


Abstract
Previous studies demonstrated that analogs of benztropine [3α-(diphenyl-methoxy)tropane (BZT)] bind to the dopamine (DA) transporter with high affinity, inhibit DA uptake, but do not maintain rates of responding in self-administration procedures comparable with those maintained by cocaine. Some BZT analogs have an onset of action that is slower than that for cocaine that may contribute to this decreased effectiveness. In addition, some BZT analogs have affinity for muscarinic-M1 receptors that may interfere with reinforcing effects. The present study assessed effects of BZT analogs in place-conditioning procedures designed to accommodate variations in onset of effect. BZT analogs with variations in relative affinities for the DA transporter over M1 receptors from equal [AHN 1-055 (3α-[bis(4′-fluorophenyl)methoxy]-tropane)] to 16-fold [JHW 007 (N-(n-butyl)-3α-[bis(4′-fluorophenyl)methoxy]-tropane)] were compared with cocaine and the muscarinic antagonist, atropine. Cocaine (10–20 mg/kg) but not atropine (1.0–5.6 mg/kg) produced dose-related place conditioning. The N-methyl-substituted BZT analog, AHN 1-055, was without significant effects at doses that ranged from 0.3 to 3.0 mg/kg and when administered up to 90 min before conditioning trials. In contrast, effects of AHN 2-005 (N-allyl-3α-[bis(4′-fluorophenyl)methoxy]-tropane; 0.1–10.0 mg/kg) were significant, and those of JHW 007 approached significance when administered 45 min but not immediately or 90 min before trials. Atropine blocked the effect of AHN 2-005 and approached significant antagonism of cocaine. The present study further supports and extends previous results showing minimal preclinical indications of abuse liability of BZT analogs and suggests that these differences from cocaine are not entirely accounted for by a slower onset of action or muscarinic M1 receptor affinity.
Benztropine [BZT; 3α-(diphenylmethoxy)tropane] is the parent compound in a class of dopamine (DA) transport inhibitors with structural similarity to cocaine (i.e., a tropane ring), as well as to the 1,4-dialyky(en)ylpiperazines (i.e., a diphenyl-ether moiety), exemplified by GBR 12909. A series of BZT analogs showed enhanced affinities for binding to the DA transporter relative to BZT and cocaine (Katz et al., 1999). Despite this increased affinity, the behavioral effects of these compounds were generally different from those of cocaine and other DA uptake inhibitors. For example, 4-chloro-3α-(diphenylmethoxy)tropane (4-Cl-BZT) has a 30 nM affinity for the DA transporter, which is comparable with the high-affinity binding of cocaine. However, 4-Cl-BZT has reduced effectiveness as a stimulant of locomotor activity compared with cocaine and does not produce cocaine-like discriminative stimulus effects (Katz et al., 1999; Tolliver et al., 1999). Furthermore, this compound does not maintain rates of responding as high as those maintained by cocaine in a self-administration procedure (Woolverton et al., 2000) or break points as high as those for cocaine under a progressive ratio schedule (Woolverton et al., 2001). The differences between the BZT analogs compared with cocaine may appear at odds with the DA transporter hypothesis of the behavioral effects of cocaine, which suggests that compounds that bind to the DA transporter and inhibit DA uptake will have behavioral effects like those of cocaine (Kuhar et al., 1991).
Some of the BZT analogs have a time course that is different from that of cocaine. For example, Tolliver et al. (1999) and Tanda et al. (2003) showed that increases in extracellular DA produced by cocaine occurred sooner after injection than those produced by the 4-Cl analog of BZT. Those results with 4-Cl-BZT are similar to results for BZT reported previously (Church et al., 1987). A pharmacokinetic comparison of cocaine and a series of BZT analogs indicated that all of these compounds have high permeability and are detectable in brain minutes after injection (Raje et al., 2003), suggesting that differences in onset of action are not fully attributable to differences in central nervous system permeability between cocaine and the BZT analogs. Regardless of the source, a delay in the onset of action of BZT analogs could account for the differences between cocaine and BZT analogs in studies of their reinforcing effects. Because self-administration procedures involve the direct reinforcement of a response with the injection of drug, a small delay in onset of effect could act like a delay in reinforcement, and decrease the rate of responding maintained by the drug (e.g., Gollub and Yanagita, 1974; Stretch et al., 1976; Beardsley and Balster, 1993; see also Lile et al., 2003).
Delays in onset of action may be accommodated in place-conditioning procedures by modifying the time between injection and placement of the subject in the conditioning chamber. For example, De Beun et al. (1992) examined place conditioning induced by gonadotropin-releasing hormone at various times before the subject was placed in the conditioning chamber. Injection of the hormone effectively produced place conditioning when administered between 15 and 45 min before placement of the subject in the chamber. In contrast, the hormone did not produce place conditioning when administered either immediately or at 75 to 120 min before subjects were placed in the chamber. The authors noted that the time course of effect was consistent with the time course of other behavioral effects of the peptide. Thus, the assessment of effects of a drug with delayed onset of effect under place-conditioning procedures may involve selection of an appropriate time between injection and conditioning session and need not compromise the conditioning of drug and environment (see also Fudala and Iwamoto, 1990; Cunningham et al., 2002).
There have also been suggestions that the reinforcing effects of BZT analogs may be limited by their antagonist actions at muscarinic M1 receptors. For example, Ranaldi and Woolverton (2002) found that combinations of the anticholinergic, scopolamine, and cocaine were generally less effective in maintaining high rates of responding than was cocaine alone. A decrease in rates of responding maintained by cocaine due to coadministration of a muscarinic antagonist is consistent with previous results indicating a lesser effectiveness of BZT analogs compared with cocaine in self-administration procedures (Woolverton et al., 2000, 2001). Therefore, the present study examined a series of BZT analogs in which the affinity for the muscarinic M1 receptor was varied with changes in chemical structure, and those effects were compared with the effects of atropine, using a place-conditioning procedure.
Materials and Methods
Subjects. Sprague-Dawley rats (Taconic Farms, Germantown, NY), weighing about 180 to 200 g upon arrival in the laboratory, served. The subjects were housed in a temperature- and humidity-controlled colony room with a 12-h light/dark cycle. All experiments were conducted during the light phase of the light/dark cycle. Subjects were allowed to habituate to the colony room for at least 1 week prior to their use in these experiments, and each subject was routinely handled three times before the start of the experiment. Food and water were available at all times except during experimental sessions. Experiments were conducted in accordance with National Institutes of Health Guidelines under Institutional Animal Care and Use Committee-approved protocols.
Apparatus. Place conditioning was conducted in 40- × 40-cm acrylic test chambers that were comprised of two equally sized (20 × 40 cm) separate compartments (AccuScan Instruments, Inc., Columbus, OH). The two compartments were constructed of either clear or black acrylic and separated by inserting a clear acrylic 10- × 6-cm guillotine door. The floor of the clear compartment was constructed of stainless steel mesh, under which there was β-chip bedding. The floor of the black compartment was a constructed of stainless steel rods under which there was no bedding. Each chamber was placed within a monitor that determined the location of the subject by light-sensitive detectors spaced 2.5 cm apart along two perpendicular walls and infrared light sources directed at the detectors on the opposing walls. Time spent in each compartment and locomotor activity (total distance traveled) was recorded and generated by the VersaMax system (Accuscan Instruments, Inc.). The chambers and monitors were housed within a dimly illuminated room.
Procedures. Each experiment was carried out for 13 consecutive days and was divided into three phases (Li et al., 2002). During the preconditioning phase, subjects were placed in the chamber close to the middle of the apparatus in the clear side and allowed access to both compartments for 15 min each day on 4 consecutive days. The time spent in each compartment was recorded on the 4th day. If the time for an individual subject in either compartment exceeded 10 min, it was removed from the study to select a subject pool without inherent predispositions for occupancy of one or the other compartment. During the present studies, about 25% of the subjects were removed based on the results of this initial preconditioning phase.
The conditioning phase lasted for eight daily sessions. Drug or saline injections were administered on alternate days, and subjects were placed in one of the two compartments for 30 min with the door separating the compartments in place. The drug treatments that alternated with vehicle injections in different groups of subjects were: cocaine, atropine, AHN 1-055, AHN 2-005, JHW 007, cocaine and atropine, and AHN 2-005 and atropine. Unless otherwise indicated, all injections were administered immediately before subjects were placed in the chamber. The effects of each treatment on locomotor activity were assessed during its first administration by tabulating the number of photocell beams broken during the 30-min exposure period.
For studies using the 45- and 90-min pretreatment intervals, subjects were injected and placed back in their home cages until 45 or 90 min later, when they were placed into the test chamber. Returning the subject to the home cage ensured that the appropriate compartment was only paired with drug during a designated 30-min period and precluded the pairing of the designated side of the chamber with drug effects at a different time. For the study of the effects of cocaine with atropine, the atropine was injected 10 min before and cocaine was injected immediately before the subject was placed into the test chamber. For the study of the effects of AHN 2-005 with atropine, the AHN 2-005 was given, and the subject was placed back in its home cage until 35 min later, when it was given the atropine injection and placed 10 min later into the test chamber.
The postconditioning session was conducted 24 h after the last conditioning session. During this session, as in the preconditioning phase, subjects were placed in the chamber close to the middle of the apparatus in the clear side and allowed access to both compartments for 15 min, and the time spent in each compartment was recorded.
For studies of the effects of the BZT analogs, cocaine (10 mg/kg) and saline groups were run concurrently as positive and negative controls, respectively. Whenever possible, these controls were shared between tests of different drugs to minimize animal usage. Because these studies were conducted in different shipments of subjects, these positive and negative controls were repeated in each different shipment. Shared control groups were used for tests of AHN 1-055 and JHW 007, and a second group of shared controls was used for tests of cocaine and AHN 2-005.
Drugs. The drugs studied were cocaine hydrochloride (Sigma-Aldrich, St. Louis, MO), atropine sulfate (Sigma-Aldrich), and several N-substituted analogs of BZT (Fig. 1): AHN 1-055, the 4′,4″-difluorophenyl analog of BZT; AHN 2-005; and JHW 007. The synthesis of these analogs was conducted in the Medicinal Chemistry Section of the National Institute on Drug Abuse Intramural Research Program and has been described previously (Agoston et al., 1997). All drugs were dissolved in saline (cocaine and atropine) or distilled water (AHN 1-055, AHN 2-005, and JHW 007), with heat and sonication, as necessary. The drug solutions were administered i.p. at 1 ml/kg, with doses administered on the basis of body weight.
Statistical Analysis. For the place-conditioning test, the time spent in the drug-paired compartment during the postconditioning session was expressed as a difference from that during the last preconditioning session (CPP score). The locomotor activity was expressed as the total horizontal distance traveled (centimeters) during the first conditioning sessions (30 min). The difference in group means were analyzed by one-way analysis of variance (ANOVA) followed by Dunnett's multiple comparisons of treatments versus saline controls. When single doses of cocaine were tested along with one of the benztropine analogs as a positive control, its effects were compared with vehicle controls by unpaired Student's t tests for independent samples. In studies of combinations of cocaine or AHN 2-005 and atropine, ANOVA and Dunnett's tests employed either saline or cocaine effects as the comparison. Effects with calculated p < 0.05 were considered statistically significant.
Results
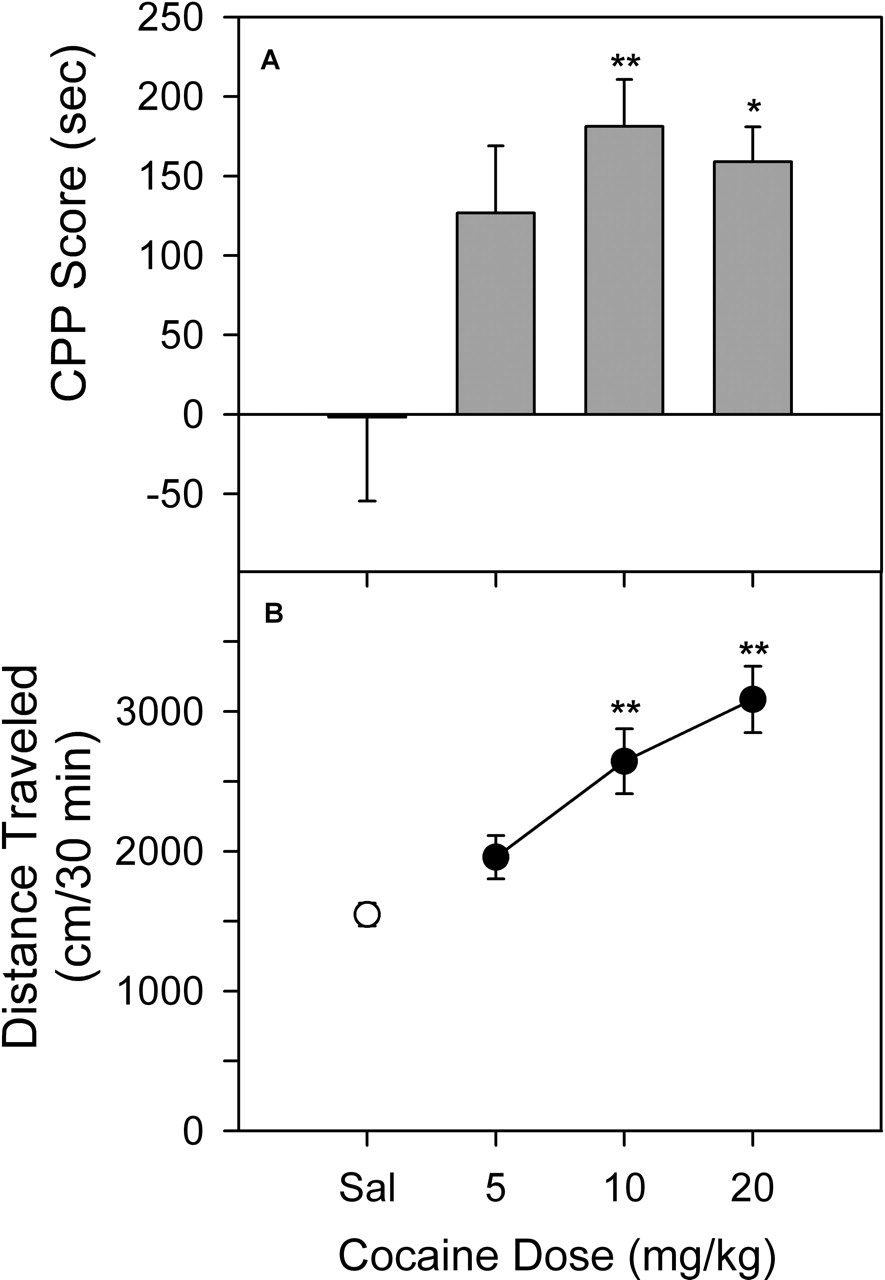
Cocaine-Induced Place Conditioning and Activity. At no point during the study did subjects receiving saline treatment prior to placement in both compartments exhibit a significant change in the time spent in one of the compartments after conditioning (the mean value for the change in time from all of the saline conditions in the study was -1.7 ± 52.8 s). In contrast, cocaine produced a significant (F3,28 = 4.53, p < 0.05) dose-related increase in time spent in the drug-paired compartment (Fig. 2A). The effects of 5 mg/kg cocaine were not significant compared with the saline group; however, a significant increase in time was obtained at doses of 10 and 20 mg/kg, with the maximum response obtained at 10 mg/kg cocaine (Dunnett's q = 3.36, p < 0.01). Results obtained in a separate group of subjects showed that cocaine also increased locomotor activity during the first conditioning session (Fig. 2B). The increases were directly related to doses from 5 to 20 mg/kg (F3,68 = 13.4, p < 0.001), and activity after 10 and 20 mg/kg was significantly different from that for the saline group (Dunnett's q = 4.13 and 5.80, p < 0.01).
Atropine-Induced Place Conditioning and Activity. Atropine was studied at doses that had previously been shown to potentiate the effects of cocaine (Katz et al., 1999). At these doses, atropine did not significantly affect (F3,28 = 2.21, p = 0.109) time spent in the drug-paired compartment when compared with the saline group (Fig. 3A). At the 3.0-mg/kg dose, there was an indication of a decrease in time spent on the drug-paired side that was not significant (Dunnett's q = 2.21; p > 0.05). The cocaine-conditioning control group, as in the previous study, showed significant increase in time spent on the drug-paired compartment (t14 = 2.53, p < 0.05). Atropine, at doses from 1.0 to 5.6 mg/kg (Fig. 3B), did not significantly alter locomotor activity during the first conditioning session (F3,28 = 0.0327, p = 0.992). In contrast, the group given 10.0 mg/kg cocaine showed a significant stimulation of locomotor activity compared with the saline group (t14 = 4.30, p < 0.001).

Basic structures of the presently studied BZT analogs with substitutions on the nitrogen.
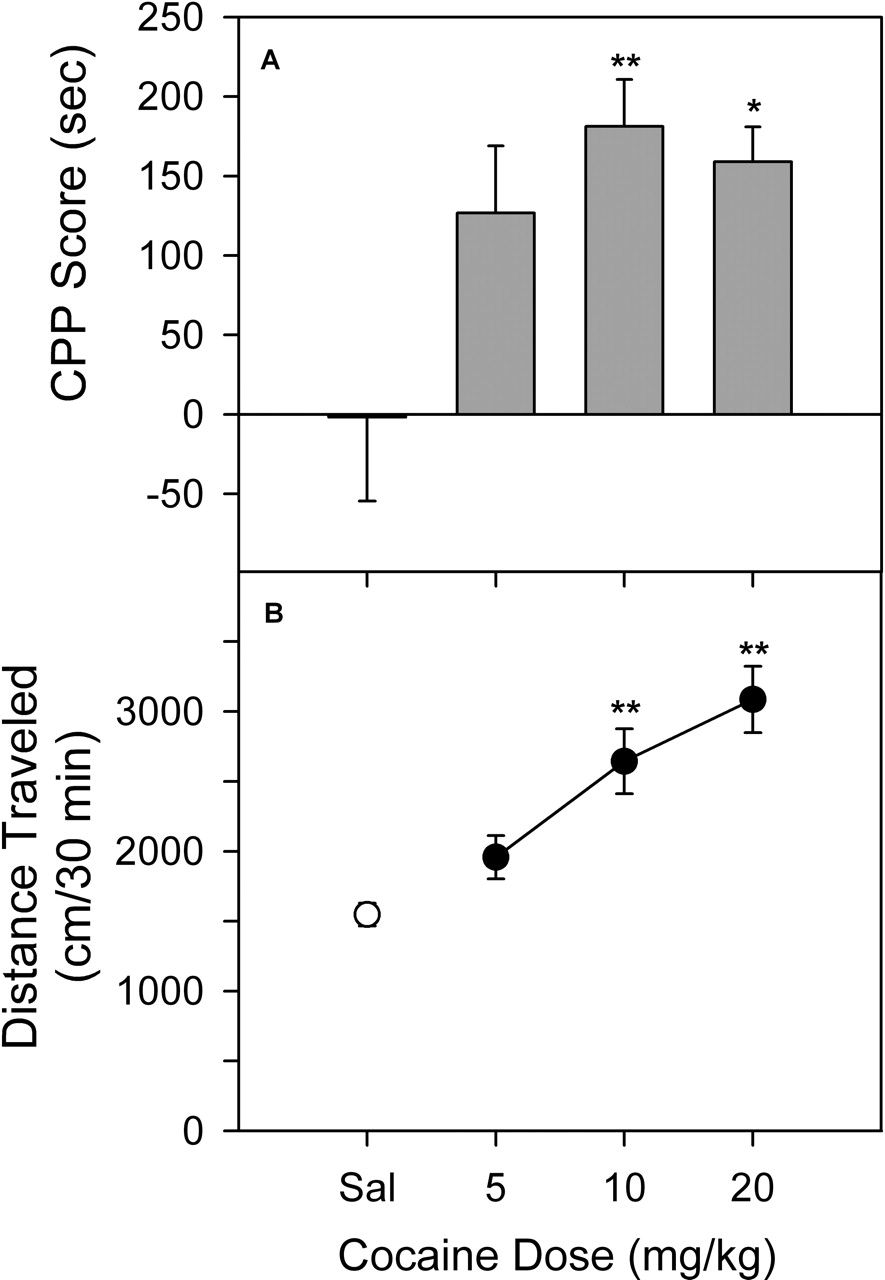
Place-conditioning (A) and locomotor (B) effects of cocaine in rats. The CPP scores represent the time spent in the drug-paired side during the postconditioning expressed as a difference from that during the last preconditioning session. The locomotor activity was expressed as the total horizontal distance traveled (centimeters) during the first exposure to drug in the first conditioning session. Data are presented as means ± S.E.M. *, p < 0.05; **, p < 0.01, as determined by the Dunnett's test versus saline group.
AHN 1-055-Induced Place Conditioning and Activity. Treatment with the various doses of AHN 1-055 immediately before the conditioning sessions did not produce a significant change (F3,28 = 0.704, p = 0.558) in the time spent in either compartment, whereas 10 mg/kg cocaine produced significant (t14 = 2.16, p < 0.05) place conditioning compared with that of saline (Fig. 4A). Because increases in time between injection and testing increased the efficacy of AHN 1-055 in substituting for cocaine (Katz et al., 1999), the place-conditioning tests were also conducted with injections administered at 45 and 90 min before conditioning sessions. Neither the 45-min (Fig. 4B; F3,28 = 0.624, p = 0.606) nor the 90-min (Fig. 4C; F3,28 = 0.698, p = 0.561) pretreatment time increased the time spent in the drug-paired compartment across the range of doses examined.

Place conditioning (A) and locomotor effects (B) of atropine in rats. All details are as described in Fig. 2.
There was no significant locomotor stimulating effects of AHN 1-055 when it was injected immediately before conditioning (Fig. 4D, diamonds), whereas 10 mg/kg cocaine, as before, significantly increased locomotor activity (t14 = 2.37, p < 0.05). Although there was a trend for an increase in activity when AHN 1-055 was administered immediately before testing, the increase was not significant. Similarly, there was no effect on activity when the drug was administered 45 min before the session (Fig. 4D, triangles). In contrast to these results, when AHN 1-055 was administered 90 min before the first conditioning session, there was a significant dose-related decrease in locomotor activity (F3,28 = 4.12, p < 0.05; Fig. 4D, squares).
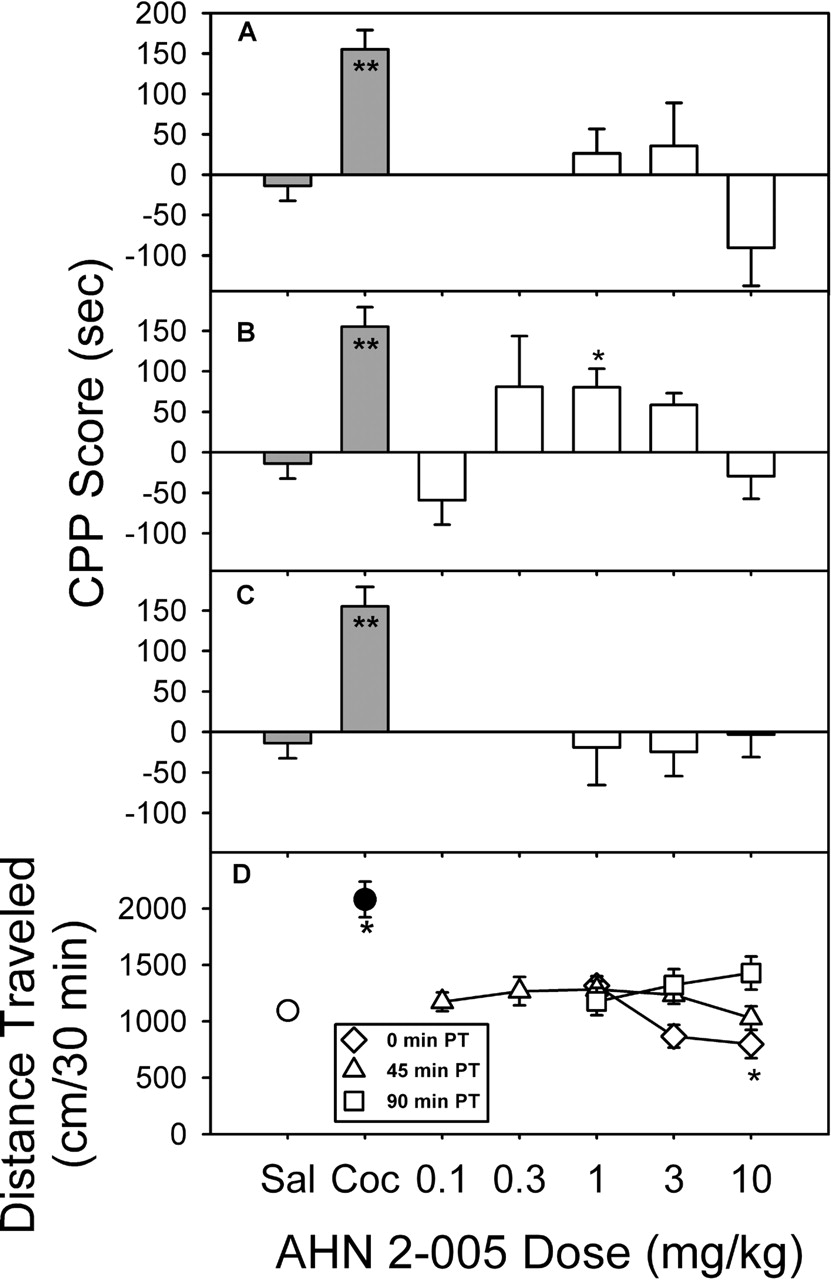
AHN 2-005-Induced Place Conditioning and Activity. With conditioning sessions conducted immediately after injection (Fig. 5A), there was no significant change (F3,43 = 2.28, p = 0.0934) in the amount of time spent on either side of the chamber. In contrast, when AHN 2-005 was administered 45 min before conditioning sessions (Fig. 5B), there was an increase in the amount of time spent in the drug-paired compartment (F5,82 = 4.59, p = 0.001), with 1.0 mg/kg showing significant effects (Dunnett's q = 2.92, p < 0.05). With conditioning sessions conducted 90 min after injection (Fig. 5C), there was no significant change (F3,44 = 0.0745, p = 0.973) in the amount of time spent on either side of the chamber.

Place conditioning (A–C) and locomotor effects (D) of AHN 1-055 administered to rats at different times before placement in the conditioning chamber. A to C, place conditioning effects of AHN 1-055 given 0, 45, and 90 min before conditioning sessions, respectively. Data are presented as means ± S.E.M. Significant differences from control (p < 0.05) are indicated by *, as determined by Student's t test versus saline group for the cocaine groups and by ANOVA followed by the Dunnett's test versus saline group for AHN 1-055 groups.
AHN 2-005 injected immediately before the first conditioning session (Fig. 5D, diamonds) produced a dose-related decrease in locomotor activity (F3,43 = 5.12, p < 0.01), whereas 10 mg/kg cocaine produced a significant stimulation of locomotor activity compared with the saline group (t46 = 5.78, p < 0.0001). In contrast, when administered either 45 or 90 min before the conditioning sessions, there were no significant effects of AHN 2-005 on locomotor activity (F5,81 = 1.49, p = 0.203 and F3,44 = 2.25, p = 0.0953; Fig. 5D, triangles and squares, respectively).

Place conditioning (A–C) and locomotor effects (D) of AHN 2-005 administered to rats at different times before placement in the conditioning chambers. All details are as described in Fig. 4.
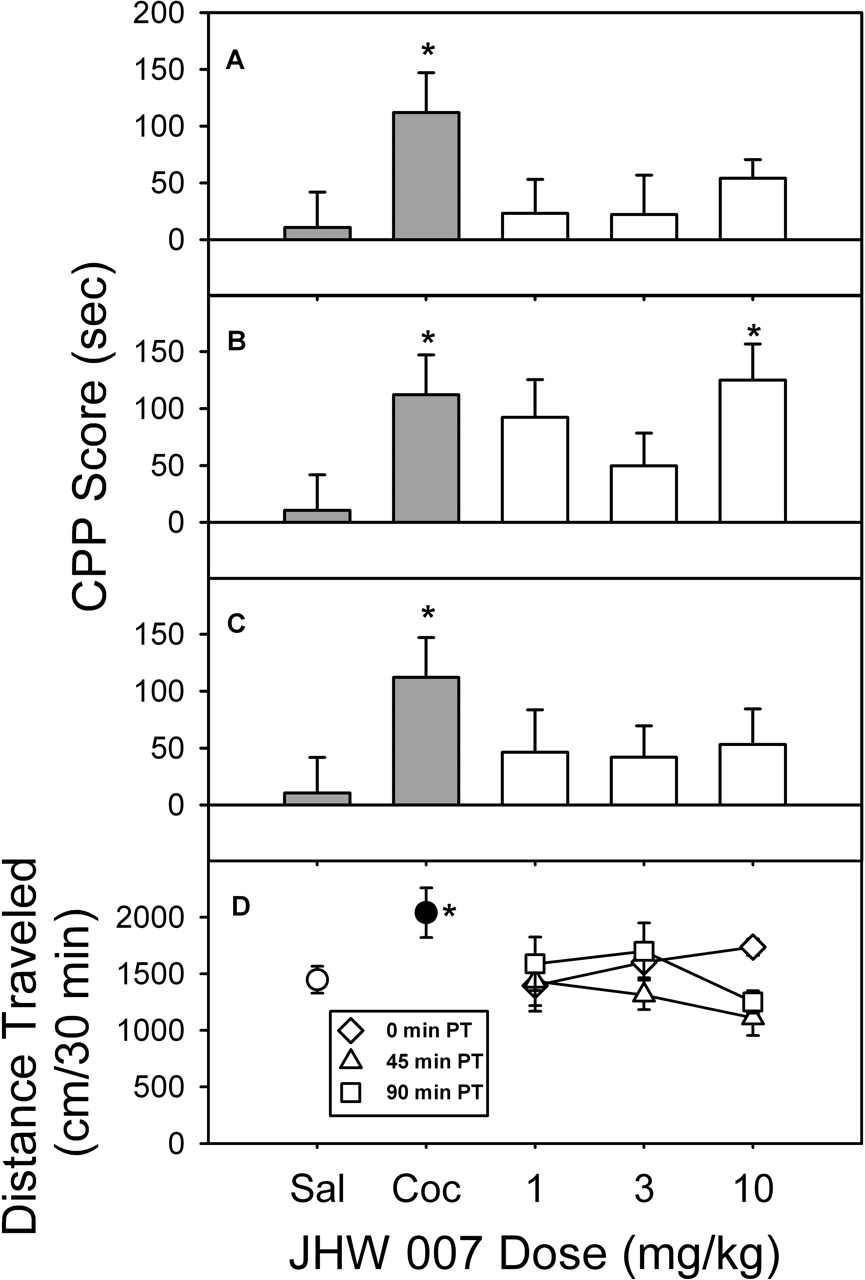
JHW 007-Induced Place Conditioning and Activity. As with the other BZT analogs, when JHW 007 was administered immediately before conditioning sessions (Fig. 6A), there was no significant change in the amount of time spent on either side of the chamber (F3,28 = 0.415, p = 0.744). With the 45-min pretreatment (Fig. 6B), the overall ANOVA approached significance (F3,28 = 2.56, p = 0.0754) and a Dunnett's test indicated that the time spent in the drug-paired compartment at the 10-mg/kg dose was significantly different from vehicle (Dunnett's q = 2.59, p < 0.05). When JHW 007 was administered 90 min before conditioning sessions (Fig. 6C), there was no significant effect on the distribution of time in the two compartments across the entire dose range studied (F3,28 = 0.349, p = 0.790).
JHW 007 did not produce a significant change (F3,28 = 1.07, p = 0.378) in locomotor activity when it was injected immediately before the first conditioning session (Fig. 6D, diamonds). At the highest dose administered 45 min after injection (Fig. 6D, triangles), there was a decrease in activity that was not significant (F3,28 = 0.951, p = 0.430). Similarly, there was no significant change in locomotor activity when JHW 007 was injected 90 min before conditioning (F3,28 = 1.05, p = 0.387; Fig. 6D, squares).

Time course of the place conditioning (A–C) and locomotor effects (D) of JHW 007 in rats. All details are as described in Fig. 4.
Effects of Atropine on Cocaine-Induced Place Conditioning and Activity. There were no significant treatment effects revealed by an ANOVA of the effects of saline alone and atropine-cocaine combinations (Fig. 7A; F3,27 = 2.41, p = 0.0891). As in previous tests, the place-conditioning effects of cocaine alone were significantly different from saline alone (t14 = 5.51, p < 0.0001). The effects of cocaine with 1.0 to 5.6 mg/kg atropine were not significantly different from those of cocaine alone treatment (F3,27 = 2.33, p = 0.0963).
During the first conditioning session, cocaine significantly enhanced locomotor activity (t14 = 2.25, p < 0.05) compared with the saline group (Fig. 7B). There were no significant differences between effects of saline alone and combinations of atropine and cocaine on locomotor stimulation (F3,27 = 2.15, p = 0.118), and there was no significant effect of atropine on cocaine-induced locomotor stimulation (F3,27 = 1.02, p = 0.399).
Effects of Atropine on AHN 2-005-Induced Place Conditioning and Activity. As in the previous study of AHN 2-005 administered 45 min before place conditioning (Fig. 5B), there was a significant effect of 1.0 mg/kg AHN 2-005 administered alone (t14 = 2.21; p < 0.05; Fig. 8A). This effect of AHN 2-005 was attenuated by combined administration of atropine (F3,28 = 5.64, p < 0.01). In addition, atropine (3.0 and 5.6 mg/kg) decreased time spent on the drug-paired side of the chamber produced by AHN 2-005 compared with AHN 2-005 alone (Dunnett's q = 3.93 and 2.60, p < 0.01 and 0.05, respectively). There was a trend toward a conditioned place avoidance produced by combinations of atropine and AHN 2-005; however, that effect was not significant (F3,28 = 2.75, p = 0.0616). There was a small but significant attenuation of locomotor activity in the first session (Fig. 8B) produced by 1.0 mg/kg AHN 2-005 administered 45 min before testing compared with the saline group (t14 = 4.07, p < 0.01) and no attenuation of this effect by atropine (F3,28 = 0.451, p = 0.718).

The effects of atropine on cocaine-induced place conditioning (A) and locomotor (B) effects. Atropine and cocaine were given 10 and 0 min before conditioning. All details are as described in Fig. 2.
Discussion
Analogs of BZT bind to the dopamine transporter and inhibit DA uptake but differ from cocaine with regard to their reinforcing effects that are predictive of abuse liability. It is possible that a slower onset of action or an affinity for muscarinic M1 receptors may contribute to differences between BZT analogs and cocaine. Therefore, the present study compared effects of several BZT analogs that had differences in affinity for M1 receptors relative to their affinity for the DA transporter. In addition, we examined the place-conditioning effects of these drugs that, like reinforcing effects, are predictive of abuse liability, with a procedural modification to accommodate different onsets of action.
As has been shown previously (Carr et al., 1989; Tzschentke, 1998), cocaine showed significant place conditioning, with maximal effects at 10 mg/kg. In contrast, atropine did not show any significant effects, although there was a trend toward an increase in the time spent in the saline-paired compartment. AHN 1-055 did not produce place-conditioning effects across the range of doses up to 3.0 mg/kg, despite effects on locomotor activity. Similarly, neither AHN 2-005 nor JHW 007 produced significant place-conditioning effects when administered immediately or 90 min before conditioning sessions, although each of these drugs produced a significant effect when administered 45 min before conditioning sessions.
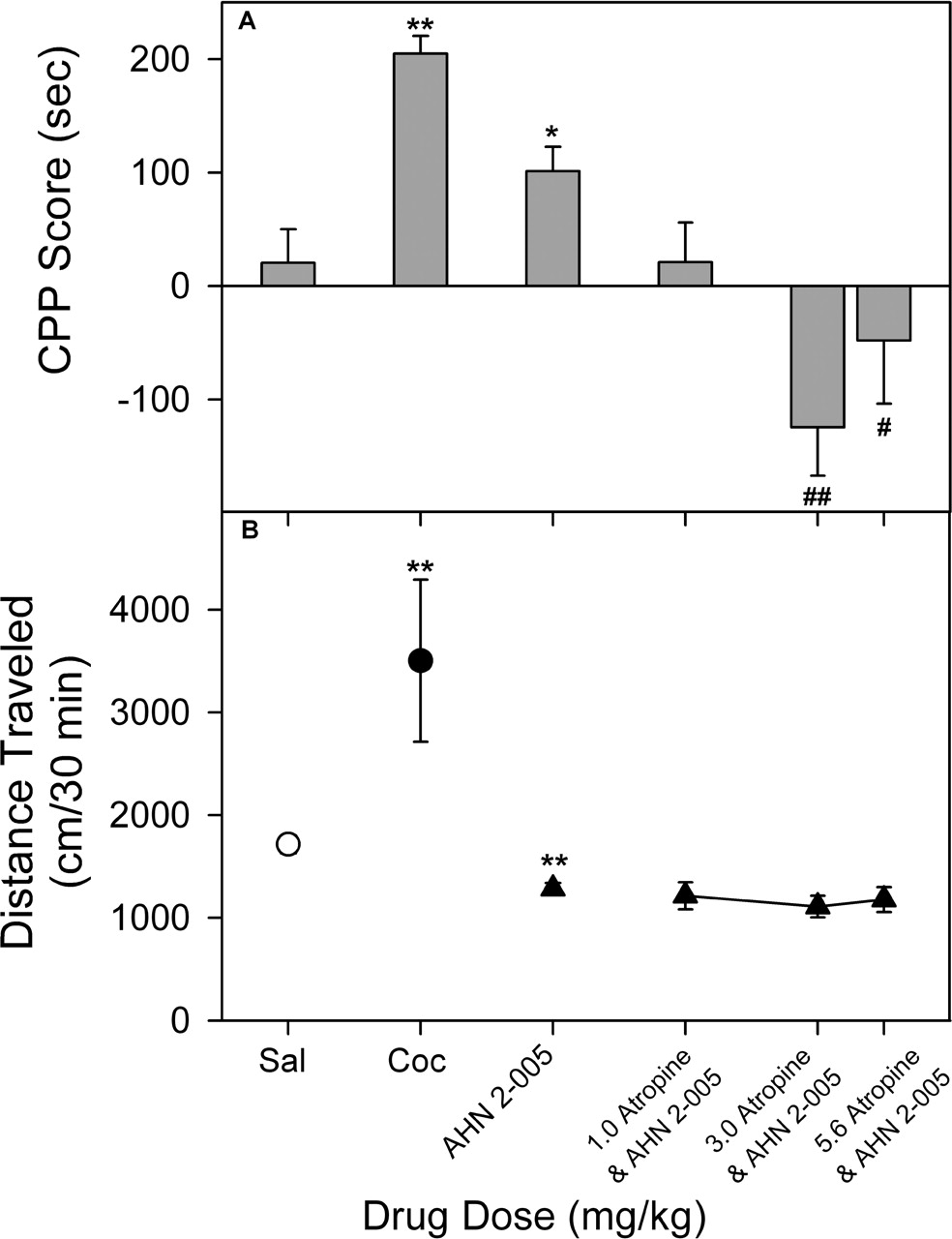
The effects of atropine on AHN 2-005-induced place conditioning (A) and locomotor effects (B). Atropine and cocaine were given 10 and 45 min before conditioning. All details are as described in Fig. 1 except that: #, p < 0.05; ##, p < 0.01 determined by ANOVA followed by the Dunnett's test versus AHN 2-005 group.
The lack of consistent place conditioning with the BZT analogs contrasts with results obtained with other DA uptake inhibitors, such as in addition to cocaine, mazindol, methylphenidate, nomifensine, and bupropion (Schechter and Calcagnetti, 1993; Tzschentke, 1998). As mentioned above, the BZT analogs in general are less effective than cocaine in stimulating locomotor activity, in producing cocaine-like discriminative stimulus effects (Katz et al., 1999, 2004), and in maintaining high rates of responding in self-administration procedures (Woolverton et al., 2000, 2001). These diminished behavioral effects are obtained despite increased affinity of the drugs for the DA transporter relative to cocaine (Katz et al., 1999, 2004). The decreased effectiveness of some BZT analogs in light of increased DA transporter affinity suggests differences in the mechanisms of action between BZT analogs and DA transporter ligands with substantial preclinical indications of abuse liability.
Previous in vivo microdialysis studies indicated differences in the onset of action between selected BZT analogs and cocaine, with cocaine showing a more rapid increase in extracellular DA levels after injection (Church et al., 1987; Tolliver et al., 1999; Tanda et al., 2003). As previously mentioned, delays in onset of effect can substantially diminish the effectiveness of a drug reinforcer (e.g., Balster and Schuster, 1973; Stretch et al., 1976; Beardsley and Balster, 1993; Panlilio et al., 1998). However, with delays between treatment and placement in the chamber place-conditioning procedures can be sensitive to effects of drugs with delayed onsets of action (De Beun et al., 1992).
Each of the BZT analogs presently studied had greater affinity for the DA transporter than for the NE and 5-HT transporters as measured with radioligand binding assays (see Katz et al., 2004). In contrast to the other compounds, AHN 1-055 had a relatively high affinity for the muscarinic M1 receptor, which was comparable with its affinity for the DA transporter. In contrast, AHN 2-005 and JHW 007 had, respectively, 6- and 16-fold higher affinity for the DA transporter than for M1 receptors. The greater selectivity of these compounds for the DA transporter may contribute to their greater effects in producing a place conditioning at 45 min after injection.
Recently Ranaldi and Woolverton (2002) found that combinations of the anticholinergic, scopolamine, and cocaine were generally less effective in maintaining self-administration than was cocaine alone. From this finding, the authors suggested that the antimuscarinic effects of BZT analogs limit their effectiveness in self-administration procedures (cf. Woolverton et al., 2000, 2001). These authors further suggested that the decreased effectiveness of scopolamine-cocaine combinations, compared with cocaine alone, could be due to a pharmacological antagonism of the reinforcing effects of cocaine by scopolamine or due to a behavioral interaction of reinforcement by cocaine injection and punishment by the response-dependent administration of scopolamine.
There are several studies that relate to these interactions, and potentially relate to the present effects of BZT analogs. Wilson and Schuster (1973) found that atropine increased rather than decreased rates of responding maintained by cocaine. Increases in response rates are inconsistent with a punishment of responding. The dose of cocaine used by Wilson and Schuster was on the descending limb of the dose-effect curve (Wilson et al., 1971), indicating that the atropine-induced increases were similar to the effect of lowering cocaine dose. Furthermore, because the atropine was administered independently of responding before experimental sessions, it could not function as a punishing stimulus. Thus, Wilson and Schuster's results suggest a pharmacological noncompetitive antagonism of the reinforcing effects of cocaine by the anticholinergic. Other findings suggest the interaction is pharmacological. For example, the anticholinergics, scopolamine and trihexyphenidyl, have both been reported to be inactive in place-conditioning procedures (Lynch, 1991; Shimosato et al., 2001). In addition, there is at least one paper reporting self-administration with scopolamine (Glick and Guido, 1982). Thus, there is no published evidence supporting punishing effects of anticholinergic drugs.
Trends that failed to reach significance in the present study moderate firm conclusions regarding the pharmacological or behavioral basis for the interaction. For example, atropine when administered alone produced a nonsignificant trend toward conditioned place avoidance, suggesting a behavioral rather than pharmacological interaction. Furthermore, the trends suggesting place avoidance with combinations of atropine and AHN 2-005 and decreases in effectiveness of cocaine when administered with atropine both suggest a behavioral basis to the interaction. Thus, selected aspects of the present results suggest the potential of a anticholinergic punishing effect; however, those effects are not significant, and the weight of the published literature currently suggests a physiological antagonism of the reinforcing effects of DA uptake inhibitors by anticholinergics.
The alternative hypotheses discussed above relate specifically to drug self-administration and place conditioning procedures. However, there is a wider range of behavioral differences between the BZT analogs and cocaine-like DA uptake inhibitors. For example, BZT analogs are generally less effective than cocaine-like DA uptake inhibitors in stimulating locomotor activity (Katz et al., 1999, 2004) and in producing effects on conditioned behavior (Acri et al., 1996). Biochemical studies of novel BZT analogs are providing evidence that these drugs act at the DA transporter in a manner that differs from that of other DA transporter ligands, such as cocaine (Vaughan et al., 1999; Reith et al., 2001; Chen et al., 2004). Those findings suggest that fundamental differences between the mode in which the BZT analogs and cocaine-like compounds bind to the DA transporter protein may in part account for the differences in their behavioral effects.
Studies of novel BZT analogs may further discovery research on leads for the development of pharmacotherapies for cocaine abuse (Newman and Kulkarni, 2002). In this latter context, positive place conditioning results might suggest some undesirable abuse liability of potential treatment drugs. However, consideration of the fact that the place conditioning was obtained only with a relatively long delay after injection and at only one time point suggests that their direct reinforcing effects would be muted by a delay of reinforcement. In addition, some minimal reinforcing effect could potentially facilitate compliance, if these compounds were ever used as treatments. Methadone and buprenorphine, which are currently used as treatments for opioid abuse, each show place-conditioning effects (Steinpreis et al., 1996; Tzschentke, 2004) and are self-administered (Mello and Mendelson, 1985).
In conclusion, the results of the present study further support previous studies showing a decreased effectiveness of BZT analogs compared with cocaine and extend those findings to an additional animal model predictive of abuse liability. The weight of the evidence at this time suggests that the decreased effectiveness in animal models of abuse liability is not due to a punishment-like effect conferred on the compounds through antimuscarinic effects. Alternative mechanistic explanations for lower predicted abuse liability may involve antagonism of cocaine-like effects by the antimuscarinic effects of the compounds; however, this explanation is restricted to the effects related to abuse liability and does not necessarily apply to other behavioral effects of the BZT analogs (Katz et al., 1999) or to all BZT analogs, in particular, those with reduced affinity for muscarinic receptors (Newman and Kulkarni, 2002). Previous suggestions that BZT analogs act at the DA transporter in a manner that is fundamentally different from that for cocaine may relate more generally to all of the behavioral effects of novel BZT analogs.
Acknowledgments
We thank Jianjing Cao for expert technical assistance in resynthesizing AHN 1-055, AHN 2-005, and JHW 007; Paul Nuzzo, Rajeev I. Desai, Leigh V. Panlilio, Charles W. Schindler, and Philip Terry for advice on the manuscript; and Patty Ballerstadt for administrative assistance.
Footnotes
-
Article, publication date, and citation information can be found at http://jpet.aspetjournals.org.
-
doi:10.1124/jpet.105.084541.
-
ABBREVIATIONS: BZT, benztropine; DA, dopamine; GBR 12909, 1-{2-[bis-(4-fluorophenyl)methoxy]ethyl}-4-(3-phenylpropyl)piperazine; 4-Cl-BZT, 4-chloro-3α-(diphenylmethoxy)tropane; AHN 1-055, 3α-[bis(4′-fluorophenyl)methoxy]-tropane; AHN 2-005, N-allyl-3α-[bis(4′-fluorophenyl)-methoxy]-tropane; JHW 007, N-(n-butyl)-3α-[bis(4′-fluorophenyl)methoxy]-tropane; ANOVA, analysis of variance; CPP, conditioned place preference.
- Received February 3, 2005.
- Accepted February 28, 2005.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}