS)-induced colitis. Microarray analysis was performed by comparison of naïve control mice with DSS-treated mice (4 and 8 days after treatment). RNA samples of three mice per group and time point were pooled for the analysis. Genes upregulated in DSS-treated mice versus the naïve controls are shown in red, downregulated genes in green and genes not differentially regulated in black (A). Paraffin-embedded colon sections of wild-type (WT) and lmp7−/− mice, either naïve or 8 days after DSS treatment, were stained for DNA (blue) and myeloperoxidase (MPO; red) (×40 magnification). Representative fluorescence images of four mice per group and time point are shown (B). Lamina propria mononuclear cells (LPMCs) were isolated from naïve and DSS-treated mice 8 days after exposure, stained with antibodies specific for Gr-1 and major histocompatibility (MHC) class II, and analysed by flow cytometry (C). The total numbers of neutrophils were calculated from the total number of LMPCs that were isolated from each mouse at the indicated time points. Shown are mean values ± SEM of six mice per group, and one of two representative experiments is shown (D). *p<0.05; **p<0.01; ***p<0.001.
Reduced expansion of Th1 and Th17 cells in the colon of DSS-treated lmp7−/− mice
We observed increased mRNA expression and secretion of Th1 and Th17 cytokines in DSS-treated WT but not lmp7−/− mice (figure 2A,B). Accordingly, we found a lower frequency of Th1 and Th17 T cells within the LPMCs of DSS-treated lmp7−/− compared with WT mice (figure 6A, C). The difference in the expansion of these CD4+ T cell subsets was even more pronounced when the total cell numbers were calculated. The total numbers of Th1 and Th17 cells within the LPMCs of DSS-treated lmp7−/− mice did not rise above background levels, whereas a significant expansion of these populations was found in WT mice (figure 6B,D). It was shown that DSS-induced colitis in C57Bl/6 mice progresses to chronic inflammation already after a single DSS treatment and that Th1 and Th17 cells are involved in this disease progression.23 Thus, the lack of proinflammatory T cells could explain why lmp7−/− mice rapidly recovered from DSS-induced colitis, while WT mice exhibit signs of prolonged inflammation. Interestingly, although major histocompatibility complex (MHC) class I antigen presentation is reduced in lmp7−/− mice,20 the expansion of CD8+ T cells was not affected in these mice (Supplementary figure 2B).

Markedly reduced numbers of T heper 17 (Th17) and Th1 T cells in dextran sodium sulfate (DSS)-treated lmp7−/− mice. For intracellular cytokine staining of Th1 and Th17 T cells, lamina propria mononuclear cells (LPMCs) were isolated 12 days after DSS treatment and restimulated with αCD3/CD28 monoclonal antibody. The frequency of Th1 T cells (CD4+ IFNγ+) (A) or Th17 cells (CD4+ IL-17+) (C) within the LPMCs was determined by flow cytometric analysis. Representative dot plots are shown and the given frequencies represent mean values ± SEM of six mice per group (A, C). The total numbers of Th1 (B) and Th17 T cells (D) were calculated from the total number of LMPCs that were isolated from each mouse at the indicated time points. Shown are mean values ±SEM of six mice per group (B, D). *p<0.05; **p<0.01; ***p<0.001. IFNγ, interferon γ; IL-17, interleukin 17.
Dose-dependent effects of bortezomib in DSS-induced colitis
The beneficial effects of LMP7 deficiency on inflammation are caused by reduced proteasome activity. Although bortezomib is not selective for constitutive subunits or immunosubunits, we wondered whether a general reduction of the proteasome activity will be effective in attenuating colitis.
When mice were treated with bortezomib, a clear dose-dependent modulation of DSS-induced colitis was observed. The lowest dose (0.1 mg/kg) showed beneficial effects in reducing inflammation as measured by change of weight and colon length (figure 7A,C), while cytokine and chemokine secretion were only marginally affected (figure 7B, figure 8B). In contrast, intermediate doses (0.2 and 0.35 mg/kg) were effective in attenuating colitis in all parameters tested. In particular, treatment with 0.35 mg/kg bortezomib prevented the weight loss and shortening of the colon usually observed between days 6 and 12 following DSS exposure (figure 7A,B). In addition, the secretion of proinflammatory cytokines and chemokines was markedly diminished (figure 7C, figure 8B). Consequently, the neutrophil influx into the colon and the associated tissue damage was almost completely abrogated in mice treated with 0.35 mg/kg bortezomib (figure 8A). However, treatment of mice with intermediate doses induced an early weight loss on days 2–4 after DSS exposure, which was not observed in vehicle controls (figure 7A). This early weight loss was not accompanied by secretion of proinflammatory cytokines and chemokines or increased shortening of the colon in the inhibitor-treated groups and thus cannot be attributed to an earlier onset of DSS-induced colitis (figures 7B,C, 8B). Moreover, treatment with a high dose of bortezomib (0.5 mg/kg) resulted in 50% mortality in the DSS-treated mice (figure 7D). Similar results were obtained when mice were treated with the proteasome inhibitor MG132 (Supplementary figure 3). While a low dose of MG132 attenuated colitis and limited cytokine and chemokine secretion by colon explants (Supplementary figure 3A–B, D–E), high doses led to increased mortality under DSS treatment (Supplementary figure 3C). It is noteworthy that the treatment controls, which received bortezomib or MG132 but no DSS, showed neither early weight loss nor increased mortality (data not shown, figure 7D, Supplementary figure 3C). In summary, these results show that gradual proteasome inhibition is effective in attenuating inflammation. However, the effective dose of non-selective proteasome inhibitors must be carefully adjusted as an overdosage can cause detrimental adverse effects in non-sterile inflammation-like colitis.
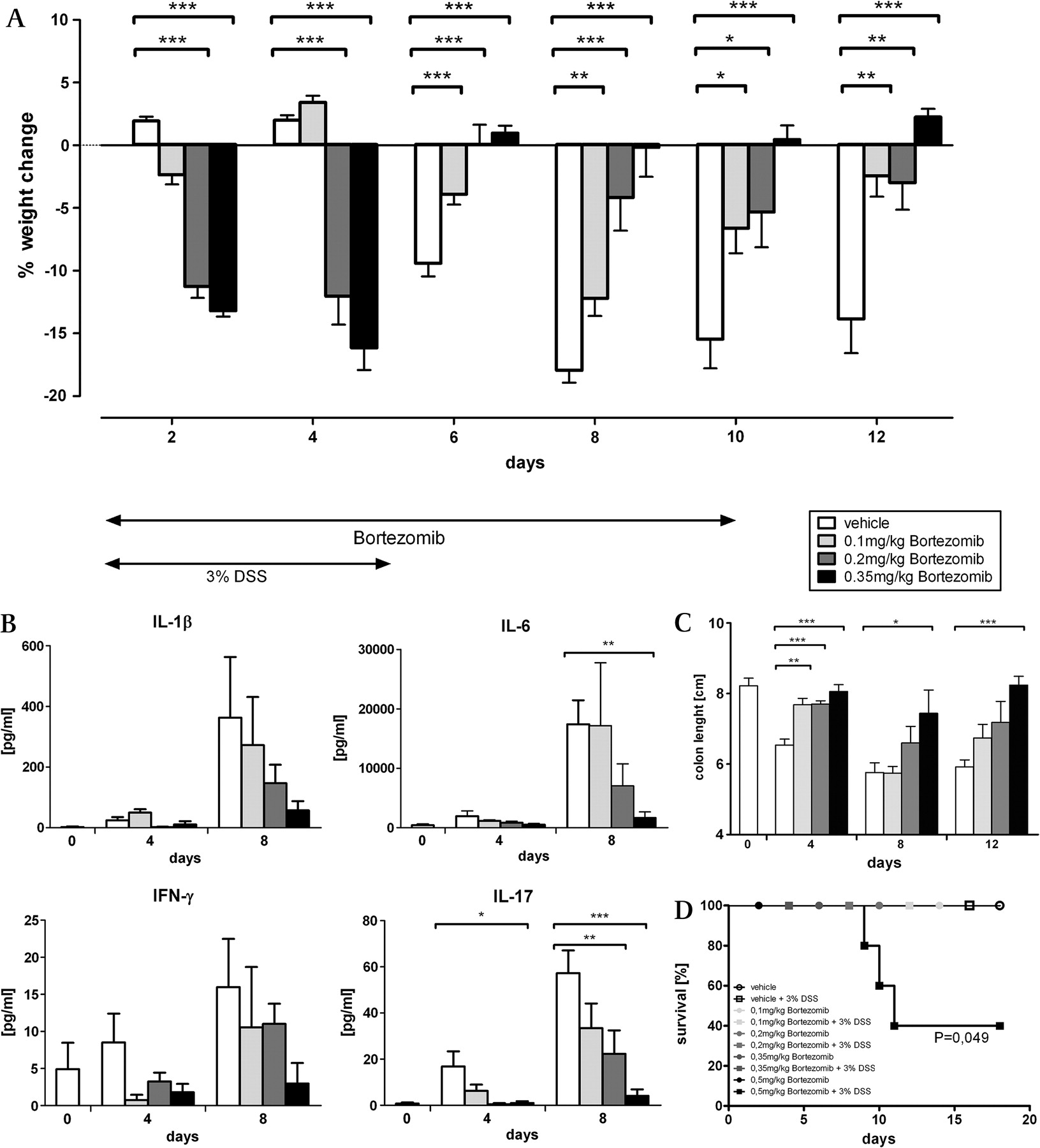
Treatment of mice with bortezomib attenuates dextran sodium sulfate (DSS)-induced colitis. 3% DSS was administered to C57Bl/6 mice in the drinking water over a period of 5 days. Mice were treated daily with a vehicle control or the indicated doses of bortezomib given intraperitoneally up to day 10. The change of weight was followed over a period of 12 days in 20 mice per group and one of two independent experiments is shown (A). The cytokine concentrations secreted by colon explants of the different treatment groups were analysed using the Bioplex bead array technology. Shown are mean values ± SEM of five mice per group and time point (B).The mean values of colon length ± SEM were determined in five mice per group and time point. One of two representative experiments is shown (C). *p<0.05; **p<0.01; ***p<0.001 Additionally, mice were treated with 3% DSS or left untreated and the indicated doses of bortezomib were administered intraperitoneally daily up to day 10. The survival of the mice (n=5) was followed daily (D). The p values were determined by the Mantel Cox log rank test.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Bortezomib treatment limits neutrophil influx in dextran sodium sulfate (DSS)-induced colitis. C57Bl/6 mice were given 3% DSS for 5 days and were treated daily with a vehicle control or the indicated doses of bortezomib given intraperitoneally up to day 10. Shown are images of H&E-stained paraffin-embedded colon sections (×20 magnification) and fluorescence images of cryosections stained for DNA (blue) and myeloperoxidase (MPO; red) (40× magnification), both taken 8 days after DSS exposure. Representative images of five mice per group are shown (A). The cytokine concentrations secreted by colon explants of the different treatment groups were analysed using the Bioplex bead array technology. (B) Shown are mean values ± SEM of five mice per group and time point. *p<0.05; **p<0.01; ***p<0.001. MCP, monocyte chemotactic protein; MIP, macrophage inflammatory protein; nd, not detectable; RANTES, regulated upon activation, normal T cell expressed and secreted.
Discussion
A central regulator of inflammation in IBD is the transcription factor NF-κB. While basal NF-κB signalling in epithelial cells is required to maintain intestinal immune homoeostasis, excessive and prolonged activation leads to chronic inflammatory disease.3 4 24 25 We have previously shown that increased proteasome activity induced by high level expression of immunosubunits mediates sustained activation of NF-κB in patients with IBD.13 Thus we wondered whether targeting these subunits is suitable to limit excessive NF-κB activation while preserving basal signalling required for intestinal immune homoeostasis. The immunosubunit LMP7 was chosen due to its high abundance in patients with CD or UC, rendering it a potential target for both diseases. Our current work underlines the role of LMP7 in driving inflammation, because the expression of many NF-κB-regulated inflammatory mediators, cytokines and chemokines was drastically reduced in DSS-treated lmp7−/− mice, resulting in markedly attenuated colitis.
Enhanced influx of neutrophils in the intestine is characteristic for the pathogenesis of IBD. Consequently, various strategies that target neutrophils have been tested in experimental colitis: (1) reduction of neutrophil influx by blocking adhesion molecules,26 neutrophil-attracting chemokines or their receptors27–29; and (2) blockade of effector functions by inhibition of matrix metalloproteases or reactive oxygen and nitrogen intermediates.30–32 These approaches revealed that reduced neutrophil activity at the site of inflammation is beneficial, while complete depletion is detrimental. In conclusion, neutrophils not only promote tissue damage but also are essential for wound healing and bacterial clearance in non-sterile inflammation.
DSS-treated lmp7−/− mice revealed reduced but not absent neutrophil influx in the colon. This prevented excessive tissue damage, while bacterial clearance and wound healing appeared to be unaffected. Thus, a balanced neutrophil response can be achieved by targeting LMP7. The neutrophil influx was also dampened by treatment with bortezomib. However, only low to intermediate doses of bortezomib were beneficial in the treatment of colitis, while high doses, which abrogate neutrophil influx, were detrimental. We assume that this immunosuppressive effect is critical for the increased mortality found with DSS treatment. Still, our results demonstrate that the neutrophil activity can be regulated with non-selective proteasome inhibitors. However, in order to avoid adverse effects the optimal dose must be carefully titrated, which reveals a potential safety risk for the use of such inhibitors during severe inflammation.
Contrasting results with non-selective proteasome inhibitors have already been reported previously. While treatment with bortezomib or MG132 ameliorated petidoglycan/polysaccharide-induced colitis in rats or spontaneous colitis in IL-10−/− mice, respectively, treatment of DSS-induced colitis with MG132 resulted in aggravated inflammation.16 18 These results might be reflected by the opposing effects we observed with different doses of bortezomib. Thus, we assume that the different outcome of these studies might depend on the various doses and treatment schedules that were used.
In contrast to non-selective inhibition, targeting of LMP7 appears to be more suitable to dampen excessive inflammation, as adverse effects such as complete blockade of neutrophil influx were never observed in lmp7−/− mice. Thus, lmp7−/− mice are still capable of controlling microbial infections.33 34 A possible explanation is that targeting a specific subunit such as LMP7 only affects a subset of proteasomes while preserving the activity of others. Various reports describe the occurrence of a large variety of proteasome subtypes with mixtures of constitutive and immunosubunits.35–37 Thus, following deletion of LMP7-containing proteasomes there is still sufficient activity of other proteasome subsets able to support basic immune functions and re-establishment of intestinal immune homoeostasis after DSS exposure. Since LMP7 is predominantly expressed in cells of the immune system as well as in inflamed tissues, targeting of this subunit will preferentially reduce NF-κB activation at the site of inflammation, thus reducing the risk of adverse effects compared with non-selective inhibition. Indeed, the bone marrow chimera experiments confirmed that LMP7 deficiency predominantly affects the initiation of an inflammatory response within the mucosal immune system, suggesting that inhibition of LMP7 is a promising approach for the treatment of IBD.
Besides innate effector mechanisms, CD4+ T cells play a dominant role in the progression of IBD. While the CD4+ T cell population in UC is predominantly of the Th2 phenotype, CD is characterised by a Th1/Th17 T cell response.1 38 In the DSS colitis model, Th1 and Th17 T cell populations expand in the inflamed colon of WT mice, resembling the phenotype of CD.23 In lmp7−/− mice, expansion of both Th1 and Th17 T cells was reduced, demonstrating that the lack of LMP7 also affects CD4+ T cell responses. Whether this is a consequence of reduced early inflammation or an intrinsic defect of CD4+ T cells cannot be discerned here. Further, it has to be investigated whether specific inhibition of LMP7 would also act on expanded and activated T cell populations present in patients with IBD.
In summary, our data demonstrate that modulation of the proteasome activity by specifically targeting LMP7-containing proteasomes substantially reduces intestinal inflammation. The advantage of targeting this specific subset of proteasomes is that the basic functionality of the proteasome system is preserved. In contrast, high-dose treatment with non-selective proteasome inhibitors, such as bortezomib, can cause detrimental adverse effects during inflammation. Thus, inactivation of specific proteasome subsets seems to be favourable in the treatment of colitis. In conclusion, we propose selective inhibition of LMP7 as a promising therapeutic approach in IBD.
Acknowledgments
We thank Petra Krienke and Dagmar Oberbeck-Müller for excellent technical assistance, and Karin Hahnke for support with the Gene Array Analysis. Further, we thank Christiane Desel and David Ermert for critically reading the manuscript.
References
Supplementary materials
web only files gut.2009.203554
Files in this Data Supplement:
Footnotes
US and TJ contributed equally to this work.
Funding This work was funded by the DFG within the projects SFB633 and SFB650.
Competing interests None.
Provenance and peer review Not commissioned; externally peer reviewed.
Linked Articles
- Digest