


Abstract
In rat caudal artery, contraction to melatonin results primarily from activation of MT1 melatonin receptors; however, the role of MT2 melatonin receptors in vascular responses is controversial. We examined and compared the expression and function of MT2 receptors with that of MT1 receptors in male rat caudal artery. MT1 and MT2 melatonin receptor mRNA was amplified by reverse transcription-polymerase chain reaction from caudal arteries of three rat strains (i.e., Fisher, Sprague-Dawley, and Wistar). Antisense (but not sense)33P-labeled oligonucleotide probes specific for MT1 or MT2 receptor mRNA hybridized to smooth muscle, as well as intimal and adventitial layers, of caudal artery. In male Fisher rat caudal artery denuded of endothelium, melatonin was 10 times more potent than 6-chloromelatonin to potentiate contraction to phenylephrine, suggesting activation of smooth muscle MT1melatonin receptors. The MT1/MT2 competitive melatonin receptor antagonist luzindole (3 μM), blocked melatonin-mediated contraction (0.1–100 nM) with an affinity constant (KB value of 157 nM) similar to that for the human MT1 receptor. However, at melatonin concentrations above 100 nM, luzindole potentiated the contractile response, suggesting blockade of MT2 receptors mediating vasorelaxation and/or an inverse agonist effect at MT1constitutively active receptors. The involvement of MT2receptors in vasorelaxation is supported by the finding that the competitive antagonists 4-phenyl 2-acetamidotetraline and 4-phenyl-2-propionamidotetraline, at MT2-selective concentrations (10 nM), significantly enhanced contractile responses to all melatonin concentrations tested (0.1 nM–10 μM). We conclude that MT2 melatonin receptors expressed in vascular smooth muscle mediate vasodilation in contrast to vascular MT1 receptors mediating vasoconstriction.
Accumulating evidence indicates that the hormone melatonin regulates vascular tone; however, the nature of the response remains controversial. Both vasoconstrictor and vasodilator responses have been reported; however, data supporting the presence of melatonin receptors is found in some, but not all, vascular beds (Mahle et al., 1997). In isolated rat caudal arteries, nanomolar concentrations of melatonin potentiate contraction induced by either endogenous or exogenous vasoconstrictors (Viswanathan et al., 1990; Evans et al., 1992; Krause et al., 1995; Mahle et al., 1997; Ting et al., 1997; Geary et al., 1998). In rat cerebral arteries and arterioles, melatonin is a direct vasoconstrictor (Geary et al., 1997; Viswanathan et al., 1997). Melatonin-mediated contraction in caudal and cerebral arteries is blocked by melatonin receptor antagonists (Viswanathan et al., 1992; Krause et al., 1995; Geary et al., 1997; Ting et al., 1997, 1999).
Melatonin also dilates rat and rabbit aorta, iliac, renal, and basilar arteries (Satake et al., 1986; Shibata et al., 1989; Weekley, 1991); however, the nature of the receptor(s) involved is not known. The lack of specific 2-[125I]iodomelatonin binding in rat aorta (Viswanathan et al., 1990) suggests either that the melatonin-mediated vasodilation is not mediated by high-affinity melatonin receptors or that the sensitivity of the binding assay was not sufficient for detection of all melatonin receptor types (Dubocovich et al., 1998). In porcine pulmonary, coronary, and marginal colon arteries, high concentrations of melatonin and melatonin analogs may induce vasodilatation by a novel mechanism (Ting et al., 2000).
The MT1 and MT2 melatonin receptors are possible mediators of the physiological effects of melatonin (Dubocovich et al., 2000). They exhibit distinct structural, chromosomal, and pharmacological differences (Reppert et al., 1995,1996; Slaugenhaupt et al., 1995). Available agonists generally show little discrimination between the melatonin receptor types; however, 6-chloromelatonin exhibits lower affinity than melatonin on the MT1 receptor but equal affinity on the MT2 receptor (Dubocovich et al., 1997). Luzindole, an often-used competitive melatonin receptor antagonist, shows 25 times higher affinity for MT2 than MT1 receptors. In contrast, the amidotetraline ligands 4P-ADOT and 4P-PDOT show at least 1000-fold higher affinity for the MT2 compared with the MT1 type and they act as competitive MT2 receptor antagonists in native tissues (Dubocovich et al., 1998; Hunt et al., 2001). Luzindole or the amidotetralines, when used at 0.1 μM or higher concentrations, may also act as MT1 competitive antagonists (Krause et al., 1995; Regrigny et al., 1999; Browning et al., 2000). Furthermore, luzindole and 4P-PDOT are inverse agonists in systems where receptors are constitutively active, including the rat caudal artery (Dubocovich and Masana, 1998; Browning et al., 2000; Masana and Dubocovich, 2001; Ersahin et al., 2002).
It is now well accepted that activation of MT1melatonin receptors in caudal arteries facilitates adrenergic vasoconstriction (Krause et al., 1995; Ting et al., 1997, 1999; Bucher et al., 1999; Lew and Flanders, 1999). However, definitive demonstration of vascular MT2 receptors has been hampered by technical difficulties and discrepancies in technical approaches, rat strains and gender, drug types, and concentrations. Results from our laboratory suggest that the contractile responses to melatonin in male Fischer rat caudal arteries are mediated through activation of at least two distinct receptors (Doolen et al., 1998,1999). The enhanced contractile responses to melatonin in the presence of 4P-ADOT, at a concentration that does not discriminate between MT1 and MT2 melatonin receptors, suggest blockade of a receptor-mediating relaxation (Doolen et al., 1998). A role for MT2 melatonin receptors in mediating vasorelaxation was also suggested in female Fisher rat caudal arteries (Doolen et al., 1999). However, the presence of MT2 melatonin receptors and their role in vasodilation has been challenged by Ting et al. (1999). These authors were unable to demonstrate MT2 melatonin receptor mRNA expression in male Wistar caudal arteries and reported that 4P-PDOT did not affect melatonin's potentiation of electrically induced vasocontraction.
To address these discrepancies, in this study we determined the expression of MT1 and MT2melatonin receptor mRNA using highly sensitive reverse transcription-polymerase chain reaction (RT-PCR) and in situ hybridization techniques in caudal arteries of three rat strains. Furthermore, we reassessed the effects of selective MT2 melatonin receptor antagonists (Dubocovich et al., 1997, 1998) on melatonin-mediated potentiation of arterial contraction. We demonstrate that the MT2melatonin receptor is indeed present in caudal artery and acts to attenuate MT1 melatonin receptor-mediated contraction.
Materials and Methods
Animals.
Four-month-old Fisher-344, Wistar, or Sprague-Dawley male rats, obtained from Harlan (Indianapolis, IN), were maintained in temperature- (22 ± 1°C) and humidity-controlled rooms. Food and water were provided ad libitum. Animals were maintained in a 12-h light/dark cycle for at least 2 weeks before each experiment. Animals were sacrificed during the middle of the light cycle. All animal care and procedures were performed in accordance with institutional guidelines.
RT-PCR.
Fisher-344 (n = 8), Wistar (n = 8), and Sprague-Dawley (n = 8) male rats were sacrificed by decapitation and the caudal arteries immediately isolated and frozen at −80°C. Total RNA was extracted using the TRIzol reagent (Invitrogen, Carlsbad, CA) according to manufacturer's instructions. The RNA pellet was dissolved in diethylpyrocarbonate-treated water and stored at −80°C until use. To eliminate any residual contamination of DNA, duplicate tubes containing 1 μg of total RNA were pretreated with 1 unit of amplification grade DNase I (Invitrogen) in DNase I reaction buffer in the presence of 20 units of RNase inhibitor (PerkinElmer Life Sciences, Boston, MA) at room temperature. After 15 min, 1 μl of 25 mM EDTA solution, pH 8.0, was added to each tube, samples were heated for 10 min at 65°C to inactivate the DNase I, and thereafter immediately cooled on ice. Synthesis of first-strand cDNA and subsequent polymerase chain reaction (PCR) amplification were carried out using the Titan One Tube RT-PCR system (Roche Applied Science, Indianapolis, IN) in a PCR DNA thermocycler (PerkinElmer Life Sciences). DNase-treated RNA samples were reverse transcribed to cDNA in the presence of dNTP (each 2.5 mM) and AMV reverse transcriptase, using gene-specific primers. This procedure was followed by amplification in the same tube using an Expand High Fidelity enzyme blend (Taq polymerase and Pwo DNA polymerase) in the presence of 1.5 mM MgCl2and 1 μM each of sense- and antisense-specific primers. The samples were incubated at 50°C for 30 min followed by thermocycling: one cycle at 94°C for 2 min; 10 cycles at 94°C for 30 s, 50°C for 30 s, and 68°C for 1 min; and 30 cycles at 94°C for 30 s, 50°C for 30 s, and 68°C for 1 min, plus elongation of 5 s for each cycle and a final elongation time of 10 min at 68°C. A reverse transcriptase negative reaction (enzyme inactivated at 94°C for 2 min) was run to determine whether the amplification product came exclusively from the RNA. RNA extraction, DNA reverse transcription, and PCR assays were conducted in a laboratory where melatonin receptor clones were never handled. Forward and reverse primer oligonucleotides were designed from the partial cDNA sequences derived from GenBank and by computer analysis using the PrimerSelect program in the LASERGENE Navigator software (DNAStar, Madison, WI) and synthesized at the Northwestern University Biotechnology Facility (Chicago, IL) (Fig. 1A). For the MT1 receptor (accession no. U14409), one set of primers amplified a 431-base pair (bp) fragment: forward primer 5′-TAGGATATACAGTAACAAGAAT-3′, corresponding to nucleotides 30 to 51, and reverse primer 5′-CTAGCCACGAAGAGCCACT-3′, complementary to nucleotides 460 to 442. Another set of nested primers amplified a 366-bp fragment: forward primer 5′-AGAATTCCCTGTGCTACG-3′, corresponding to nucleotides 47 to 64, and reverse primer 5′-CTGAGGCCACAATAAGACC-3′, complementary to nucleotides 412 to 394. For the MT2 receptor (accession no. U28218) one set of primers amplified a 310-bp fragment: forward primer 5′-TGCGACCTACCACCGAGCC-3′, corresponding to nucleotides 21 to 39, and reverse primer 5′-T(T/C)A(A/G)(G/A)GG(G/A)GCCCAGC AAA(C/T)(T/G)GCAA-3′, a degenerate primer that can amplify MT1 or MT2 sequences corresponding to nucleotides 814 to 791 of the human receptor sequence (accession no. U25341). The set of nested primers amplified a 264-bp fragment: forward primer 5′-CCCTCTACATCAGCCTCA-3′, corresponding to nucleotides 59 to 77, and reverse primer 5′-CACTGGGTCTCAGGCGTA-3′, complementary to nucleotides 334 to 315 of the rat MT2 melatonin receptor.

RT-PCR amplification of MT1 and MT2 melatonin receptors from rat caudal artery. A, alignment of the nucleotide sequences for the rat melatonin receptors MT1 and MT2, showing the position of the oligonucleotide primers used for the PCR amplification. Solid arrow denotes sequence of MT1 primers; dotted arrow denotes sequence of MT2 primers. B, agarose gel electrophoresis of PCR-amplified products from Fisher-344, Sprague-Dawley, and Wistar rat caudal artery mRNA. The single bands at approximately 366 bp for the MT1 and 264 bp for the MT2 melatonin receptors correspond to the expected length of the cDNA products produced by the oligonucleotide primers for these receptors after a second amplification using nested primers.
In Situ Hybridization.
Sprague-Dawley rats were sacrificed during the light period, and the caudal artery was isolated, embedded in Tissue-Tek OCT compound, and immediately frozen in isopentane in dry ice. Antisense and sense oligonucleotide probes designed against specific sequences of MT1 and MT2 receptor mRNA (Hunt et al., 2001) were synthesized at the Northwestern University Biotechnology Facility. Probes were tailed on the 3-prime end with33P-tagged dATP (Amersham Biosciences, Piscataway, NJ). Each oligonucleotide probe was incubated with [33P]dATP in a 1:5 ratio in the presence of terminal transferase for 1 h at 37°C and purified using Nick columns (Amersham Biosciences). Cryostat sections of caudal artery were hybridized with 33P-labeled oligoprobes following methods reported previously (Dubocovich et al., 1998; Hunt et al., 2001). The hybridization signal was detected either by apposing the artery sections on the slides to a Kodak film or by dipping them in NTB-2 emulsion (Eastman Kodak, Rochester, NY) and developing in D19 after 4 to 6 weeks. To determine the selectivity of the MT1 and MT2 oligonucleotide probes used to hybridize to target mRNAs,33P-labeled antisense oligonucleotide probe (MT1 and MT2) was hybridized to tissue sections under three experimental conditions: first, alone to determine the total hybridization signal; second, in the presence of 100 times excess of nonlabeled heterologous sense probe to block cross-hybridization; and third, in the presence of 100 times excess of nonlabeled homologous sense probes to determine the extent of nonspecific hybridization (Hunt et al., 2001).
Measurement of Contractile Force.
Four-month-old Fisher-344 male rats were sacrificed and caudal arteries removed. Proximal segments of the arteries (3 mm in length) were denuded of endothelium by intimal rubbing and then mounted on platinum wires in oxygenated 37°C Krebs' solution: 122 mM NaCl, 5.2 mM KCl, 1.6 mM CaCl2, 1.2 mM KH2PO4, 25.5 mM NaHCO3, 1.2 mM MgSO4, 0.027 mM disodium EDTA, and 11.5 mM glucose. Isometric contractions were recorded using Fort 10 force transducers and MacLab analog to digital converter system (World Precision Instruments, New Haven, CT). After 1 h, tissues were stretched to 1-g resting tension. Maximum contraction of each tissue was determined by exposure to 100 μM phenylephrine. Arterial segments were precontracted to approximately 10 to 20% of this maximum using 0.1 to 1 μM phenylephrine and then increasing concentrations of melatonin (0.1 nM–10 μM) were added cumulatively. Responses were measured as grams of contraction above the initial control response to phenylephrine. Some tissues were incubated with 4P-PDOT (10 nM), 4P-ADOT (10 nM and 3 μM), or luzindole (3 μM) for 10 min before and during exposure to melatonin. The absence of functional endothelium was verified in each segment by the lack of relaxation to 1 μM acetylcholine (<15%) after precontraction with 0.7 μM norepinephrine.
Data Analysis.
In some cases, the contractile responses in grams were normalized as a percentage of the maximum response observed with melatonin (100%). Theoretical values for EC50 and maximum response (Emax) were calculated from sigmoidal concentration-response curves by nonlinear regression using GraphPad Prism (GraphPad Software, San Diego, CA). The mean EC50 values with corresponding 95% confidence intervals were determined from values obtained in individual experiments. The concentration ratio of EC50values for melatonin in the presence and absence of antagonist was determined to calculate the apparent dissociation constant (KB) of the antagonist according to the equation pKB = log (concentration ratio − 1) − log [antagonist] (Dubocovich et al., 1997). Unless otherwise indicated, results are expressed as mean ± S.E.M., and n refers to the number of animals tested. Statistical significance was determined using Student's unpairedt test with P < 0.05.
Drugs.
Melatonin (N-acetyl-5-methoxytryptamine), 6-chloromelatonin, phenylephrine hydrochloride, and acetylcholine chloride were purchased from Sigma-Aldrich (St. Louis, MO). Luzindole (2-benzyl-N-acetyltryptamine) and 4-phenyl-2-propionamidotetraline (4P-PDOT) were purchased from Tocris Cookson (St. Louis, MO). 4-Phenyl 2-acetamidotetraline (4P-ADOT) was synthesized in the Department of Medicinal Chemistry, University of Groningen (Groningen, The Netherlands). Luzindole, 4P-ADOT, and 4P-PDOT are melatonin receptor-specific (Dubocovich et al., 1998). Their affinities on the hMT1 and hMT2 melatonin receptors expressed in mammalian cell lines were verified for each new batch. Drugs were dissolved in double distilled water unless otherwise indicated. Stock solutions (10 mM) of melatonin and 6-chloromelatonin were made in 20% ethanol and then diluted in Krebs' solution to appropriate concentrations. Luzindole (1 mM), 4P-ADOT (1 mM), and 4P-PDOT (1 mM) were prepared in 50% ethanol and then diluted in Krebs' solution to appropriate concentrations. The final ethanol concentrations in the tissue bath did not affect contractility.
Results
MT1 and MT2 Melatonin Receptor mRNA Expression in Rat Caudal Artery.
The expression of melatonin receptors mRNA in caudal arteries was determined by RT-PCR and in situ hybridization autoradiography using MT1 and MT2 melatonin receptor-specific oligonucleotides. Figure 1A shows the nucleotide sequence of the MT1 and MT2 melatonin receptors, as well as the specific sequences of the forward and reverse primers used for the first and second (nested primers) round of amplification. PCR products of the expected size, corresponding to the cDNA sequence of both receptors were amplified from Fisher-344, Sprague-Dawley, and Wistar rat caudal arteries after two rounds of amplification (Fig. 1B). For the MT1 receptor, a band of 431 bp was observed after the first amplification, and the corresponding band of 366 bp was observed after reamplification with the nested primers. For the MT2 receptor, no visible bands were observed after the first round, but nested primers, specific for the rat MT2 receptor, amplified a product of the expected size (264 bp). Nucleotide sequence of the PCR products confirmed that they are 100% homologous to the published rat MT1 and MT2 melatonin receptor sequences (accession nos. U14409 and U28218, respectively)
We next localized melatonin receptor mRNA in the Sprague-Dawley rat caudal artery by in situ hybridization using33P-labeled oligonucleotide probes complementary to target mRNA sequences of either the rat MT1 or MT2 melatonin receptors. Figure2 shows the specificity of the oligonucleotides used as probes in the rat caudal arteries determined by in situ hybridization autoradiography using33P-labeled oligonucleotides in the presence of an excess of unlabeled sense homologous or heterologous probes as described previously (Hunt et al., 2001). The hybridization signal observed with antisense 33P-labeled MT1 (Fig. 2A) or MT2 probes (Fig. 2B) was not altered in the presence of a 100-fold excess of nonlabeled heterologous sense MT2 (Fig. 2C) or MT1 (Fig. 2D) oligonucleotides, respectively. In contrast, a 100-fold excess of nonlabeled homologous sense MT1 (Fig. 2E) or MT2 (Fig.2F) oligonucleotides completely blocked hybridization of the corresponding labeled probe. Hybridization signal with sense oligonucleotide probes was negligible (data not shown). Together, these results confirm the specificity of the antisense MT1 and MT2 oligonucleotide probes for their mRNA targets and the lack of cross-hybridization.

Selective hybridization of 33P-labeled MT1 and MT2 melatonin receptor oligonucleotides probes to rat caudal arteries. Rats were maintained on a 12-h light/dark cycle and sacrificed during the light period. Rat caudal arteries were dissected, immediately frozen in OCT, and processed for in situ hybridization with antisense 33P-labeled oligonucleotide probes. A, C, and E, autoradiograms showing hybridization of 33P-labeled antisense oligonucleotides specific for the MT1 melatonin receptor to rat caudal artery sections in the absence (A) or presence of unlabeled heterologous MT2 (C) or homologous MT1 (E) sense oligonucleotides. B, D, and F, autoradiograms showing hybridization of 33P-labeled antisense oligonucleotides specific for the MT2 melatonin receptor to rat caudal artery sections in the absence (B) or presence of unlabeled heterologous MT1 (D) or homologous MT2 (F) sense oligonucleotides. Scale bar, 1 mm.
We next determined the localization of the hybridization signal in cross sections of rat caudal arteries by emulsion autoradiography (Fig.3). MT1 melatonin receptor oligonucleotide probes hybridized primarily to the muscle layer (media > intima > adventitia) of the rat caudal artery (Fig. 3A). Antisense oligonucleotides specific for the MT2 melatonin receptor mRNA hybridized to all three layers of the rat caudal artery (Fig. 3B).
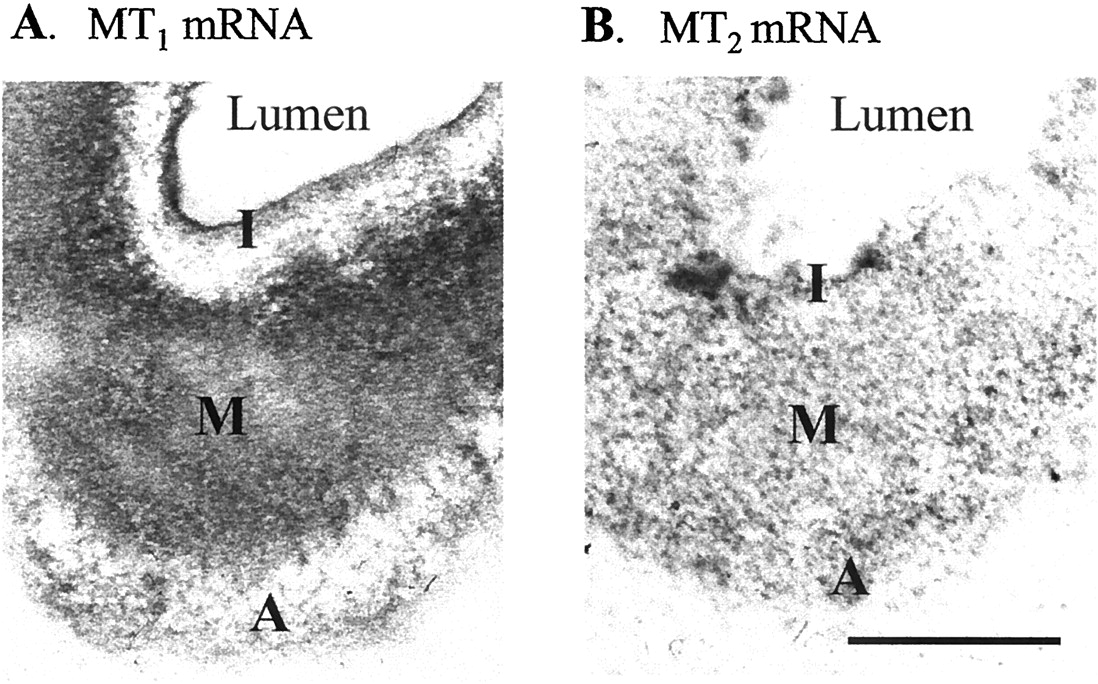
Hybridization of MT1 (A) or MT2 (B) melatonin receptor-specific 33P-labeled antisense oligonucleotide probes to sections of the rat caudal artery. MT1-specific oligonucleotide probe hybridized primarily to the muscle layer [media (M) > intima (I) > adventitia (A)]. Hybridization signal specific for the MT2 melatonin receptor was found in the three layers of the rat caudal artery. Scale bar, 100 μm.
Melatonin Receptor-Mediated Contractile Responses in Rat Caudal Artery.
In male Fisher rat caudal artery segments, melatonin by itself did not affect the resting tension (data not shown). However, melatonin significantly enhanced vascular tone when added cumulatively to tissues precontracted with phenylephrine (Fig.4). The vasocontractile responses were maximal within 3 to 5 min of melatonin addition and sustained throughout the duration of melatonin exposure.
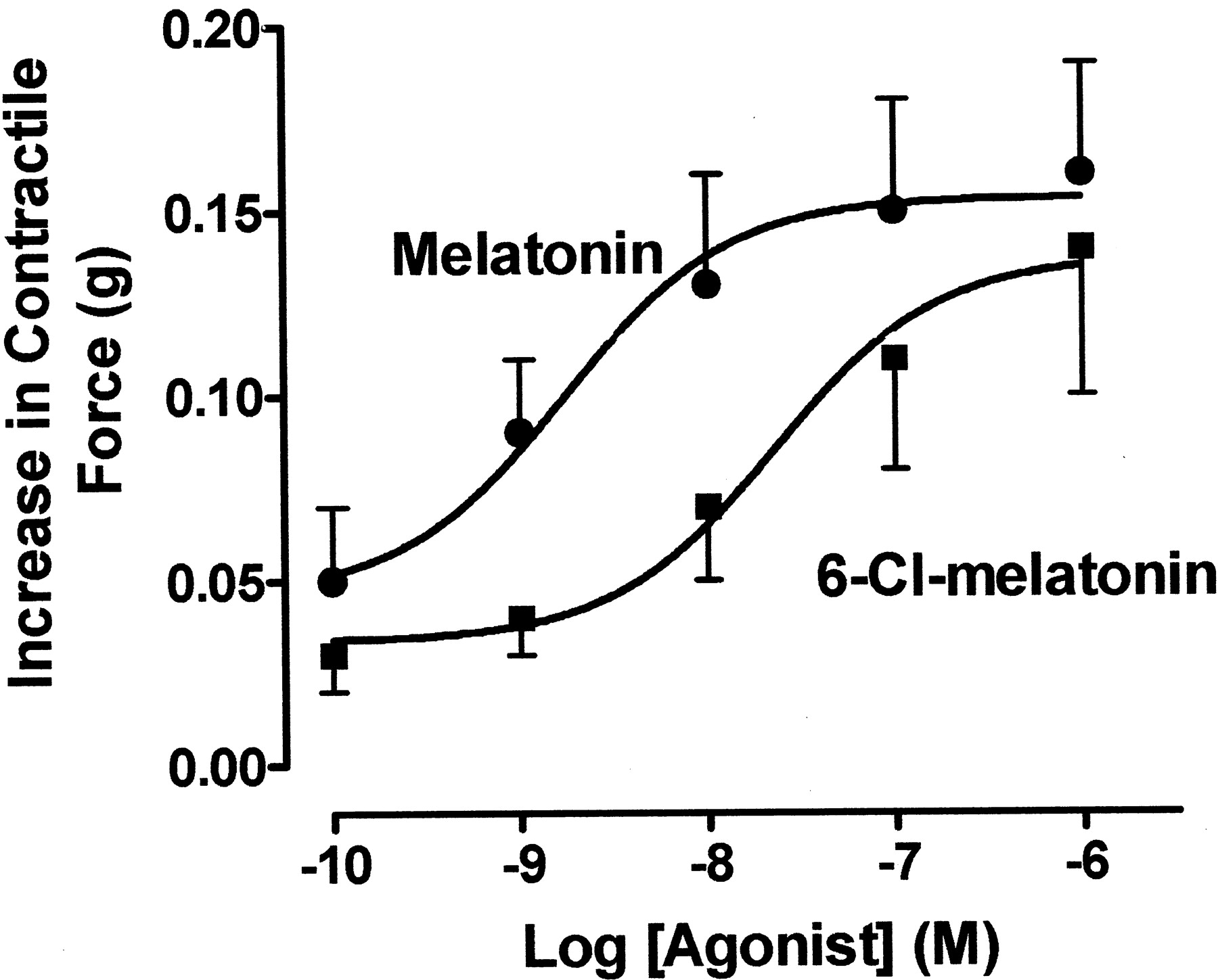
Effect of melatonin receptor agonists on isolated segments of male rat caudal arteries precontracted with phenylephrine. The ordinate represents increases in contractile response expressed in grams above control response to phenylephrine (0.1–1 μM), induced by melatonin and 6-chloromelatonin. Each agonist was added cumulatively to tissues contracted to 10 to 20% of maximum contraction with phenylephrine. Values shown represent the mean ± S.E.M. of four to six experiments.
Pharmacological characterization of the melatonin receptor-mediated arterial contraction was conducted using melatonin and 6-chloromelatonin. Melatonin and 6-chloromelatonin (0.1 nM–10 μM) potentiated phenylephrine-induced contraction in a concentration-dependent manner (Fig. 4) with potencies (EC50) of 1.6 and 17 nM, respectively.
We next compared the ability of the competitive antagonists 4P-ADOT, 4P-PDOT, and luzindole, at concentrations selective for either the MT2 (10 nM) or both the MT1and MT2 (3 μM) receptors (Dubocovich et al., 1997, 1998), to potentiate or block the contraction elicited by a single melatonin (10 nM) concentration in phenylephrine-treated caudal arteries. 4P-ADOT and 4P-PDOT, when tested alone did not affect arterial tone or phenylephrine-induce contraction, excluding a possible partial agonistic effect on either melatonin receptor (Browning et al., 2000). However, in the presence of a 10 nM concentration of either 4P-ADOT or 4P-PDOT, melatonin (10 nM) markedly potentiated the phenylephrine-induced arterial contraction (Fig.5A). In contrast, concentrations of either 4P-ADOT (3 μM) or luzindole (3 μM) that block MT1 and MT2 melatonin receptors nonselectively (Dubocovich et al., 1997, 1998) significantly inhibited the contractile effect of 10 nM melatonin (Fig. 5B). This result is consistent with blockade of contraction mediated by MT1 melatonin receptors.

Effect of the competitive melatonin receptor antagonists 4P-ADOT, 4P-PDOT, and luzindole on melatonin-mediated contraction in male rat caudal arteries. The effects of melatonin (10 nM) in the presence and absence of the antagonist 4P-ADOT (10 nM and 3 μM), 4P-PDOT (10 nM), and luzindole (3 μM) are expressed as percentage of the maximal melatonin response attained in each individual experiment. The antagonists were present 10 min before and during exposure to melatonin (10 nM). Values represent means ± S.E.M. of five to seven experiments. ∗, P < 0.05 compared with melatonin alone.
To further support a role for MT2 receptor blockade in vascular relaxation, we determined the effect of an MT2-selective concentration of 4P-PDOT (10 nM) on melatonin-mediated, concentration-dependent contractile responses (Fig.6). Melatonin (0.1 nM–10 μM) potentiated phenylephrine-mediated contractions in caudal arteries in a concentration-dependent manner with an pEC50value of 8.6 and a maximal effect at 10 nM (Fig. 6A; Table1). 4P-PDOT at 10 nM shifted to the left the melatonin concentration-effect curve and increased the maximal response to melatonin with no change in potency (Fig. 6A; Table 1). On the contrary, the nonselective MT1/MT2 antagonist luzindole (3 μM) shifted the melatonin concentration-response curve to the right in a competitive manner, significantly decreasing the potency of melatonin to contract the artery (Table 1), with an estimated affinity constant (KB) of 151 nM (pKB = 6.8). Luzindole also increased the maximal response to melatonin (Fig. 6B; Table 1).
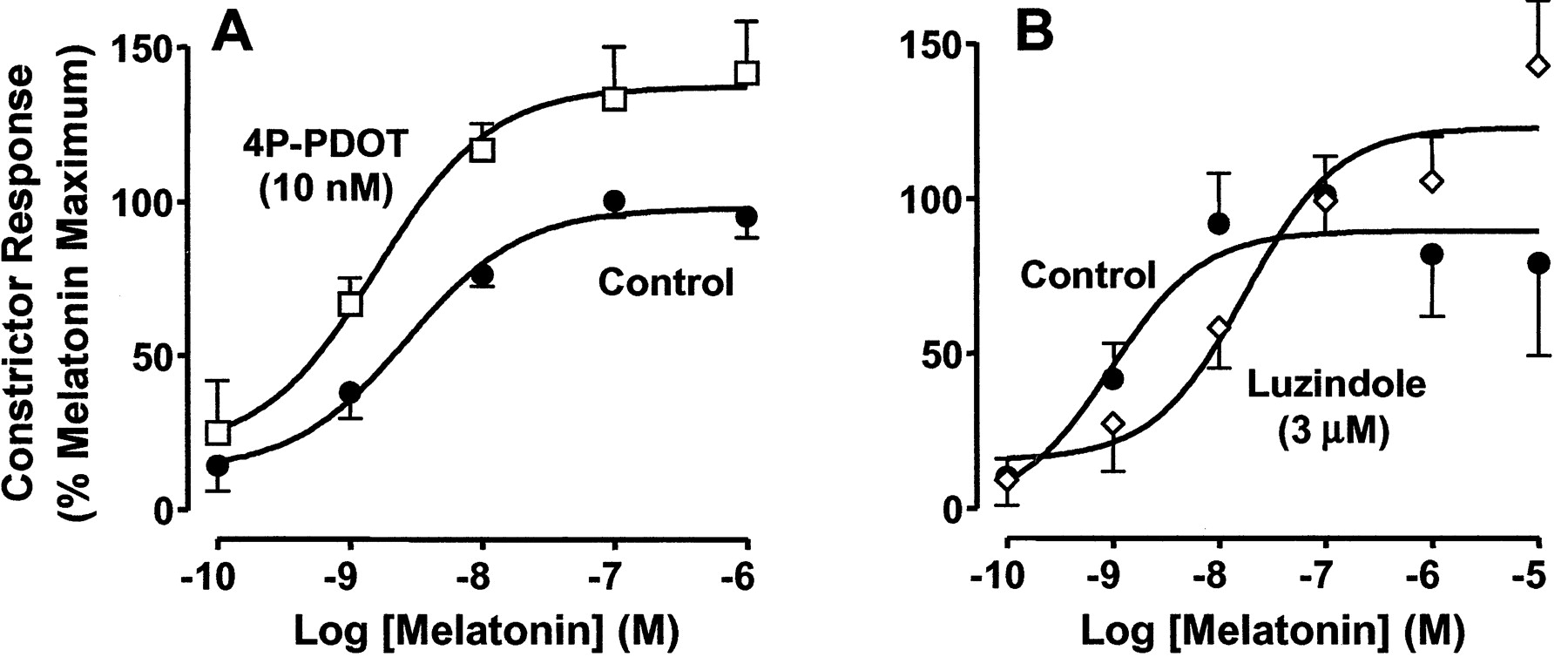
Effect of the melatonin receptor antagonists, luzindole and 4P-PDOT on melatonin-mediated contraction in male rat caudal arteries. The effects of melatonin in the presence and absence of antagonist are expressed as percentage of the maximal melatonin response attained in each individual experiment. Melatonin (0.1 nM–10 μM) was added cumulatively. The antagonists were added 10 min before and were present during exposure to melatonin. Values represent means ± S.E.M. of five to seven experiments.
MT2 and MT1/MT2 antagonists on efficacy and maximum contractile effect of melatonin
Discussion
This study demonstrates expression of MT2melatonin receptor mRNA and establishes MT2-mediated functional responses in the rat caudal artery. The mRNA for both the MT1 and MT2 melatonin receptors was localized in the smooth muscle layer. This finding is consistent with the presence of pharmacologically distinct melatonin receptors (MT1 and MT2) that modulate contractile responses to melatonin in caudal artery segments denuded of endothelium. These results suggest that in the arterial smooth muscle, activation of the MT2 melatonin receptor mediates vasodilation, whereas activation of the MT1receptor enhances vasoconstriction.
Herein, we report for the first time expression of vascular MT2 melatonin receptor mRNA using caudal arteries of three rat strains, i.e., Fisher-344, Sprague-Dawley, and Wistar. MT1 melatonin receptor mRNA from Wistar rat caudal arteries was previously detected by RT-PCR (Ting et al., 1997), but these authors were unable to amplify mRNA for the MT2 melatonin receptor. The discrepancy with our results could be attributed to differences in experimental conditions. First, our RT-PCR studies used RNA isolated from caudal arteries pooled from six to eight rats, providing a higher number of MT2 mRNA copies. In the previous study, RNA was isolated and reverse transcribed from arteries of individual animals (Ting et al., 1997). Second, in the present study, the sensitivity of mRNA detection was increased by using the One-Step PCR technique (Limbach et al., 1999). Detection of residual genomic DNA can be excluded because samples were DNase-treated, and no amplification was observed in negative controls consisting of mRNA samples that were not reverse transcribed, or in samples from a rat tissue that does not express MT1/MT2 receptors (i.e., mammary gland; data not shown). Amplification of MT1 and MT2 cDNAs from three strains of rats further supports the validity of the assay.
Hybridization of complementary oligonucleotide probes to both MT1 and MT2 melatonin receptor mRNAs also indicate expression of the both receptor types in the artery. MT1 and MT2melatonin receptor mRNAs were localized by in situ hybridization to distinct layers of the arterial wall using33P-oligonucleotide probes (this study) and digoxigenin-labeled probes (data not shown). The hybridization of both antisense oligonucleotide probes was specific as determined using sense probes and competition of 33P-labeled antisense probes with homologous and heterologous unlabeled oligonucleotides. Using the same techniques, MT1 and MT2 mRNA was localized to human cerebellum (Al-Ghoul et al., 1998) and mouse (Dubocovich et al., 1998) and rat (Hunt et al., 2001) suprachiasmatic nuclei. The high level of mRNA observed by in situ hybridization studies compared with the need for extensive rounds of amplification to demonstrate expression of MT2 receptors by RT-PCR is intriguing. The high sensitivity of our in situ hybridization procedures could be attributed to the use of small oligonucleotide probes instead of riboprobes and to the immediate preparation and processing of arterial sections for in situ hybridization (Al-Ghoul et al., 1998).
MT1 receptor mRNA was localized primarily in the smooth muscle layer of the caudal artery, whereas MT2 receptor mRNA appeared more evenly distributed throughout the vessel wall. Expression of receptor protein in vascular smooth muscle is indicated by specific binding of 2-[125I]iodomelatonin, a nonselective MT1/MT2 radioligand, in the medial layer of caudal and cerebral arteries (Viswanathan et al., 1990,1992). This binding, however, likely reflects MT1melatonin receptors because the density of endogenous MT2 receptors is generally too low in mammalian tissues to be detected by receptor autoradiography (Liu et al., 1997;Dubocovich et al., 1998). Although in the adventitia we found primarily mRNA for MT2 receptors, Savaskan et al. (2001)observed immunoreactivity in the adventitia layer of human superficial and intrahippocampal arteries using an antibody directed against MT1 melatonin receptors. The function of putative melatonin receptors in the adventitia is not known; however, it is interesting that levels of immunoreactivity were significantly higher in arteries from Alzheimer patients (Savaskan et al., 2001). The localization of MT2 receptor mRNA in the intimal layer is intriguing as well. Recently, MT2melatonin receptors in the endothelium were shown to inhibit leukocyte rolling (Lotufo et al., 2001) and may also be involved in the modulation of vascular tone (Geary et al., 1998).
Based on agonist and antagonist data from male Fisher-344 rat caudal arteries, denuded of endothelium, activation of MT1 melatonin receptors seems to mediate potentiation of smooth muscle contraction. Melatonin was 10-fold more potent than 6-chloromelatonin to enhance contraction, in agreement with the relative agonist affinities at recombinant hMT1 melatonin receptors (Dubocovich et al., 1997). In Wistar rat caudal artery, however, using electrically evoked contraction in an isometric preparation (Ting et al., 1997) or phenylephrine-mediated contraction in endothelium-denuded, pressurized arteries (Bucher et al., 1999), melatonin and 6-chloromelatonin were equipotent. The discrepancy with our results could be attributed to differences in the arterial preparation and/or the relatively small differences in agonist potency. With regard to antagonists, luzindole (3 μM) inhibited contraction to melatonin with an estimated affinity constant (KB = 157 nM) that corresponds to the affinity of this ligand for recombinant hMT1 receptors (Ki = 180 nM) (Dubocovich et al., 1998). The melatonin receptor antagonists 4P-ADOT and 4P-PDOT, at MT2-selective concentrations (Dubocovich et al., 1997, 1998), did not block melatonin-mediated contraction. Along with previous pharmacological studies in Sprague-Dawley and Wistar rat caudal arteries (Krause et al., 1995; Ting et al., 1997, 1999; Bucher et al., 1999; Lew and Flanders, 1999), the present results strongly support the conclusion that melatonin-mediated potentiation of adrenergic vasocontraction involves activation of smooth muscle MT1 melatonin receptors.
The presence of functional MT2 melatonin receptors in caudal arteries denuded of endothelium is suggested by the effects of the selective MT2 receptor antagonists. 4P-ADOT and 4P-PDOT, at MT2-selective concentrations, significantly enhanced contractile responses to melatonin. The potency of melatonin to elicit contraction was not altered, as expected for an MT1-mediated response (Table 1). The enhancement of melatonin-mediated contraction in the presence of MT2 antagonists could be explained if activation of MT2 melatonin receptors were to mediate vasorelaxation. Similar results were obtained with 4P-PDOT in caudal arteries of female Fisher-344 rats; and in fact, the effect of the MT2 antagonist was increased after in vivo estrogen exposure (Doolen et al., 1999). The MT1/MT2-nonselective antagonist luzindole also potentiated melatonin-induced contraction, which is consistent with blockade of MT2receptors-mediating dilatation. Unfortunately, without selective MT1 antagonists to block melatonin-induced vasocontraction, it is not possible to directly measure the postulated MT2-mediated vasorelaxation in the caudal artery. It should be noted that potentiation of contraction by high concentrations (100 nM or above) of the mixed antagonist luzindole or the MT2-selective amidotetralines could result in part from their inverse agonist properties at MT1constitutively active melatonin receptors in caudal artery (Browning et al., 2000; Masana and Dubocovich, 2001; Ersahin et al., 2002). Inverse agonists such as luzindole or 4P-PDOT could shift the equilibrium of constitutive active receptors in caudal artery (Ersahin et al., 2002) to the uncoupled form, making more receptors available for activation by high concentrations of melatonin leading to potentiation of vasocontraction. It is unlikely that the potentiation observed at higher melatonin concentrations is due to 4P-PDOT partial agonist properties at the MT1 receptor (Ting et al., 1999), because a partial agonist effect of 4P-PDOT in this model should have been more evident at lower concentrations of melatonin. Furthermore, 4P-PDOT at 10 nM shows no affinity for the MT1 melatonin receptor (Dubocovich et al., 1998).
In conclusion, both MT1 and MT2 melatonin receptors are present and functional in vascular tissue. Melatonin seems to modulate vascular smooth muscle tone via activation of MT1melatonin receptors that mediate contraction and MT2 receptors that mediate vasorelaxation. Although the functional significance of this finding remains to be elucidated, the presence of multiple receptor subtypes with opposing actions is a common scheme in the vasculature. Because regulation of blood flow in the tail is an important mechanism for thermoregulation in the rat, the ability to regulate two different melatonin receptors with opposing vascular actions could provide greater flexibility in adapting body temperature to circadian and seasonal changes. In humans, melatonin selectively vasodilates (and therefore increases heat loss) in distal skin regions having mainly thermoregulatory functions, such as fingertips and toes, whereas it does not affect proximal regions, such as thorax and abdomen (Krauchi et al., 2000). In this way, responses to melatonin may be involved in the fine-tuning of vascular tone in selective vascular beds, as circulating melatonin levels rise and fall throughout the night.
Acknowledgments
We thank Dr. Linda VanEldik for the use of laboratory facilities to perform the RNA isolation and RT-PCR of melatonin receptors.
Footnotes
-
↵1 Current address: Department of Pharmacology, University of Colorado Health Sciences Center, Denver, CO 80262.
-
↵2 Current address: Department of Surgery, Shock and Trauma Institute, Loyola University Medical School, Maywood, IL 60153.
-
This research was supported by American Heart Association Grant-in-Aid 9708073 (to M.I.M.), U.S. Public Health Service Grants MH-52685 and MH-42922 (to M.L.D.), and National Institutes of Health R01 HL-50775 (to S.P.D.) and T32 ES 07124 (to W.A.G.).
- Abbreviations:
- 4P-ADOT
- 4-phenyl-2-acetamidotetralin
- 4P-PDOT
- 4-phenyl-2-propionamidotetralin
- RT-PCR
- reverse transcription-polymerase chain reaction
- PCR
- polymerase chain reaction
- bp
- base pair(s)
- hMT
- human melatonin receptor
- Received December 7, 2001.
- Accepted February 9, 2002.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}