Article Text

Abstract
Background: Although the contractile, migratory, and proliferative responses of subepithelial myofibroblasts to injury have been postulated to be important events in intestinal wound healing, contractile force generation and migration by these cells has not been investigated previously, and the signals that regulate proliferation by these cells are poorly understood.
Aims: The primary aim of this study was to test the hypothesis that the inflammatory mediator endothelin-1 modulates contraction, migration, and proliferation of intestinal myofibroblasts. We also sought to examine the signal transduction pathways which might underlie these putative effects.
Methods: Contraction, migration, proliferation, cytosolic [Ca2+], and myosin phosphorylation were measured in human colonic subepithelial myofibroblasts in the absence and presence of endothelin receptor agonists and antagonists.
Results: Endothelin-1, but not interleukin 1α, interleukin 6, interleukin 8, interleukin 10, or tumour necrosis factor α, induced a rapid and robust generation of contractile force, which was associated with an increase in cytosolic [Ca2+] and myosin phosphorylation. Inhibition of rho associated kinase reduced endothelin-1 stimulated myosin phosphorylation and contractile force development. Endothelin-1 stimulated migration with a dose-response relationship similar to that observed for contraction. Endothelin A and B receptors mediated contraction while migration was mediated predominantly through endothelin B receptors. Platelet derived growth factor and serum, but not endothelin-1, induced proliferation.
Conclusions: Endothelin-1 stimulates colonic subepithelial myofibroblast contraction and migration via endothelin receptor mediated myosin phosphorylation. These results support an important role for subepithelial myofibroblasts in the injury response of the gut and consequently intestinal wound repair.
- intestinal wound healing
- subepithelial myofibroblasts
- contraction
- migration
- endothelin
- IL, interleukin
- TNF, tumour necrosis factor
- PDGF, platelet derived growth factor
- HEPES, 4-(2-hydroxyethyl)-1-piperazine-ethanesulphonic acid
- [Ca2+]i, cytosolic [Ca2+]
Statistics from Altmetric.com
- IL, interleukin
- TNF, tumour necrosis factor
- PDGF, platelet derived growth factor
- HEPES, 4-(2-hydroxyethyl)-1-piperazine-ethanesulphonic acid
- [Ca2+]i, cytosolic [Ca2+]
Intestinal subepithelial myofibroblasts (which will be referred to as intestinal myofibroblasts) have been postulated to play an important role in the injury response of the small and large intestine.1,2 These mesenchymally derived cells are situated within the lamina propria just beneath the epithelium, adjacent to lymphocytes and macrophages, making them ideally situated to influence epithelial repair. Similar to the better characterised roles of fibroblasts and stellate cells in cutaneous and hepatic wound healing, respectively,1,3 intestinal myofibroblasts are believed to facilitate epithelial restitution. Indirect evidence indicates that injury stimulated contraction of intestinal myofibroblasts is responsible for the rapid diminution of wound surface area observed immediately following epithelial damage in the intestine.4 Subsequently, myofibroblasts are thought to proliferate and migrate to the site of injury permitting scar formation, tissue remodelling, as well as production of growth factors which direct epithelial cell proliferation and cytotaxis.2 Although regulation of intestinal myofibroblast proliferation by growth factors (for example, insulin like growth factor and platelet derived growth factor) has been a topic of recent study,5–7 neither contraction nor migration by intestinal myofibroblasts has been directly investigated.
A role for endothelin-1 in the pathophysiology of intestinal wound repair has been suggested by the localised increase in mucosal endothelin-1 and endothelin receptor levels observed during Crohn's disease and ulcerative colitis.8,9 This peptide, which acts through G protein coupled endothelin A and B receptors, has been demonstrated to stimulate contraction in smooth muscle and myofibroblast like cells,10,11 and migration in lymphocytes, smooth muscle, and endothelial cells.12–14 Moreover, endothelin-1 has been shown to modulate proliferation of numerous myofibroblast like cell types, including Swiss 3T3 fibroblasts, renal mesangial cells, and hepatic stellate cells.15–18 Although the role of endothelin-1 in intestinal myofibroblast contraction, migration, and proliferation has not been explored, the presence of endothelin receptors in this cell type has been reported.19,20
Based on these observations, we hypothesised that the inflammatory mediator endothelin-1 modulates contraction, migration, and proliferation of intestinal myofibroblasts. The primary aim of this study was to test this hypothesis by directly quantitating the effects of endothelin-1 on contraction, migration, and proliferation in colonic subepithelial myofibroblasts (which will be referred to as colonic myofibroblasts). We also sought to examine the signal transduction pathways which might underlie these putative effects.
METHODS
Materials
Interleukin 1α (IL-1α), interleukin 6 (IL-6), interleukin 8 (IL-8), interleukin 10 (IL-10), tumour necrosis factor α (TNF-α), platelet derived growth factor-BB (PDGF-BB), endothelin-1, sarafotoxin, BQ-123, and BQ-788 were obtained from Calbiochem (LaJolla, California, USA) and salts, buffers, and other chemicals from Sigma (St Louis, Missouri, USA). Y-27632 was a gift from the Welfide Corporation (Osaka, Japan).
Colonic myofibroblast model system
Passaged human colonic subepithelial myofibroblasts shown to exhibit most known characteristics of colonic myofibroblasts in situ were employed in this study.7,20 These colonic myofibroblasts (No Co18) were obtained at passage five from the American Type Culture Collection (Rockville, Maryland, USA), having been isolated from a 2.5 month old girl. The cells were cultured on Primaria treated dishes (Falcon; Becton Dickinson, Franklin Lakes, New Jersey, USA) in NEAA Earle's salts containing 10% fetal bovine serum, 2% 𝓁-glutamine, 1% sodium pyruvate, and 1% penicillin and streptomycin in a 37°C humidified incubator with 10% CO2. Stock cultures of these cells were split 1:3 or 1:4 every three days to prevent them from becoming confluent. These colonic myofibroblasts were discarded at passage 15, after which they have been reported to acquire an altered phenotype.20
Measurement of contractile force generation
The contractile force generated by colonic myofibroblasts was directly quantitated using a method previously used to investigate contraction by other types of non-muscle cells.11,21 Briefly, 1×106 cells were cultured within a solid but elastic collagen gel. After 2–3 days, collagen gels containing colonic myofibroblasts were attached vertically between an isometric force transducer and a fixed rod, and placed in an organ bath containing serum free media with 25 mM 4-(2-hydroxyethyl)-1-piperazineethanesulphonic acid (HEPES) at 37°C. The gels containing colonic myofibroblasts were equilibrated for one hour to ensure a stable baseline tension. Agonists and antagonists were added directly to the bath, and changes in contractile force were recorded digitally.
Measurement of cytosolic [Ca2+]
Colonic myofibroblasts (4×104/0.1 ml collagen) were grown within a solid but elastic collagen gel (2×10 mm) attached horizontally across a No 1 glass coverslip. After 2–3 days, this preparation was transferred to an electrolyte solution (135 mM NaCl, 5 mM KCl, 0.8 mM MgCl, 1.2 mM CaCl2, 0.8 mM NaH2PO4, 10 mM HEPES, and 5 mM glucose) containing 5 μM of the acetoxy-methyl ester form of the calcium sensitive fluorescent dye, fluo3 (Molecular Probes, Eugene, Oregon, USA).22 After one hour at 24°C, to permit loading and de-esterification of the fluo3, the preparation was washed twice with electrolyte solution and placed into an imaging chamber heated to 37°C which was mounted to an epifluorescence microscope. Using a 40× 0.75 numerical aperture objective, epifluorescent images of cells loaded with fluo3 were collected at a light wavelength of 535 nm after excitation at a wavelength of 488 nm. Images were collected with a cooled CCD camera every 30 seconds. Image analysis software (Metamorph; Universal Imaging, Westchester, Pennsylvania, USA) was used to measure changes in fluorescent intensity induced by endothelin-1. Change in cytosolic [Ca2+] ([Ca2+]i) was determined as the change in fluorescence intensity divided by basal fluorescence intensity (ΔF/F0) within the cytosol.22 Transmitted light images were examined before and after treatment with endothelin-1, and no changes in cell morphology were observed.
Measurement of myosin phosphorylation
Colonic myofibroblasts were grown to confluence and placed in serum free media for 60 minutes at 37°C prior to each experiment. Agonists and antagonists were added directly to the culture dishes as indicated. Myosin phosphorylation was determined as described previously using glycerol/acrylamide gel electrophoresis and immunoblot with an antibody directed against the myosin II regulatory light chain.23–25 Myosin phosphorylation is presented as the sum of mono- and diphosphorylated myosin II regulatory light chain as a percentage of total myosin II regulatory light chain, as determined by densitometry.
Measurement of migration
Wound induced migration was measured as previously described with some modifications.26 Colonic myofibroblasts were cultured on 22 mm glass coverslips (2.0×105 cells/coverslip). After forming a confluent monolayer, cells were placed in serum free media for 16 hours prior to creation of linear “wounds” through the cell monolayer using a sterile needle. Following wounding, cells were placed in serum free media in the presence or absence of agonists and/or inhibitors. To quantitate the rate of migration, phase contrast images were collected using a 10× 0.5 numerical aperture objective. Initial wound quantitation was performed on images collected 20 minutes after wounding because wound size was unstable prior to this. Further images were collected randomly from at least five different wounded areas 24 hours after wounding. Wound edges were identified, and the width and area of the cell free region measured using imaging software (Metamorph; Universal Imaging Corporation). Wound induced migration was calculated as the reduction in wound area as a percentage of wound area at initial quantitation (that is, maximal wound area).
Measurement of proliferation
Colonic myofibroblast proliferation was investigated using the CyQUANT proliferation kit (Molecular Probes). Cells plated onto 96 well plates at a density of 600 cells/well were exposed to agonist and/or antagonists for two days and then stained with a well characterised fluorescent nucleic acid binding dye.27,28 The method was validated by staining serial dilutions of colonic myofibroblasts. Fluorescence emission plotted against cell number created a calibration curve with r2≈1.0.
Statistical methods
Data are presented as mean (SEM) and were evaluated for statistical significance using the Student's t test. Statistical significance was defined as p<0.05.
RESULTS
Endothelin-1 stimulated contractile force generation
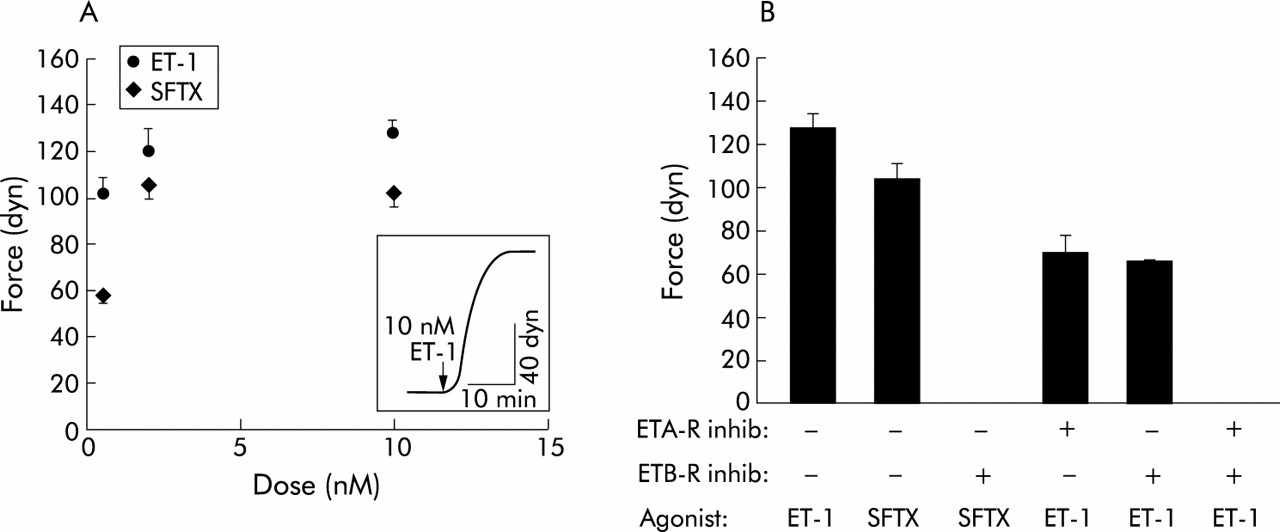
To test the hypothesis that endothelin-1 stimulates colonic myofibroblast contraction, cells were treated with a range of endothelin-1 doses (0.5–10 nM) or carrier (0 nM endothelin-1). Endothelin-1 stimulated colonic myofibroblast contraction in a dose dependent manner (fig 1A). In all experiments, cells achieved a preponderance of contractile tension within five minutes of stimulation, and a plateau within 10–15 minutes. Maximum contraction of 128 (6) dyn (n=7) occurred in response to 10 nM endothelin-1.

Contractile force generation by colonic myofibroblasts. (A) Endothelin-1 (ET-1) and sarafotoxin (SFTX) dose-response curves. Each data point represents the mean (SEM) (n>3 for each point). The ET-1 and SFTX dose curves are significantly different (p<0.05). (Inset) Representative contraction stimulated by ET-1. (B) Contractile force generation in response to 10 nM SFTX or 10 nM ET-1 was measured in the presence or absence of 10 μM BQ-123 (ETA-R inhib) and/or 10 μM BQ-788 (ETB-R inhib), as indicated. Each bar represents the mean (SEM) (n=7 for ET-1 and SFTX without inhibitor and n=3 for all other experiments). ET-1 stimulated contraction was significantly (p<0.05) greater than SFTX stimulated contraction. BQ-123 and BQ-788 each significantly (p<0.05) inhibited ET-1 stimulated contraction.
Endothelin-1 stimulated contractile force generation was mediated by both endothelin A and endothelin B receptors
To examine whether endothelin-1 stimulated contractile force generation was mediated by endothelin B receptors, a selective endothelin B receptor agonist, sarafotoxin, was tested for its ability to induce colonic myofibroblast contraction. Sarafotoxin stimulated contraction of colonic myofibroblasts in a dose dependent manner (fig 1A) which was significantly less than that induced by endothelin-1. A maximum contraction of 103 (7) dyn (n=7) was achieved at a concentration of 10 nM sarafotoxin. To investigate the specificity of sarafotoxin as an endothelin B receptor agonist, we exposed colonic myofibroblasts to a selective endothelin B receptor antagonist, BQ-788, prior to sarafotoxin stimulation. BQ-788 completely blocked sarafotoxin stimulated contractile force generation.
To determine the role of endothelin A and endothelin B receptors in endothelin-1 stimulated colonic myofibroblast contraction, a selective endothelin A receptor antagonist, BQ-123, and BQ-788 were used (fig 1B). Incubation with BQ-123 significantly inhibited endothelin-1 stimulated contraction by ∼45% while BQ-788 significantly inhibited endothelin-1 stimulated contraction by ∼48%. Incubation with a combination of BQ-123 and BQ-788 completely abolished endothelin-1 stimulated contractile force generation.
Other inflammatory mediators did not affect contraction
To examine the effects of other peptides, which are released in response to injury, on colonic myofibroblast contraction, force generation was characterised in response to 100 ng/ml of IL-1α, IL-6, IL-8, IL-10, or TNF-α following a 60 minute incubation in serum free media. None of these cytokines altered contractile force generation during 15 minutes of exposure (data not shown). Subsequently, the same cells were stimulated with 10 nM endothelin-1, which resulted in a level of contractile force generation no different from that of cells treated with endothelin-1 in the absence of cytokines.
Endothelin-1 induced increases in [Ca2+]i and myosin phosphorylation
To explore the mechanisms by which endothelin-1 stimulates colonic myofibroblast force generation, [Ca2+]i and phosphorylation of myosin were examined in response to endothelin-1 stimulation (fig 2). Exposure to 10 nM endothelin-1 stimulated an increase in [Ca2+]i, which was biphasic (fig 2A). There was an initial relatively large increase in [Ca2+]i followed by a sustained [Ca2+]i at a level between peak and basal levels.

Endothelin-1 (ET-1) induced increases in [Ca2+]i and myosin phosphorylation in colonic myofibroblasts. (A) Time course of [Ca2+]i in the presence of 10 nM ET-1. The change in [Ca2+]i is presented as the change in fluo3 fluorescence intensity divided by the fluo fluorescence intensity at baseline (ΔF/F0). Each data point represents the mean (SEM) (n=3 for each point). (B) Time course of myosin phosphorylation stimulated by 10 nM ET-1 at time 0. Myosin phosphorylation is presented as the sum of monophosphorylated and diphosphorylated forms of myosin II regulatory light chain as a percentage of total myosin II regulatory light chain. Each data point represents the mean (SEM) (n=8 for each point).
Endothelin-1 10 nM stimulated a greater than threefold increase in myosin phosphorylation by colonic myofibroblasts, from a basal level of 25 (16)% phoshorylation to a maximum of 90 (7)% at three minutes (n=8). Maximal phosphorylation of myosin was observed by two minutes and remained stable for the duration of the experiments (15 minutes).
Y-27632 inhibited myosin phosphorylation and contractile force generation
To further characterise the mechanisms by which endothelin-1 stimulated contractile force generation, we used Y-27632, a selective inhibitor of rho associated kinase, which is an important downstream target of the small GTPase, rhoA.29 Y-27632 treatment resulted in a dose dependent decrease in endothelin-1 stimulated myosin phosphorylation (fig 3A). Y-27632 also caused dose dependent inhibition of endothelin-1 induced contractile force generation (fig 3B).

Effects of Y-27632 on endothelin-1 (ET-1) stimulated myosin phosphorylation and contractile force generation by colonic myofibroblasts. (A) Myosin phosphorylation in response to 10 nM ET-1 (ET-1 +) or carrier (−) in the presence of different concentrations of the rho kinase inhibitor Y-27632 (rho K inhib in μM) for 30 minutes. Myosin phosphorylation is presented as the sum of monophosphorylated and diphosphorylated forms of myosin II regulatory light chain as a percentage of total myosin II regulatory light chain. Each bar represents the mean (SEM) (n=3 for each point). (B) Y-27632 (rho K inhib in μM) dose-response curve for 10 nM ET-1 stimulated contractile force generation after 30 minutes of exposure to Y-27632. In this figure, contractile force is presented as a percentage of control (that is, force generated by 10 nM ET-1 and no Y-27632). Each bar represents the mean (SEM) (n=3 for each point).
Endothelin-1 stimulated migration
The role of endothelin-1 in the regulation of wound induced migration by colonic myofibroblasts was investigated. Endothelin-1 stimulated a dose dependent increase in migration (fig 4A). The rate of wound induced migration was ∼4 μ/min and ∼7 μ/min for untreated and 10 nM endothelin-1 treated colonic myofibroblasts, respectively. The receptors mediating the effects of endothelin-1 were examined using selective endothelin receptor agonists and antagonists. The endothelin B receptor agonist, sarafotoxin, stimulated migration almost to the same extent as endothelin-1 (fig 4B). This endothelin B receptor predominance was confirmed by the observation that the endothelin B receptor antagonist BQ-788 strongly inhibited the effects of endothelin-1 on migration while the endothelin A receptor antagonist BQ-123 had no observed effect.

Effect of endothelin-1 (ET-1) on migration by colonic myofibroblasts. (A) ET-1 stimulated migration in a dose dependent manner. Migration is presented as the reduction in wound area at 24 hours as a percentage of the maximal wound area (that is, initial quantitation). Each data point represents the mean (SEM) (n=5 for each point). (B) Migration in response to 10 nM sarafotoxin (SFTX) or 10 nM ET-1 was measured at 24 hours in the presence or absence of 10 μM BQ-123 (ETA-R inhib) or 10 μM BQ-788 (ETB-R inhib), as indicated. Each bar represents the mean (SEM) (n=5). ET-1 stimulated migration was significantly (p<0.05) inhibited by the endothelin B receptor antagonist but not by the endothelin A antagonist.
Endothelin-1 did not modulate proliferation
To investigate the effects of endothelin-1 on colonic myofibroblast proliferation, colonic myofibroblasts were grown in the presence and absence of 10 nM endothelin-1 for two days, and the relative number of colonic myofibroblasts were quantified (fig 5). Endothelin-1 did not stimulate colonic myofibroblast proliferation or inhibit fetal bovine serum stimulated colonic myofibroblast proliferation. In contrast, both 10% fetal bovine serum and 100 ng/ml PDGF-BB significantly stimulated proliferation of colonic myofibroblasts.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Endothelin-1 (ET-1) did not modulate proliferation by colonic myofibroblasts. Colonic myofibroblasts were treated with ET-1, platelet derived growth factor-BB (PDGF-BB), fetal bovine serum (FBS), ET-1 and FBS, or carrier (control) at the concentrations indicated. Proliferation was determined after two days. FBS and PDGF-BB significantly (p<0.05) enhanced proliferation compared with controls or ET-1. Each bar represents the mean (SEM) (n=6 trials with 12 samples/trial).
DISCUSSION
It has been proposed that myofibroblasts play an important role in the response of the intestine to injury. Circumstantial evidence has suggested that early during injury, contraction of intestinal myofibroblasts rapidly reduces mucosal defects and decreases the area requiring re-epithelialisation.4 Later in the injury response, it has been believed that intestinal myofibroblasts are stimulated to migrate into the wound, proliferate, and mediate wound healing by secreting growth factors, which direct epithelial cell proliferation and chemotaxis, and extracellular matrix proteins and matrix metalloproteases, which permit tissue remodelling.2,6,30–33 In this study, we have ascertained for the first time that colonic myofibroblasts are capable of contractile force generation and wound induced migration. Moreover, we have demonstrated that endothelin-1, one of a cascade of inflammatory mediators released during epithelial injury,8,9 stimulates contraction and migration by colonic myofibroblasts with a time course that is consistent with in vivo observations.4 Together, these data provide novel evidence supporting a role for colonic myofibroblasts and endothelin-1 in intestinal wound healing.
In this study, among a number of substances (that is, IL-1α, IL-6, IL-8, IL-10, and TNF-α) that are produced during intestinal injury,34,35 only endothelin-1 stimulated colonic myofibroblast contraction. Endothelin-1, a small peptide which acts through G protein coupled endothelin A and B receptors, is best recognised as a potent agonist for smooth muscle contraction.36 In smooth muscle it has long been known that contractile force is powered by the myosin II motor protein complex (myosin), which is activated by phosphorylation of its regulatory light chain by Ca2+ calmodulin stimulation of the myosin light chain kinase.37 More recently it has been demonstrated that the monomeric GTPase, rhoA, acting through its downstream effector, rho associated kinase, also stimulates myosin phosphorylation and consequently contractile force generation.38 However, the signals that mediate stimulus-contraction coupling in non-muscle cell types, such as intestinal myofibroblasts, are poorly understood.
To investigate whether endothelin A receptors, endothelin B receptors, or both mediate endothelin-1 stimulated contractile force generation, we used sarafotoxin, an endothelin B receptor agonist, BQ-123, an endothelin A receptor antagonist, and BQ-788, an endothelin B receptor antagonist. Experiments with sarafotoxin suggested that endothelin-1 stimulated contraction is mediated mostly, but not entirely, through endothelin B receptors. This finding is consistent with prior observations that alterations in intestinal myofibroblast morphology, a surrogate for contraction, are induced principally by endothelin B receptors.19 However, experiments using BQ-123 and BQ-788 indicated that both endothelin A and B receptor subtypes play a quantitatively important role in colonic myofibroblast contraction. From these data it cannot be concluded as to whether or not endothelin B receptors play a predominant role in the regulation of contraction. We believe that the apparent differences between the sarafotoxin and endothelin receptor inhibitor data result from limitations in the specificity of these agonists and antagonists. Therefore, further experiments, which are beyond the scope of the current study, will be necessary to resolve the relative contributions of each endothelin receptor to endothelin-1 stimulated contraction. Regardless, these results strongly indicate that endothelin A and B receptors both play a role in mediating colonic myofibroblast contractile force generation.
With regard to contractile signals which lie further downstream, we observed that endothelin-1 stimulated a rapid and transient elevation in [Ca2+]i in colonic myofibroblasts, similar to that previously seen in duodenal myofibroblasts.19 In analogy to smooth muscle cells, this endothelin-1 induced increase in [Ca2+]i suggested that colonic myofibroblast contractile force generation might also be powered by activation of myosin. Therefore, we examined the effects of endothelin-1 on phosphorylation of myosin. We determined that endothelin-1 strongly induced myosin phosphorylation by colonic myofibroblasts, which supports a role for myosin in contractile force generation in this cell type. We tested this notion by employing a highly selective inhibitor of rho associated kinase, Y-27632.39 First we confirmed that Y-27632 was a potent inhibitor of endothelin-1 stimulated myosin phosphorylation in colonic myofibroblasts. We then examined the effects of blocking myosin phosphorylation with Y-27632, and found that Y-27632 strongly inhibited colonic myofibroblast contractile force generation. Moreover, these findings suggest that rhoA signalling pathways are also involved in the regulation of contractile force generation in colonic myofibroblasts. Together these data indicate that endothelin-1 stimulated contractile force generation in colonic myofibroblasts is powered by myosin, and that this may be regulated by both calcium and rhoA signalling pathways.
Cellular migration is a complex process involving dynamic changes in the actin-myosin cytoskeleton, including generation of contractile forces,40 which suggested that migration of colonic myofibroblasts might also be influenced by endothelin-1. The results of this study demonstrated that endothelin-1 stimulates wound induced migration of colonic myofibroblasts. The concentrations of endothelin-1 which altered migration were similar to concentrations which stimulated myosin phosphorylation and contractile force development. These data are consistent with the notion that the signalling pathways that mediate colonic myofibroblast contraction may also contribute to migration.
Similar to the study that demonstrated that the shape of intestinal myofibroblasts is controlled principally by endothelin B receptors,19 endothelin-1 induced colonic myofibroblast migration appeared to be predominantly mediated by endothelin B receptors. Inhibition of endothelin A receptors did not affect endothelin-1 stimulated migration while endothelin B receptor blockade substantially inhibited endothelin-1 stimulated migration. This was confirmed by the finding that the endothelin B receptor agonist, sarafotoxin, stimulated migration to almost the same magnitude as endothelin-1.
Our observations indicate that endothelin-1 had no effect on colonic myofibroblast proliferation. This is significant because endothelin-1 influences proliferation in many cell types. For example, endothelin-1 stimulates proliferation in a number of cell types, including Swiss 3T3 fibroblasts, renal mesangial cells, and aortic smooth muscle,15,16 while it inhibits proliferation in other cell types, such as activated hepatic stellate cells.17,18 Although proliferation of colonic myofibroblasts was not influenced by endothelin-1, we confirmed that another mediator of the injury response cascade, PDGF, stimulated colonic myofibroblast proliferation.6
A limitation of this study is that colonic myofibroblasts in cell culture may not behave identically to colonic myofibroblasts in situ. To maximise the clinical relevance of this study, we used a previously characterised cell model that has been demonstrated to exhibit many features of colonic myofibroblasts in situ, including a stellate morphology, expression of α smooth muscle actin, and the presence of functional endothelin-1 receptors.7,20 Moreover, the pertinence of the findings of this study are supported by the prior observations that endothelin receptor antagonists substantially altered colonic wound healing in rats8 and that the time course of wound retraction in guinea pig ileum was similar to the time course of contractile force generation measured in this study.4 Despite the potential differences between the responses of colonic myofibroblasts in situ versus in culture, this study provides the initial evidence that colonic myofibroblasts are capable of contracting and migrating.
In summary, we have demonstrated in colonic myofibroblasts that endothelin-1 stimulated contractile force generation and migration, via endothelin receptor induced myosin phosphorylation, but did not affect proliferation. These results provide novel and important evidence supporting the following model of intestinal wound repair. During epithelial injury, numerous inflammatory mediators are produced. One of those mediators, endothelin-1, rapidly stimulates subepithelial myofibroblast contraction, which results in a swift reduction in the size of the mucosal defect. Endothelin-1 also stimulates migration of these cells into the wound where other inflammatory mediators, including PDGF, induce proliferation. This augmented population of myofibroblasts mediates wound healing by secreting growth factors which direct epithelial cell proliferation and chemotaxis, and extracellular matrix proteins and matrix metalloproteases, which permit tissue remodelling.
Acknowledgments
We thank Dr Maria Abreu Martin for helpful discussions, Drs Richard Aranda and the late John Walsh for their thoughtful critiques of this manuscript, Dr Suraj Bhat for use of his epifluorescence microplate reader, Matthew Thimgan for technical assistance, and the Welfide Corporation for Y-27632. Supported by awards to HY from the National Institutes of Health (K08 DK02450) and the Glaxo-Wellcome Institute for Digestive Health, and an American Digestive Health Foundation/American Gastroenterological Association Student Research Fellowship Award to LK.
REFERENCES
Footnotes
-
L E Kernochan and B N Tran contributed equally to this work.