Article Text

Abstract
BACKGROUND Impairment of intestinal barrier function occurs under a variety of inflammatory conditions and is mediated at least in part by interferon γ (IFN-γ) induced nitric oxide (NO) production. Previous in vivo studies have shown that systemic lipopolysaccharide treatment caused an induction of the rat inducible nitric oxide synthase (iNOS) mRNA primarily in villus cells, rather than in undifferentiated crypt cells.
AIMS To examine iNOS induction by IFN-γ in vitro as a function of enterocyte differentiation.
METHODS Preconfluent and postconfluent Caco-2 cells were treated with IFN-γ in the presence or absence of various inhibitors. Northern analyses were performed to assess the magnitude of iNOS mRNA induction. IFN-γ receptor mRNA and protein levels were determined.
RESULTS iNOS mRNA induction by IFN-γ occurred at two hours and was not blocked by cycloheximide, indicating that it is an immediate early response. iNOS induction and nitrite/nitrate increases were inhibited by dexamethasone and pyrrolidine dithiocarbamate, supporting an important role for the NF-κB transcription factor in this process. The stimulated iNOS induction was seen almost exclusively under conditions of cellular differentiation—that is, in postconfluent Caco-2 cells. This increased IFN-γ responsiveness seen in postconfluent Caco-2 cells correlated with an increased expression of IFN-γ receptor, whereas T84 and HT-29 cells did not show any significant alterations in either iNOS induction or IFN-γ receptor levels as a function of postconfluent growth.
CONCLUSIONS With regard to iNOS mRNA induction, IFN-γ responsiveness is acquired during Caco-2 cell differentiation, perhaps related to an increase in the numbers of IFN-γ receptors.
- interferon γ receptor
- nitric oxide synthase
- intestinal epithelium
- inflammation
Abbreviations
- CHX
- cycloheximide
- DEX
- dexamethasone
- IFN-γ
- interferon γ
- iNOS
- inducible nitric oxide synthase
- LPS
- lipopolysaccharide
- NF-κB
- nuclear factor κB
- NO
- nitric oxide
- NO2−
- nitrite
- NO3−
- nitrate
- NOS
- nitric oxide synthase
- PDTC
- pyrrolidine dithiocarbamate
- STAT
- signal transducers and activators of transcription
- TNF-α
- tumour necrosis factor α
Statistics from Altmetric.com
The mammalian small intestine is lined by a simple columnar epithelium which forms finger-like villi that extend into the lumen and provide a large surface area for absorptive functions. The cells that comprise the intestinal epithelium arise from a proliferative zone of undifferentiated stem cells within the crypts. Enterocyte differentiation along the crypt−villus axis results from the transcriptional activation of a variety of cell specific genes, most notably the brush border enzymes and transporters responsible for nutrient digestion and absorption.1 The intestinal epithelium serves an important role as an absorptive tissue, permitting movement from the lumen into the systemic compartment of water, electrolytes, and nutrients. The intestinal epithelium also performs a vital barrier function of limiting systemic contamination by potentially toxic compounds and/or pathogenic microorganisms normally present within the lumen of the gut.2 Inflammation, whether generalised (as in endotoxaemia or thermal injuries) or localised to the gut (as in Crohn’s disease or ulcerative colitis), has been associated, both in humans2-4 and experimental animals,5 with an increase in intestinal permeability to various hydrophilic probes, a manifestation of gut barrier dysfunction.
The mechanism(s) responsible for intestinal hyperpermeability induced by inflammation remains to be completely elucidated. Nevertheless, it has been shown that the permeability of cultured enterocytic monolayers grown on permeable supports is increased following incubation with certain proinflammatory cytokines, such as tumour necrosis factor α (TNF-α)6 and interferon γ (IFN-γ).7 ,8 The increase in permeability of Caco-2BBe monolayers induced by IFN-γ seems to be mediated, at least in part, by the pluripotent signalling and effector molecule, nitric oxide (NO), as the induced hyperpermeability is ameliorated by specific inhibitors that act on the enzyme responsible for NO production.9 Furthermore, we have shown that NO causes dilatation of intercellular tight junctions leading to the increased permeability in Caco-2BBemonolayers.10
NO is derived from the amino acid l-arginine via a complex five electron redox reaction catalysed by one of three isoforms of the enzyme, nitric oxide synthase (NOS). The isoform NOS-1 is primarily expressed in neural cells, including the plexuses present within the intestinal wall, and NOS-3 is primarily expressed in endothelial cells.11 Whereas both NOS-1 and NOS-3 are constitutively expressed, a third isoform, NOS-2, also commonly referred to as inducible nitric oxide synthase (iNOS), is typically expressed only after induction by proinflammatory cytokines and/or lipopolysaccharide (LPS).11 In a wide variety of inflammatory conditions, there is a notable biosynthesis of large quantities of NO which is due to the increased expression of iNOS.12
Previous in vivo studies from our laboratory5 and others13 have shown that iNOS induction occurs in the rat small intestine and colon following systemic administration of LPS. The expression of iNOS mRNA within the gut, however, appears to be differentially regulated along both the longitudinal and crypt−villus axes of the intestine. For example, in rats challenged with LPS, iNOS mRNA levels are greatest in the ileum compared with other regions of the small or large intestine. Furthermore, within the ileum, iNOS mRNA levels are higher in well differentiated villus cells than in less differentiated crypt cells.14 Prompted by these observations, we sought to delineate further the relation between enterocyte differentiation and iNOS expression, using an in vitro model system, Caco-2BBe cells. A preliminary report of these data has been published in abstract form.15
Materials and methods
CELL CULTURE
Caco-2BBe cells (provided by Dr M Neutra, Children’s Hospital, Boston, Massachusetts, USA) were cultured in Dulbecco’s modified Eagle’s medium (DMEM) containing 25 mM glucose supplemented with 10% fetal bovine serum, 2 mM glutamine, 100 U/ml penicillin, 0.1 mg/ml streptomycin, 24 mM sodium bicarbonate, 1% non-essential amino acids, 10 mM HEPES, and 10 μg/ml transferrin in 160 cm2plastic flasks and were maintained at 37°C under an atmosphere of 5% CO2 in air. T84 and HT-29 cells were purchased from American Type Culture Collection (ATCC) and cultured in DMEM as recommended. The media were changed every three days and the cells were separated via trypsinisation when they reached confluence. Experiments were generally performed on cells at three weeks postconfluence, except where stated otherwise. Media were changed 24 hours prior to the start of each experiment. The cells were treated with various concentrations of human recombinant IFN-γ (Pharmingen), and in some cases either cycloheximide (10 μg/ml), dexamethasone (1 μM), or pyrrolidine dithiocarbamate (PDTC, 100 μM), as indicated.
MEASUREMENT OF NO2−/NO3−CONCENTRATIONS
NO production was determined by measuring the stable oxidation products, NO2 −/NO3 −, in samples of media. Samples were incubated with copper plated cadmium filings in order to reduce nitrate to nitrite. Total nitrite concentrations were measured spectrophotometrically using the Griess reaction.16 Absorbance values were compared with a linear standard curve constructed using sodium nitrite standards.
NORTHERN BLOT ANALYSES
Total RNA was extracted using the guanidium thiocyanate method.17 Northern blot analyses were performed by loading 20 μg total RNA of each sample onto an agarose/formaldehyde gel, separating through electrophoresis, transferring to nitrocellulose membranes, and baking for two hours at 80°C. Examination of ethidium bromide stained gels verified equal loading of RNA in each lane. Complementary DNA probes were 32P labelled by the random primer method,18 typically to a specific activity of 5 × 108 cpm/μg DNA. The iNOS probe is a 1.2 kb (EcoRI/BamHI) fragment derived from the human cDNA clone and provided by Dr D A Geller (University of Pittsburgh Medical School).19 The sucrase-isomaltase probe was provided by Dr P G Traber (University of Pennsylvania Medical School).20 The β actin probe is a 1.0 kb Pst1 fragment derived from a mouse β actin cDNA.21 The IFN-γ receptor and c-fos probes were derived from the human cDNAs and obtained from ATCC. Hybridisation was performed in 5× standard saline citrate (SCC)/50% (vol/vol) formamide/1% (wt/vol) sodium dodecyl sulphate (SDS) at 42°C. Washings were performed using 2× SSC/0.1% SDS at 50°C.
WESTERN BLOT ANALYSES
Cells were lysed and rinsed in phosphate buffered saline (PBS). Lysis buffer consists of 150 mM NaCl, 20 mM HEPES, 1 mM EDTA, 1 mM DTT, 0.01% SDS, 1% NP-40, and 1× cocktail protease inhibitor (Boerhinger). The buffer was added to the cells and placed on ice for 10 minutes. All lysates were then centrifuged for five minutes at 4°C, the supernatant collected, and stored at −70°C as whole cell protein lysate. The protein concentration was measured spectrophotometrically using protein standards (Sigma). Loading of 50 μg protein was performed after addition of equivalent amount of Laemmli buffer and boiling for five minutes. SDS-polyacrylamide gel electrophoresis was performed using 10% resolving gel (Bio-Rad). Proteins were electrophoretically transferred to a nitrocellulose membrane in Tris-glycine buffer. Filters were incubated overnight with PBS/5% non-fat milk at 4°C. After rinsing with PBS/0.1% Tween 20 the membrane was incubated for one hour with rabbit polyclonal anti-IFN-γ receptor β subunit antibody at 1/500 dilution (Santa Cruz), followed by washes of PBS/0.1% Tween 20 and incubation with a secondary antibody, a horseradish peroxidase conjugated goat antirabbit antibody at 1/5000 for one hour (Santa Cruz). After further washes the immune complexes were visualised using the ECL detection system.
STATISTICAL ANALYSES
For the nitrite/nitrate measurements (n=3), results are expressed as mean (SEM). Comparisons were made using the Student’s unpairedt test; p<0.05 was considered statistically significant. Each northern and western blot experiment was repeated at least three times in order to verify reproducibility.
Results
iNOS EXPRESSION IN Caco-2BBeCELLS
When cultured in the absence of IFN-γ, Caco-2BBecells express little iNOS mRNA. However, after stimulation with IFN-γ (1000 U/ml), iNOS mRNA levels became apparent by two hours (fig 1), peaked at 36 hours, and decreased by 48 hours. The magnitude of iNOS mRNA induction rose steadily as a function of IFN-γ concentration (0–1000 U/ml; data not shown).

Time course for IFN-γ induced iNOS mRNA expression in Caco-2BBe cells. Cells (three weeks postconfluent) were treated with IFN-γ (1000 U/ml) for the indicated times. Total cellular RNA (20 μg) was hybridised using both the iNOS and β actin probes.
EFFECT OF VARIOUS PHARMACOLOGICAL AGENTS ON IFN-γ INDUCED iNOS mRNA EXPRESSION
Caco-2BBe cells were pretreated with dexamethasone (DEX), cycloheximide (CHX), or pyrrolidine dithiocarbamate (PDTC) for 30 minutes prior to being incubated for six hours with IFN-γ. As seen in fig 2A, DEX inhibited iNOS mRNA induction by IFN-γ. In order to determine whether de novo protein synthesis is necessary for IFN-γ induced iNOS mRNA expression, cells were cotreated with CHX (10 μg/ml) (fig 2A). CHX did not inhibit IFN-γ induced iNOS mRNA expression; the combination of IFN-γ and CHX resulted in a greater upregulation of iNOS mRNA expression than was observed following treatment with IFN-γ alone. For control purposes, a large increase in c-fos expression was used to verify the effects of CHX. CHX treatment alone resulted in an increase in iNOS expression, further indicating that iNOS is an “immediate early response” gene in this context.

Effect of inhibitors on iNOS mRNA induction by IFN-γ. (A) Caco-2BBe cells (three weeks postconfluent) were treated with 1 μM dexamethasone (DEX) or 10 μg/ml cycloheximide (CHX) alone and in combination with IFN-γ (1000 U/ml) for six hours. (B) Caco-2BBe cells were treated with IFN-γ (1000 U/ml) and pyrrolidine dithiocarbamate (PDTC) (100 μM) for six hours. Northern analyses were performed using total cellular RNA (20 μg) and iNOS, actin, and c-fos cDNA probes. (C) Nitrite/nitrate measurements (n=3) were performed on samples of media after 48 hours of treatment with IFN-γ alone or in combination with CHX, DEX, or PDTC. ***p<0.001.
The potent antioxidant, PDTC, has been widely used to inhibit activation of genes by the transcription factor, NF-κB.22 Accordingly, we evaluated the effect of PDTC (100 μM) on IFN-γ induced iNOS mRNA expression (fig 2B). PDTC treatment alone had no effect on iNOS mRNA induction (data not shown). The cotreatment of cells with IFN-γ and PDTC almost completely blocked the induction of iNOS mRNA expression that was seen with IFN-γ alone, supporting a role for NF-κB in the induction process.23
In order to determine iNOS enzymatic activity, nitrite and nitrate (NO2 −/NO3 −) concentrations were measured on samples of media from Caco-2BBe cells after 48 hours of treatment with the pharmacological agents CHX, DEX, and PDTC, in combination with IFN-γ and compared with IFN-γ alone (fig 2C). NO biosynthesis tends to lag behind the increased expression of iNOS mRNA because of the time required for translation of mRNA into a functional protein. The large increase in NO2 −/NO3 − levels in IFN-γ treated cells indicates that the iNOS mRNA is translated into a functional protein. As expected, CHX inhibits the actual synthesis of the iNOS enzyme despite the superinduction seen at the mRNA level. In contrast, the low NO2 −/NO3 − levels seen with DEX or PDTC cotreatment support the inhibitory action at the mRNA level as shown earlier.
EFFECT OF DIFFERENTIATION ON iNOS GENE EXPRESSION
To examine the effects of cellular differentiation on IFN-γ induced iNOS mRNA expression, Caco-2BBe cells were incubated with IFN-γ under preconfluent conditions (approximately 80% confluence) and at three weeks after attainment of confluence (fig3). Minimal iNOS induction occurred in the preconfluent cells, whereas iNOS mRNA expression was very high in the postconfluent cells. The high levels of the brush border sucrase-isomaltase mRNA in postconfluent Caco-2BBe cells verified their state of differentiation.
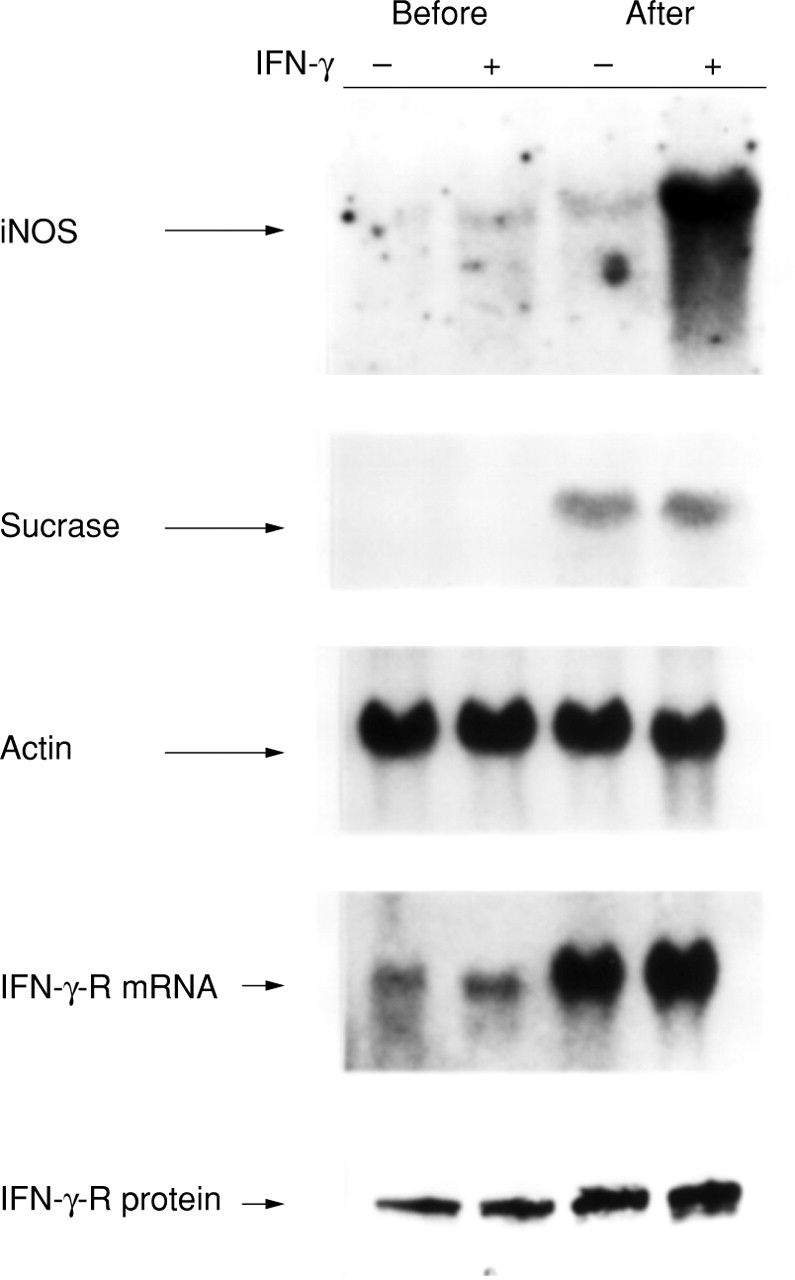
Effect of cellular differentiation on iNOS mRNA induction by IFN-γ. Caco-2BBe cells, before confluence and three weeks after confluence, were examined in untreated conditions and after six hours treatment with IFN-γ (1000 U/ml). Depicted is a representative northern blot probed with the iNOS, sucrase-isomaltase (sucrase), IFN-γ receptor (IFN-γ-R), and β actin cDNA probes. Western analyses for similarly treated samples were performed for the IFN-γ-R protein.
We hypothesised that one reason for the greater induction of iNOS mRNA by IFN-γ in postconfluent cells might be increased expression of the IFN-γ cell surface receptor. Indeed, we found that the IFN-γ receptor mRNA levels were quite low in preconfluent Caco-2BBe cells, but increased as a function of differentiation. Similarly, western blot analyses revealed that the relative amounts of IFN-γ receptor protein were higher in postconfluent compared with preconfluent cells. Thus, the IFN-γ responsiveness seen in the differentiated cells correlates with increased IFN-γ receptor mRNA and protein levels.
EFFECT OF DIFFERENTIATION ON IFN-γ RECEPTOR GENE EXPRESSION
We next examined the time course of IFN-γ receptor induction as a function of enterocyte differentiation. As seen in fig 4, there was a gradual increase in IFN-γ receptor mRNA expression as the cells became more postconfluent. The IFN-γ receptor mRNA levels appeared to reach maximal expression at four weeks after the attainment of confluence. The mRNA for sucrase-isomaltase was also induced in postconfluent Caco-2BBe cells, but induction of this differentiation marker occurred earlier than that of the IFN-γ receptor.

Effect of differentiation on IFN-γ receptor mRNA expression. Caco-2BBe cells were harvested at various times of confluence (preconfluence, and one and four weeks postconfluence) in untreated conditions. Northern analyses were performed with IFN-γ receptor (IFN-γ-R), sucrase−isomaltase (surase) and β actin cDNA probes.
COMPARISON OF IFN-γ RESPONSIVENESS IN THREE COLON CARCINOMA CELL LINES
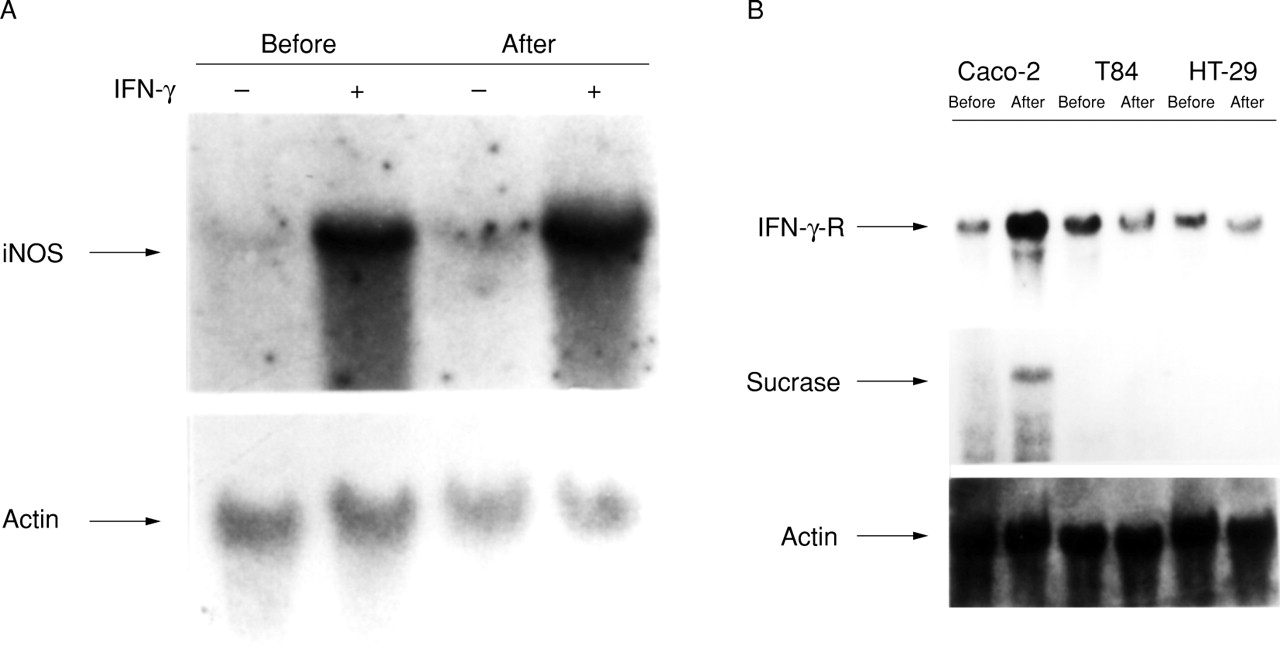
To establish further a link between the state of cellular differentiation and the ability of IFN-γ to induce iNOS mRNA expression in enterocytes, we compared three different cell lines. Caco-2BBe, T84, and HT-29 cell lines all originate from human colon carcinomas, but T84 and HT-29 cells differ from Caco-2 cells in that postconfluent growth is not accompanied by evidence of further differentiation.24 Unlike Caco-2BBecells which exhibit induction of iNOS mRNA by IFN-γ in postconfluent cells only, T84 cells respond to treatment with IFN-γ by inducing iNOS gene expression similarly in preconfluent and postconfluent states (fig 5A).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Comparison of IFN-γ responsiveness in three colon carcinoma cell lines. (A) T84 cells, preconfluent (before) and four weeks postconfluent (after), were examined in untreated conditions (−) and after six hours treatment with 1000 U/ml IFN-γ (+). Northern analyses were performed using the iNOS and β actin probes. (B) Caco-2BBe, T84, and HT-29 cells in the preconfluent (before) and postconfluent (after) states were compared in untreated conditions. Northern analyses were performed using IFN-γ receptor (IFN-γ-R), sucrase−isomaltase (sucrase), and β actin probes.
The expression of the IFN-γ receptor mRNA was then examined in preconfluent and postconfluent T84 and HT-29 cells, and compared with Caco-2BBe cells (fig 5B). Sucrase-isomaltase mRNA expression was used to confirm differentiation in postconfluent Caco-2BBe cells, but was not seen in the other cells, consistent with the fact that T84 and HT-29 cells do not differentiate as a result of postconfluent growth. In contrast to the Caco-2BBe cells, IFN-γ receptor levels in T84 and HT-29 cells were unchanged between preconfluent and postconfluent states.
Discussion
The present studies focused on IFN-γ mediated induction of iNOS and have broad implications with respect to the interaction between inflammatory cells and the gut epithelium. The effects of inflammatory cell derived cytokines on cultured intestinal epithelial cells have been well documented. Deem et al showed that activated human lamina propria T lymphocytes produce IFN-γ and TNF-α, which act in concert to kill HT-29 cells.25 The effects of IFN-γ on enterocytes may have important clinical relevance, as this cytokine has been implicated in a variety of inflammatory conditions, including ulcerative colitis, Crohn’s disease, and HIV related enterocolitis.26 Simpsonet al showed that the degree of intestinal inflammation in mice correlates with the ability of T cells to produce IFN-γ and that this proinflammatory cytokine is responsible for cell mediated tissue damage.27 Several cytokines have been shown to increase the permeability of cultured enterocytic monolayers, including IFN-γ,7 ,8 interleukin 4,28 and TNF-α.6 In previous studies, we have examined the effects of IFN-γ on the permeability of Caco-2BBe cells and found that NO mediates the induced hyperpermeability.9
The direct effects of NO on intestinal mucosal integrity are complex, as it has been shown to be either detrimental or beneficial, depending on the experimental conditions. Under both physiological and pathophysiological conditions, NO can preserve mucosal barrier function by maintaining mucosal perfusion, inhibiting neutrophil adhesion to the endothelial surfaces of the microvasculature, or inhibiting the release of other mediators, notably histamine and platelet activating factor from mast cells.29 Hoffman et al found constitutive expression of iNOS mRNA in normal unstimulated epithelial cells in the ileum of mice, suggesting a possible role for NO in intestinal homoeostasis.30However, when enterocytes are exposed to large quantities of NO for an extended period of time, mucosal damage results. For example, Teppermanet al documented that injecting rats with LPS results in diminished viability of enterocytes and colonocytes, a phenomenon which was largely blocked when NO biosynthesis was pharmacologically inhibited.31 ,32
Studies performed by our group indicate that NO causes hyperpermeability in Caco-2BBe monolayers by depleting intracellular ATP concentrations and loosening tight junctions.10 Furthermore, the increase in epithelial permeability induced by IFN-γ in vitro can be minimised by blocking NO production.9 Both human and rat intestinal epithelial cells express iNOS when appropriately stimulated. We5 and others33 have shown that administration of LPS to rats results in derangements in intestinal barrier function, an effect that is largely prevented by pharmacological antagonists of iNOS enzymatic activity. We have also shown that following systemic administration of LPS to rats, intestinal villus cells express higher levels of iNOS mRNA than crypt cells.14 The present in vitro studies extend these previous observations, and contribute to our understanding of how the process of enterocytic differentiation along the crypt−villus axis affects the ability of the intestinal epithelium to respond to IFN-γ. The fact that the magnitude of iNOS induction is dramatically increased in postconfluent (differentiated) Caco-2BBe cells is consistent with our previous in vivo study,14 as well as the work of Tepperman et al, who showed greater iNOS enzyme activity in the villus compartment.31Taken together, these observations may provide important insights into the mechanisms leading to gut mucosal dysfunction in various pathological conditions.
In the present studies, we found that iNOS mRNA was induced by IFN-γ at two hours and not blocked by the protein synthesis inhibitor, CHX, thus satisfying the criteria of an immediate early response gene.34 As is often seen with early response genes, we found that coincubation with CHX not only failed to block induction, but rather resulted in a superinduction of iNOS mRNA expression. These data are in contrast to the results reported by Melilloet al, who found that CHX blocked iNOS induction in a macrophage cell line.35 Thus, in Caco-2BBe cells, it appears that one or more transcription factors already present within the enterocytes are capable of activating the iNOS gene.
Our results with DEX confirm and extend previous work showing the inhibitory effects of glucocorticoids on iNOS activation within intestinal epithelial cells. Salzman et alhave shown that DEX partially blocked iNOS gene expression in intestinal DLD-1 cells.36 Furthermore, Teppermanet al have shown that DEX treatment inhibited both the regulation of iNOS enzymatic activity and the intestinal epithelial cytopathic effects of systemic LPS administration in rats.31 The effects of DEX may be related to its ability to block transcriptional activation by NF-κB,37as well as minimising the cytotoxic effects of NO. Our results with the NF-κB inhibitor, PDTC, further suggest that this transcription factor may play an important role in human iNOS induction in intestinal cells.
Interestingly, Vecchini et al found constitutively expressed iNOS mRNA and NOS activity in proliferating Caco-2 cells, which diminished with differentiation; however, they did not examine cytokine treatment in their studies.38 NOS activity has also been documented in untreated SW 480 and SW 620 cells, which are both human colonic adenocarcinoma cell lines.39HT-29 cells have been shown to express iNOS in response to multiple cytokines, but not when treated with IFN-γ alone.40However, our data clearly show that preconfluent Caco-2BBecells do not express a measurable iNOS mRNA level without stimulation, and even with stimulation the induction is minimal. Furthermore, our experiments in T84 and HT-29 cells suggest that iNOS mRNA induction in response to IFN-γ in Caco-2BBe cells does not merely relate to postconfluent growth, but rather appears to be specifically linked to the differentiation process. Thus, the present data suggest that the ability to produce NO following stimulation by IFN-γ is a feature of differentiated villus enterocytes that distinguishes them from crypt cells. This acquired IFN-γ responsiveness in Caco-2BBe cells correlates with an increase in IFN-γ receptor expression; however, it is likely that other factors may also play a role. For example, T84 cells express very high levels of iNOS in response to IFN-γ and yet express relatively low levels of receptor mRNA, suggesting that other factors may influence the cellular response to IFN-γ.
Intestinal mucosal hyperpermeability has been observed in victims of trauma, thermal injury, and sepsis, and it has been hypothesised that alterations in gut barrier function contribute to the development of multisystem organ failure in these critically ill patients.2 As excessive NO production may be an important factor contributing to gut barrier dysfunction, it will be of interest to define further the mechanisms which govern iNOS activation in the gut, perhaps leading to therapeutic initiatives designed to block gut NO production in critically ill patients.
Acknowledgments
This work was supported by grants from National Institute of Health: DK 50623 (RAH), DK 47186 (RAH), GM 37631 (MPF), and GM 5255–01 (MPF). A M Chavez and M J Morin were supported in part by NIH Training Grant T32 GM07806–17. N Unno was supported in part by the Uehara Memorial Foundation (Japan). We thank Dr M Neutra (Children’s Hospital, Harvard Medical School, Boston, Massachusetts, USA) for providing the Caco-2BBe cell line.
Abbreviations
- CHX
- cycloheximide
- DEX
- dexamethasone
- IFN-γ
- interferon γ
- iNOS
- inducible nitric oxide synthase
- LPS
- lipopolysaccharide
- NF-κB
- nuclear factor κB
- NO
- nitric oxide
- NO2−
- nitrite
- NO3−
- nitrate
- NOS
- nitric oxide synthase
- PDTC
- pyrrolidine dithiocarbamate
- STAT
- signal transducers and activators of transcription
- TNF-α
- tumour necrosis factor α