Article Text

Abstract
Background—Ectopic protease activation, microcirculatory changes, and leucocyte activation are the main events in the pathogenesis of acute pancreatitis. Nitric oxide (NO) is known to be a key mediator in the normal and inflamed pancreas.
Aims—To investigate the targets on which NO exerts its effect in caerulein induced pancreatitis.
Methods—Acute pancreatitis was induced in rats which additionally received either the NO synthase substrate, l-arginine; the NO donor, sodium nitroprusside; or the NO synthase inhibitor, N-nitro-l-arginine methyl ester (l-NAME). At six hours, pancreatic injury (oedema, leucocyte content, ectopic trypsinogen activation) was analysed and pancreatic oxygenation and perfusion were determined. A direct influence of NO on amylase secretion and trypsinogen activation was evaluated separately in vitro.
Results—Both NO donors reduced the grade of inflammation. l-NAME increased the severity of inflammation, while decreasing pancreatic tissue oxygenation. Although neither amylase secretion nor intracellular trypsinogen activation in caerulein stimulated pancreatic acini was influenced by either NO donors or inhibitors, both NO donors decreased intrapancreatic trypsinogen activation peptide (TAP) and pancreatic oedema in vivo, andl-NAME increased TAP.
Conclusions—NO protects against injury caused by pancreatitis in the intact animal but has no discernible effect on isolated acini. It is likely that in pancreatitis NO acts indirectly via microcirculatory changes, including inhibition of leucocyte activation and preservation of capillary perfusion.
- acute pancreatitis
- nitric oxide
- microcirculation
- leucocytes
- pancreatic secretion
Statistics from Altmetric.com
Nitric oxide (NO) has recently emerged as a biological regulatory substance in a variety of cell types including endothelium,1 macrophages,2platelets,3 and the pancreas,4 in which NO is synthesised from l-arginine by constitutive enzymes.5 Its physiological actions are mainly mediated by cyclic guanosine monophosphate (cGMP).6 In the pancreas, intracellular cGMP concentrations increase in response to certain secretagogues, such as cholecystokinin,5 and NO thereby plays an active role in pancreatic secretion.7 8 Outside the pancreatic cells NO plays a part in the control of systemic blood pressure and active dilatation of blood vessels,9 10 and is additionally involved in platelet aggregation,11leucocyte activation, and adhesion.12-14
Central events in the pathogenesis of acute pancreatitis are microcirculatory changes,15 16 leucocyte dependent inflammatory processes,17 18 and premature or ectopic pancreatic protease activation.19 20 It has recently been shown that NO has a beneficial effect on certain features of acute pancreatitis, both mild and severe.21-23 We found that NO donors decrease pancreatic oedema formation, intrapancreatic trypsinogen activation, and amylase secretion.21 These experiments did not elucidate the mechanism or site of activation of NO in pancreatitis, but the beneficial effect of inhaled NO21suggested a target in addition to the pancreatic acinar cell, possibly on leucocyte-endothelium interaction, platelet aggregation, and pancreatic perfusion. In the present study we utilised reflectance spectroscopy to evaluate in vivo the effects of NO donors and antagonists on pancreatic oxygenation in caerulein induced pancreatitis, quantitated pancreatic leucocyte infiltration by histological scoring and assay of myeloperoxidase activity, and in addition measured the conventional indexes of pancreatic injury (oedema, blood amylase levels, and ectopic trypsinogen activation). Some effects of NO on isolated acini were compared with its effects in the intact animal.
Materials and methods
in vivo studies
Animals
Experiments were performed in male Sprague-Dawley rats weighing 275–325 g. Animals were fasted overnight prior to the experiment with free access to water. Care was provided in accordance with the procedure outlined in the Guide for Care and Use of Laboratory Animals (NIH Publication 85–23, 1985). Surgical anaesthesia was induced with vaporised ether and maintained by an intramuscular injection of pentobarbital (20 mg/kg) and ketamine (40 mg/kg). Polyethylene catheters (0.5 mm internal diameter) were inserted into the right jugular vein and the left carotid artery, subcutaneously tunnelled to the back, and exited through a steel tether that allowed the animals free movement and access to water during the experiment.
Model
Animals were allocated to the control group (n=20), which received intra-arterial saline, or to the acute pancreatitis group (n=80); acute pancreatitis was induced by continuous intra-arterial infusion of caerulein (5 μg/kg/h) for six hours.24 Caerulein was reconstituted in normal saline and infused at 3 ml/kg/h.
Nitric oxide donors or inhibitors were administered simultaneously with caerulein in 60 animals of the pancreatitis group. The nitric oxide synthase substrate l-arginine (25 mg/kg bolus and subsequently 5 mg/kg/h),1 the exogenous NO donor sodium nitroprusside (SNP, 0.2 mg/kg/h),25 and the NO synthase inhibitor N-nitro-l-arginine methyl ester (l-NAME, 10 mg/kg bolus and subsequently 2.5 mg/kg/h)26 were infused intravenously over six hours in 20 animals of each subgroup.
At six hours, end point measurements of heart rate (HR), mean arterial pressure (MAP), amylase, and trypsinogen activation peptide (TAP) in serum were obtained. Ten animals of each subgroup were then sacrificed by an intra-arterial pentobarbital overdose (200 mg/kg), and the pancreas was assessed for oedema formation, ectopic trypsinogen activation, myeloperoxidase activity, and histology. In the remaining 10 animals reflectance spectroscopy of the pancreas was performed as described below.
Monitoring
MAP and HR were monitored by a thermal recorder (Mark 10–1; Western Graphetec, Irvine, California). Measurements of amylase and TAP in serum were obtained before the induction of pancreatitis (baseline) and after six hours (end point). The blood volume of each sample was replaced with double the amount of Ringer’s lactate.
Reflectance spectroscopy
In a separate set of experiments designed to evaluate end points of tissue oxygenation and oxygen consumption, pancreatic haemoglobin content and oxygen saturation of haemoglobin (ISo 2) were measured using diffuse reflectance spectroscopy as previously described.16 27 Briefly, the abdomen was opened, and a probe head (2.5 mm diameter) of a modified diode array spectrophotometer (HP 84523, Hewlett Packard, California) emitting and receiving light was placed on the pancreatic surface. Spectral changes in the reflected light were analysed by a connected personal computer (Vectra CS, Hewlett Packard, California); ISo 2 was calculated from the absorbance at 577 nm (absorbance maximum of oxygenated haemoglobin) and 555 nm (absorbance maximum of deoxygenated haemoglobin), plotted against pancreatic haemoglobin content, determined from the absorbance at the isobestic point (586 nm). Measurements were performed in 10 animals per group at five selected sites in the duodenal portion of the gland at baseline and at the end point after six hours. Data are expressed as mean (SEM) of the difference of oxyhaemoglobin or deoxyhaemoglobin content in pancreatic tissue between baseline and end point at six hours.
Myeloperoxidase activity
Excised pancreatic tissues were rinsed with saline, blotted dry, shock frozen in liquid nitrogen, and stored frozen at −80°C until thawing for determination of myeloperoxidase activity using methods previously described28-30 with minor modifications. Pancreatic tissue was homogenised in 0.1 M sodium phosphate buffer containing 0.5% hexadecyl trimethyl ammonium bromide (Sigma H5882, Sigma Chemical Co.) and 5% soybean trypsin inhibitor (Sigma T9003, Sigma Chemical Co.) before sonication and was then frozen directly on dry ice. The specimens were freeze thawed three times, and after each cycle sonication was repeated. Suspensions were then centrifuged at 20 000 g for 15 minutes and the resulting supernatant assayed. Myeloperoxidase activity was measured with a spectrophotometer (Shimazdu, UV-160, Japan) at 470 nm by mixing an aliquot (25 μl) of the supernatant with 1.0 ml of 0.1 M sodium phosphate buffer (pH 7.0) containing 0.0016 ml guaiacol (Sigma G-5502, Sigma Chemical Co.) and 0.0005% hydrogen peroxide (Sigma T9003, Sigma Chemical Co.) as substrates.
Histological assessment
The duodenal portion of the pancreas was fixed in 10% buffered formalin, embedded in paraffin wax, and one longitudinal section stained with haematoxylin and eosin. Histopathological evaluation was performed by one pathologist with expertise in pancreatic pathology who was blind to the experimental protocol. Morphometric documentation was obtained for acinar necrosis and inflammation, using a scoring system from 0 (no pathological changes) to 3 (maximal inflammatory infiltration, total necrosis of the panceas), as described by Schmidtet al.31
Oedema assessment
Pancreatic oedema was evaluated by measuring water content. A portion of the pancreas was removed immediately after sacrifice, trimmed of fat, and weighed. Pancreatic water content was determined by calculating the wet:dry weight ratio from the initial weight of the pancreas (wet weight) and its weight after incubation at 160°C for 24 hours (dry weight) as described previously.21
Trypsinogen activation peptide measurements
Ectopic trypsinogen activation has been shown within the pancreas in a variety of models of experimental pancreatic injury.20 21 30 The amount of activation, as documented by quantitation of TAP, increases with the severity of pancreatic injury32 and predicts the outcome of pancreatitis.19 For measurement of TAP concentrations in pancreatic tissue, two samples of tissue (0.1–0.3 g) were excised from the pancreas. The specimens were then immersed in 0.2 M Tris-HCl buffer (pH 7.3) containing 20 mM EDTA, immediately boiled (100°C) for 10 minutes to denature residual protease activity, and homogenised with a Brinkman Polytron (Brinkman Instruments, Westbury, New York) for 30 seconds. After centrifugation (580 g, 10 minutes, 4°C), the resulting supernatant was stored at −20°C until assayed. TAP was measured by an enzyme linked immunosorbent assay (ELISA)20 and results expressed as nmol TAP per mg pancreas. Synthetic TAP (YD4K), a conjugate of rabbit serum albumin with YD4K, and rabbit anti-TAP antiserum containing calcium independent anti-TAP antibodies were provided by Professor J Hermon-Taylor (St George’s Hospital Medical School, London, UK). Biotin-goat antirabbit IgG antibody and alkaline phosphatase labelled extravadin were purchased from Sigma Chemical Company.
Amylase measurement
Amylase activity in serum was determined according to the method of Ceska et al 33 using the Phadebas Amylase Test (Pharmacia Diagnostics, Uppsala, Sweden).
in vitro studies
Pancreatic acinar cell suspensions
For each experiment three male Wistar rats (80–120 g) were fasted overnight and sacrificed in CO2. The pancreas was quickly excised in a cold room and pancreatic acini prepared by collagenase (type CLS 4, 1000 U/ml, Worthington Biochemical Corporation, Freehold, New Jersey) digestion as described in detail previously.34After collagenase digestion the pancreatic cells were washed with medium containing HEPES 12.5 mM, NaHCO3 5.0 mM, NaCl 125 mM, KCl 5.0 mM, CaCl2 1.2 mM, KH2PO4 1.2 mM, MgSO4 1.2 mM,d-glucose 5.0 mM, aprotinin 0.01 mg/ml, soybean trypsin inhibitor 0.1 mg/ml, and bovine serum albumin 0.1% at a pH of 7.40 (all chemicals from Sigma Chemical Company). The pancreatic acinar cells were then isolated by centrifugation (three times at 100g) and filtering (polyethylene filter, mesh opening 292 μm, Spectra/Mesh, Spectrum, Houston, Texas). Cell viability was tested with the trypan blue exclusion method immediately after preparation of acini by collagenase digestion. Preparations were accepted for study only if more than 95% of the cells excluded the dye.
The cells were incubated at 37°C in 24 well cell culture plates (Falcon 3047, Becton Dickinson Labware, Lincoln Park, New Jersey) with 500 μl of cell suspension in each well, gassed with 100% oxygen. In the first study with pancreatic acinar cells, the effect of different concentrations of NO donors and inhibitors on pancreatic secretion in normal and stimulated cells was investigated. Pancreatic anini were preincubated for 15 minutes, before different dosages ofl-arginine (1, 5, 10, 50, and 100 mmol/l), SNP (1, 5, 10, 50, and 100 mmol/l), or l-NAME (1, 5, 10, 50, and 100 mmol/l) were added for 30 minutes. Saline or supraphysiological stimulation with caerulein (10−10 mmol/l) was then applied for one hour. In a second series of experiments, the influence of NO on different grades of secretory stimulation was studied. After incubation with the NO donors l-arginine (10 mmol/l) or SNP (1 mmol/l), or the NO synthase inhibitor l-NAME (10 mmol/l), or with saline (control group) for 30 minutes, pancreatic acini were exposed to different concentrations of caerulein (10−12–10−8 mmol/l) for 60 minutes. At the end of the one hour incubation period 150 μl of the medium was removed to assay for amylase and TAP in the supernatant. All measurements were performed in duplicate, and controls were run on each plate. Then 400 μl Tris (0.1 M, pH 8.9) and Triton X-100 (2%) was added for cell homogenisation and the plates were incubated on a mini shaker for 30 minutes at room temperature; amylase (total amylase) and TAP were again assayed as described above. Finally, to ascertain the relation of the observed TAP levels to the total trypsinogen pool, the acinar cells were incubated with enterokinase (1 U/ml) for one hour at 37°C to convert all trypsinogen to equimolar quantities of trypsin and TAP. The added TAP (total TAP) at this last step thus represents the trypsinogen content of the cells, as opposed to the activated trypsin. TAP levels were expressed as a function of total trypsinogen to indicate the degree of activation.
statistical analysis
Results are presented as mean (SEM). Differences between groups were compared using analysis of variance followed by Student’st test. When a normal distribution was not present (TAP), the non-parametric analysis of variance (Kruskal-Wallis test) was used for statistical analysis followed by the Mann-Whitney rank sum test. A 5% probability of type I experimental error (p<0.05) was considered statistically significant.
Results
in vivo studies
There was no difference among the five subgroups at baseline with respect to MAP or HR. Six hours after induction of pancreatitis, there was still no difference in MAP or HR between control animals and those in which pancreatitis was induced without additional administration of NO donors or inhibitors. Based on pilot studies to determine the appropiate effective doses of NO donors and NO inhibitors, the dosages were chosen such that the MAP in animals with pancreatitis was lowered approximately 10 mm Hg if l-arginine or SNP was administered, and raised 10 mm Hg by l-NAME (table 1). These changes in MAP indicate that the dosages were sufficient to produce an effect but small enough to be unlikely to influence the further course of pancreatitis.
Influence of nitric oxide on vital signs, pancreatic secretion and pancreatic histology in patients with pancreatitis
Reflectance spectroscopy
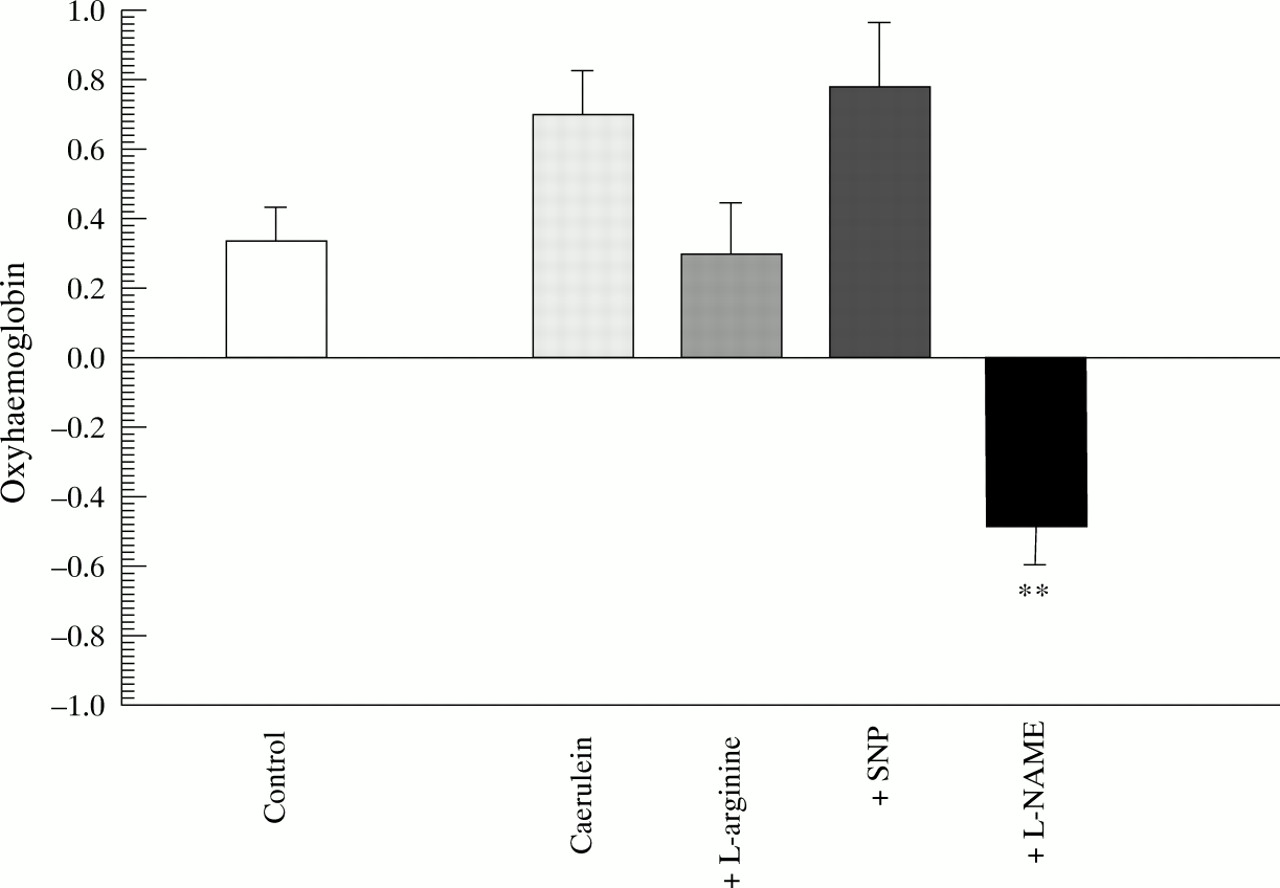
Control rats maintained their oxyhaemoglobin (fig 1) and deoxyhaemoglobin (fig 2) concentrations in pancreatic tissue over the six hour observation interval, indicating stable oxygenation and presumably stable perfusion in the experimental conditions. Similarly there were no significant changes in these parameters after six hours of caerulein induced pancreatitis. The addition ofl-arginine reduced the amount of deoxyhaemoglobin in the pancreatic tissue, while oxyhaemoglobin remained constant. The same was true for the addition of SNP, suggesting a relative increase effective pancreatic perfusion by the action of NO donors. In contrast, the addition of l-NAME led to a significant decrease in pancreatic oxygenation, as shown by a decreased oxyhaemoglobin content and increased deoxyhaemoglobin content (figs 1 and 2).

Diffuse reflectance spectroscopy. Difference in oxyhaemoglobin content in pancreatic tissue between baseline (0 hours) and end point (6 hours) measurements. **p<0.01 compared with caerulein.
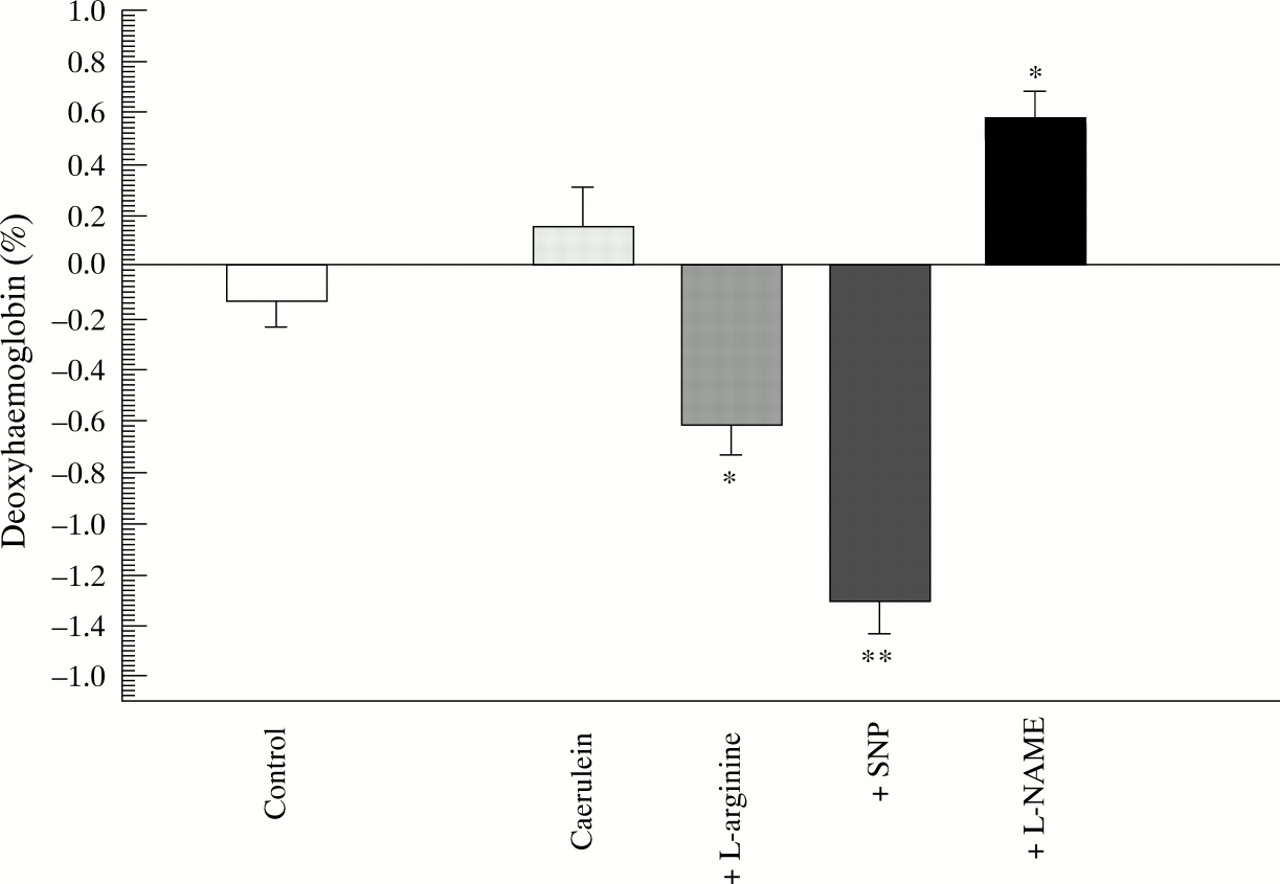
Diffuse reflectance spectroscopy. Difference in deoxyhaemoglobin content in pancreatic tissue between baseline (0 hours) and end point (6 hours) measurements. *p<0.05, **p<0.01 compared with caerulein.
Myeloperoxidase activity in pancreatic tissue
Pancreatitis led to significant leucocyte accumulation of pancreatic myeloperoxidase activity (p<0.001 compared with controls), which was reduced by both l-arginine and SNP, but greatly increased by l-NAME (fig 3).

Leucocyte infiltration of pancreatic tissue as evaluated by myeloperoxidase activity. **p<0.01 compared with caerulein; †p<0.05 compared with control.
Histological assessment
Specimens of control animals (no pancreatitis) showed normal acinar architecture. Caerulein induced pancreatitis led to obvious interstitial oedema and slight inflammation but little necrosis.l-arginine decreased inflammation and necrosis significantly, and SNP had a lesser effect in the same direction. In contrast, administration of l-NAME increased the inflammatory response, but there was no additional necrosis detectable by six hours (table 1).
Pancreatic oedema
Caerulein infusion induced gross pancreatic oedema as reflected by an increase in wet:dry ratio (table 1). The NO donorsl-arginine and SNP greatly decreased the water content; the NO synthase inhibitor l-NAME also reduced water content.
Amylase and TAP in serum
There was significant hyperamylasaemia in animals with pancreatitis induced by supramaximal caerulein stimulation. Compared with caerulein alone, l-arginine and SNP significantly decreased the hyperamylasaemia (p<0.001); l-NAME had no effect (table 1). TAP in serum (TAP at six hours minus TAP at baseline) was significantly increased in animals with pancreatitis in comparison with control animals (table 1). The addition of NO promoters significantly decreased TAP in serum, while the NO synthase inhibitor had no significant effect on serum TAP values compared with animals with pancreatitis.
Pancreatic tissue TAP
Caerulein induced pancreatitis increased TAP values in the pancreatic tissue homogenates significantly (p<0.001). Bothl-arginine and SNP reduced trypsinogen activation whilel-NAME increased the tissue TAP (table 1).
in vitro studies
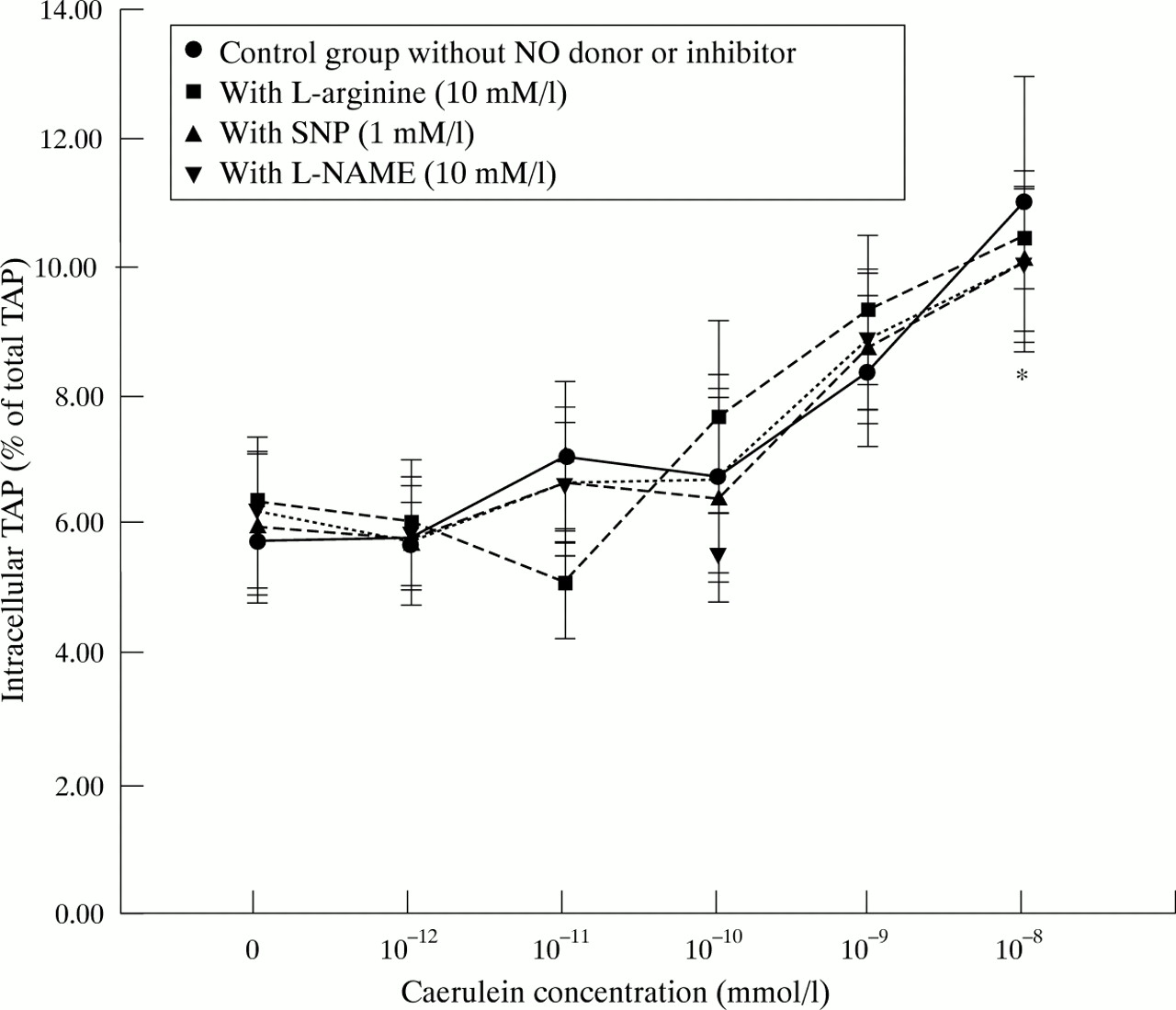
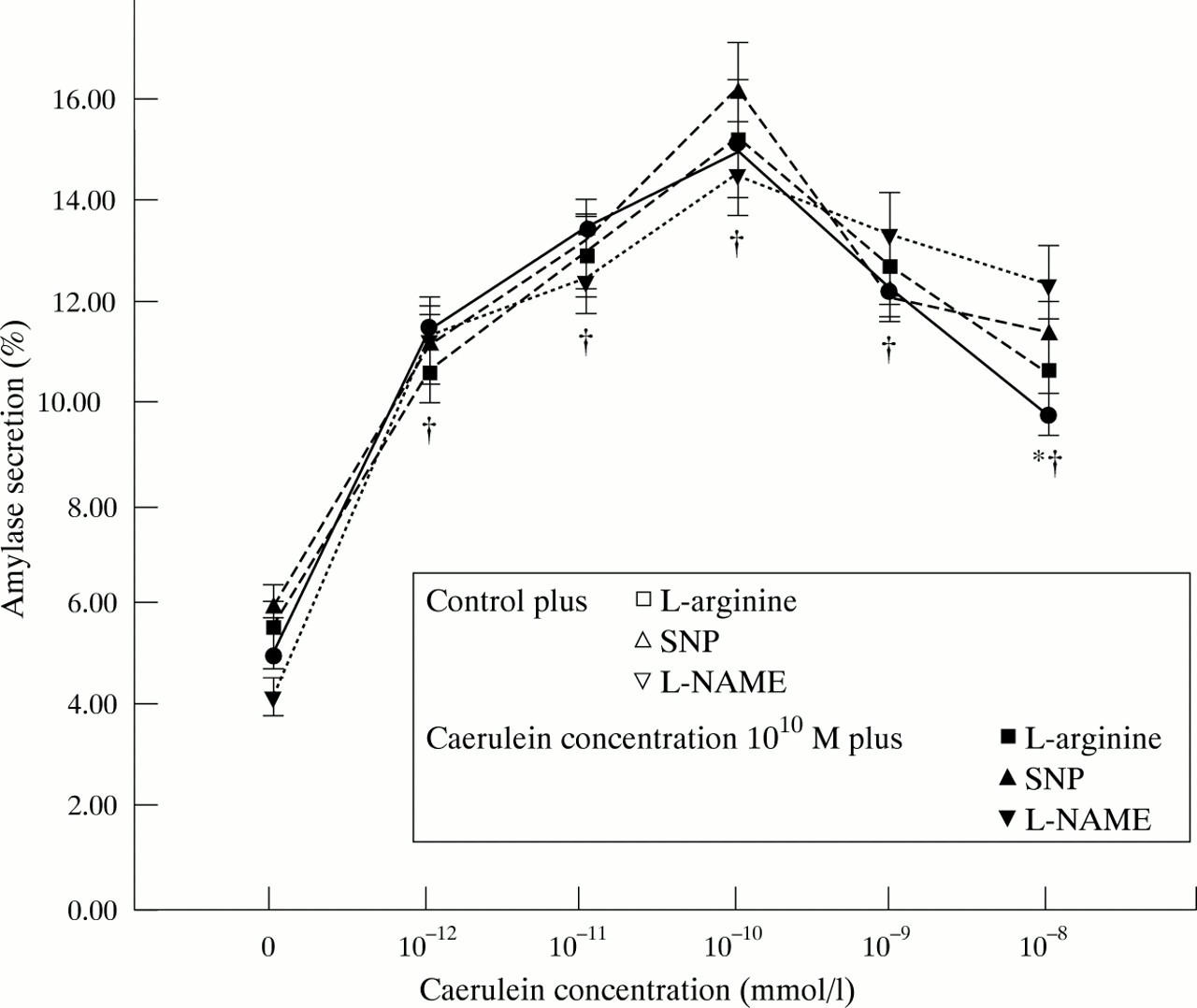
Amylase secretion was notably increased when acinar cells were stimulated with a supraphysiological concentration of caerulein (10−10 mmol/l) compared with unstimulated cells (p<0.001; fig 4) and intracellular trypsinogen activation was slightly increased by caerulein stimulation (fig 5). The typical biphasic shape of the caerulein dose-response curve of amylase secretion, which indicates the functional viability of the acinar cell preparation, was not altered by l-arginine (10 mmol/l), SNP (1 mmol/l), orl-NAME (10 mmol/l) (fig 6). There was no effect on amylase secretion from the addition of different concentrations of NO donors or inhibitors to either unstimulated or stimulated cells (fig 4). There was no influence of l-arginine, SNP, or l-NAME on intracellular TAP at any concentration tested (data not shown) or at any level of secretory stimulation (fig5).

Influence of NO donors and inhibitors on amylase secretion of unstimulated versus secretory stimulated pancreatic acini.

Influence of NO donors and inhibitors on intracellular TAP formation in pancreatic acinar cells (as a measurement of intracellular trypsinogen activation) in response to different grades of secretory stimuli with caerulein. *p<0.05 compared with control group and with caerulein 10−12 M.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Influence of NO donors and inhibitors on amylase secretion by pancreatic acinar cells in response to different grades of secretory stimuli with caerulein.*p<0.05 compared with caerulein 10−12 M; †p<0.05 compared with control.
Discussion
We have previously shown that NO donors in vivo may confer some protection in acute experimental pancreatitis, both mild and severe, by reducing oedema formation, hyperamylasaemia, and ectopic trypsinogen activation and that inhibition of NO synthase produces the obverse deleterious effects. Other studies have been in accord,7 21 23 25 35 but disagreement also exists.22 36 37 None of these previous investigations have shed light on the mode or site of action of NO, whether it be directly on the acinar cell or indirectly mediated through microcirculatory changes or other remote mechanisms. In this study we analysed the influence of l-arginine, SNP, andl-NAME on the development of pancreatitis both in vivo and in vitro. At the cellular level we utilised parameters to indicate secretory function and premature intracellular protease activation, and in vivo we measured leucocyte accumulation, tissue oxygenation, and protease activation. These studies were designed to distinguish between effects of NO at the cellular level and those which are the vector of systemic actions. The present study supports our previous findings that NO plays a protective role in the evolution of acute pancreatitis in that all the measures of injury were ameliorated by the administration of l-arginine or SNP and worsened by l-NAME. The new observations of greatest importance are that NO donors reduce the numbers of leucocytes accumulating in the pancreas in pancreatitis and improve tissue oxyhaemoglobin concentrations, while inhibition of NO synthase is harmful to these same parameters.
Pancreatic perfusion and tissue oxygenation have been considered to be key factors in the pathogenesis and progression of pancreatitis,15 16 and the benefits of NO therefore appear to be particularly relevant. Recent reports about NO, also described as endothelium derived relaxing factor,10describe cytoprotective effects through increase of organ blood flow in several organs including the intestine,38liver,39 and pancreas.35 40 Progression of pancreatitis may well be dependent on the balance between protectors of microcirculatory perfusion and those agents which serve to produce the reduction in capillary flow and perfusion which characterise severe necrotising pancreatitis.15 16 We suggest that NO plays a significant role in this balance. Our diffuse reflectance spectroscopy data show that the addition of NO reduces the pancreatic tissue content of deoxyhaemoglobin, while NO synthase inhibition increases deoxyhaemoglobin and diminishes oxyhaemoglobin in caerulein induced pancreatitis. In our previous studies these changes correlated with improved or impaired tissue blood flow respectively.16 27We infer from the present observations that the effects of nitric oxide are mediated in part by changes in effective microcirculation.
Leucocyte adhesion to the endothelium in postcapillary venules, progressive accumulation, and infiltration into pancreatic tissue is a prominent and constant feature of all models of acute pancreatitis.17 30 41 Firstly, NO is known to be a protective modulator against leucocyte adherence,11 12and impairment of endogenous NO generation enhances leucocyte adhesion and migration into tissues.6 12 This phenomenon is exemplified in the present study by the increased myeloperoxidase activity in animals treated with l-NAME and by the complementary finding of decreased myeloperoxidase produced byl-arginine and SNP. Leucocytes may initiate or potentiate cell damage by several mechanisms, including free oxygen radical and cytokine production.42 43 Hypoxic tissue and leucocytes are the two principal sources of free radicals which are generated early in the course of pancreatitis and lead to exacerbation of pancreatic cell injury.43 Secondly, NO functions as a superoxide scavenger and reduces superoxide anion production in neutrophils.14 44 Finally, NO is thought to maintain vascular integrity in the leucocyte independent early increase in microvascular permeability as well as in the delayed vascular dysfunction that is probably mediated by toxic secretions from adherent leucocytes.13 45 The reduction of caerulein induced pancreatic oedema by both l-arginine and SNP may therefore be a consequence of either or both of these mechanisms of NO action. It is paradoxical at first inspection that l-NAME also led to some reduction in oedema formation, but that effect may actually be due to a reduction in pancreatic perfusion, representing a worsening of the pancreatitis, as more severe forms of pancreatitis are characterised by a massive increase in inflammation with less pronounced oedema formation.21 30
In this study pancreatic amylase secretion and intracellular trypsinogen activation in acinar cells were not altered by the addition of either NO donors or inhibitors in vitro, whether in resting, stimulated, or hyperstimulated cell preparations. Hyperstimulation of acini with caerulein can be considered a “cellular pancreatitis model”,46 so that the absence of NO effects in vitro is further evidence that the alterations in amylase and TAP in vivo are secondary effects rather than primary effects on the acinar cell. These findings are consistent with the results of Moleroet al,25 who showed no alteration in basal and caerulein stimulated amylase release after the addition of l-NAME, and with those of Konturek and coworkers7 using NG-nitro-l-arginine, a different NO synthase inhibitor. However, Wrenn and coworkers47 reported that treatment of acini with l-arginine increased nitrite, cGMP, and amylase release, which were prevented by NO synthase inhibitors, and that NG-nitro-l-arginine blocked carbachol induced amylase release. Whereas we used dispersed preparations of isolated acinar cells, Wrenn et al used cell clusters of 20–30 acinar cells, which are known to be more responsive to secretory stimulation. As neurones and microvascular endothelium may also have been included in those preparations, the effects of NO on pancreatic exocrine function in the study by Wrennet al may be the result of paracrine rather than intracellular action. This hypothesis is supported by the observation that NO synthase activity, identified by immunohistochemical staining for nicotinamide adenine dinucleotide phosphate diaphorase, has been found in nerve fibres, nerve bodies, and the vascular endothelium, but not within the acinar cells in the pancreas of several mammalian species, including the rat and human.4
Our findings suggest that the primary effects of NO during acute pancreatitis seem to be via influences on pancreatic perfusion and on leucocyte-endothelium interaction. While reduced microcirculatory flow has been shown to be characteristic of severe pancreatitis,15 16 it is possible that the increased numbers of leucocytes adhering to pancreatic endothelium and becoming activated at those sites precedes and leads to the perfusion deficits. This order of events is consistent with our finding of an increased presence of leucocytes even in mild caerulein induced pancreatitis in which capillary perfusion is still normal or even high.16 40
Although NO and its action on leucocytes in pancreatitis appear to be beneficial under the conditions of this study, there may be a paradoxical dose-response phenomenon. Morikawa et al found that NO could either enhance or inhibit oxygen free radical generation in polymorphonuclear leucocytes at different concentrations,14 and high levels of NO produced by macrophages and lymphocytes can apparently cause tissue damage.6 These observations may explain how NO donors rather than NO inhibitors could make pancreatitis worse22or how cytotoxicity from NO may participate in the lung injury associated with pancreatitis.36
In summary, the present study reconfirms that NO has a positive beneficial effect in acute oedematous pancreatitis and indicates that this action is mediated by: (1) reduction in leucocyte adhesion and subsequent tissue infiltration; and (2) modulation of pancreatic microcirculation and tissue oxygenation. Direct effects of NO on pancreatic acinar cells, including regulation of pancreatic secretion, are likely to play only a minor role in the pathogenesis of this protective mechanism.
Acknowledgments
Jens Werner is supported by the Deutsche Forschungsgemeinschaft, grant We2008/1–1 and 1–2.