


Abstract
Agonists at δ, μ, and κ opioid receptors produce interacting effects in rodents and nonhuman primates. To further evaluate the determinants of these interactions, this study examined the effects of mixtures of δ + μ and δ + κ agonists in rhesus monkeys (n = 4–5) using two behavioral procedures, an assay of schedule-controlled responding for food reinforcement and an assay of thermal nociception. Results were analyzed using dose-addition analysis. In the assay of schedule-controlled responding, the δ agonist (+)-4-[(αR)-α-((2S,5R)-4-allyl-2,5-dimethyl-1-piperazinyl)-3-methoxy-benzyl]-N,N-diethyl-benzamide (SNC80); the μ agonists methadone, fentanyl, morphine, and nalbuphine; and the κ agonists (5α,7α,8β)-(–)-N-methyl-N-(7-(1-pyrrolidinyl)-1-oxaspiro(4,5)dec-8-yl) benzeneacetamide (U69,593) and bremazocine all dose dependently decreased rates of food-maintained responding when administered alone. Fixed ratio mixtures of SNC80 + μ agonists produced additive or subadditive effects, whereas SNC80 + κ agonist mixtures produced only additive effects. In the assay of thermal nociception, SNC80 produced no measurable effects when administered alone, whereas μ and κ agonists produced dose-dependent antinociception. SNC80 + μ agonist mixtures produced superadditive effects manifested as leftward shifts in μ agonist dose-effect curves. This synergism was antagonized by the δ-selective antagonist naltrindole, suggesting that SNC80-induced enhancement of μ agonist antinociception was δ receptor-mediated. SNC80 did not enhance the antinociceptive effects of the highly selective κ agonist U69,593, and it produced only a marginal enhancement of antinociception produced by the less selective κ agonist bremazocine. These results suggest that δ agonists may selectively enhance the antinociceptive effects of μ agonists in rhesus monkeys. These results also confirm that opioid agonist interactions may depend on the receptor selectivity and relative doses of the agonists and on the experimental endpoint.
Biochemical and behavioral evidence indicates the existence of three opioid receptor types, the δ, μ, and κ receptors (Martin et al., 1976; Lord et al., 1977; Evans et al., 1992; Thompson et al., 1993; Yasuda et al., 1993). Agonists selective for δ, μ, and κ receptors produce distinct profiles of physiological and behavioral effects (Gutstein, 2001). In addition, selective δ, μ, and κ agonists may also produce interacting effects. Studies of opioid receptor interactions have been conducted primarily with δ and μ agonists in rodents using assays of antinociception, and results from these studies suggest that the nature of the interaction depends on such variables as the particular agonists used, the relative doses tested, and the behavioral endpoint (Heyman et al., 1989; Jiang et al., 1990; Adams et al., 1993). For example, the peptidic δ agonists [d-Pen2,d-Pen5]-enkephalin (DPDPE) and deltorphin II enhanced the effects of some intermediate-efficacy μ agonists (morphine, normorphine, and codeine), but not of higher efficacy μ-agonists (PL017, fentanyl, or sufentanil), in an assay of thermal nociception in mice (Heyman et al., 1989; Jiang et al., 1990). In a subsequent study, DPDPE enhanced the antinociceptive effects of morphine in a cold-water tail-flick test but not in a warm-water tail-flick test in rats (Adams et al., 1993). These studies have also demonstrated the utility of dose-addition analysis in the evaluation of agonist interactions. Dose-addition analysis permits an assessment of the degree to which a drug combination produces effects that are additive, superadditive, or subadditive, and results can be displayed using isobolograms (Wessinger, 1986; Woolverton, 1987; Tallarida, 2000). Using dose-addition analysis, for example, it was found that a mixture of 20% DPDPE + 80% morphine produced superadditive antinociceptive effects in the cold-water tail-flick test in rats, but a mixture of 40% DPDPE + 60% morphine produced only additive effects (Adams et al., 1993).
Interactions between δ and μ agonists have also been reported for other behavioral endpoints in rodents, such as bladder motility, convulsions, Straub tail, and respiratory depression (Sheldon et al., 1989; O'Neill et al., 1997; Su et al., 1998). Importantly, δ and μ agonists often either do not modify each other's nonantinociceptive effects or attenuate each other's nonantinociceptive effects. For example, the nonpeptidic δ agonist BW373U86 produced convulsions and the μ agonist fentanyl produced Straub tail in mice, and both compounds produced antinociceptive effects in a hot-plate assay of antinociception. Relative to the effects of either drug alone, combinations of BW373U86 and fentanyl produced greater antinociception but fewer convulsions and a lower incidence of Straub tail (O'Neill et al., 1997). BW373U86, DPDPE, and deltorphin II also attenuated the respiratory depressant effects of the μ agonist alfentanil in rats (Su et al., 1998). These findings suggest that mixtures of δ and μ agonists may produce robust analgesic effects with fewer undesirable effects than highly selective δ or μ agonists.
Interactions between δ and μ agonists might be expected to depend critically on the anatomical relationship of δ and μ receptors. Because there are species differences in both the density and anatomical distribution of opioid receptors (Mansour et al., 1986), it may be especially important to assess the degree to which δ/μ agonist interactions observed in rodents extend to nonrodent species. Toward this end, two studies to date have examined interactions between δ and μ agonists in nonhuman primates (Dykstra et al., 1993, 2002). In these studies, both BW373U86 and the structurally related and more selective δ agonist SNC80 enhanced the antinociceptive effects of several μ agonists in a shock titration procedure in squirrel monkeys. As in rodents, δ/μ interactions were dependent on the doses of the agonists. However, in contrast to results obtained in rodents (Heyman et al., 1989; Jiang et al., 1990; Adams et al., 1993), δ agonists were most effective in enhancing the effects of high-efficacy μ agonists.
The purpose of the present study was to further evaluate interactions between δ and μ agonists in nonhuman primates using fixed ratio mixtures of δ and μ agonists to facilitate the application of dose-addition analysis (Tallarida, 2000). The effects of a δ agonist (SNC80) and four μ agonists that differ in relative efficacy at the μ opioid receptor (methadone, fentanyl, morphine, and nalbuphine; Emmerson et al., 1994, 1996; Gatch et al., 1995; Negus and Mello, 1999) were initially determined in an assay of schedule-controlled responding for food reinforcement in rhesus monkeys. The δ, μ, and κ opioid agonists all produce dose-dependent decreases in response rates in this procedure (Negus et al., 1993a), and as a result, this procedure was used to determine the relative potencies of the agonists and to provide an empirical basis for the relative proportions of drugs subsequently used in drug mixtures. Interactions between δ and μ agonists were then examined in an assay of thermal nociception in rhesus monkeys that has been used extensively to examine the antinociceptive effects of δ, μ, and κ opioids administered alone (Dykstra et al., 1987; Negus et al., 1993a, 1998; Gatch et al., 1995; Walker et al., 1995). Finally, interactions were examined between SNC80 and two κ opioid agonists (U69,593 and bremazocine) for comparison with the effects of δ and μ agonists.
Materials and Methods
Subjects. Five male rhesus monkeys (Macaca mulatta) were used in studies of schedule-controlled responding, and four male monkeys were used in studies of thermal nociception. Subjects weighed 4.5 to 12 kg during the course of these studies. All monkeys had prior exposure to drugs (primarily dopaminergic and opioid compounds) and to the behavioral procedures in which they were tested. The subjects were individually housed, and water was freely available. Their diet consisted of PMI Feeds Jumbo monkey diet (2–6 biscuits/day; PMI Feeds, Inc., St. Louis, MO). This diet was supplemented with fresh fruit twice daily. In addition, monkeys in the assay of schedule-controlled behavior could earn additional food pellets during experimental sessions. A 12-h light/12-h dark cycle was in effect (lights on from 7:00 AM to 7:00 PM).
Animal maintenance and research were conducted in accordance with the guidelines provided by the National Institutes of Health Committee on Laboratory Animal Resources. The facility was licensed by the United States Department of Agriculture, and protocols were approved by the Institutional Animal Care and Use Committee. The health of the monkeys was monitored daily by the staff veterinarian. Monkeys had visual, auditory, and olfactory contact with other monkeys throughout the study. Monkeys also had access to puzzle feeders, mirrors, and chew toys to provide environmental enrichment. Nature videotapes or music were played daily in all housing rooms.
Behavioral and Pharmacological Procedures. Behavioral studies were conducted using two procedures: 1) an assay of schedule-controlled responding for food presentation, and 2) an assay of thermal nociception using warm water as the noxious stimulus. These procedures have been used extensively by our laboratory to examine the effects of other opioids in rhesus monkeys (Negus et al., 1993a; Gatch et al., 1995). In each assay, the effects of the δ agonist SNC80; the μ agonists methadone, fentanyl, morphine, and nalbuphine; and the κ agonists U69,593 and bremazocine were examined alone and in various mixtures. We have reported previously that SNC80 acts as a selective δ opioid receptor agonist with relatively high efficacy at δ receptors in rhesus monkeys (Negus et al., 1998; Brandt et al., 1999). Methadone, fentanyl, morphine, and nalbuphine all produce behavioral effects mediated primarily by μ-opioid receptors in rhesus monkeys; however, these compounds differ in their relative efficacy at and selectivity for μ receptors. Both in vivo and in vitro data suggest that the rank order of both efficacy and selectivity is methadone ≥ fentanyl > morphine > nalbuphine (Emmerson et al., 1994, 1996; Gatch et al., 1995; Negus and Mello, 1999). U69,593 and bremazocine produce behavioral effects mediated primarily by κ receptors in rhesus monkeys; however, these compounds also differ in their relative selectivity for and efficacy at κ receptors (Remmers et al., 1999). Specifically, U69,593 is more selective for κ receptors and has slightly higher efficacy at κ receptors than bremazocine.
Assay of Schedule-Controlled Responding. Experiments were conducted in each monkey's home cage (dimensions: 60 × 65 × 75 cm). The home cages of all monkeys were modified to include an operant response panel (28 × 28 cm) mounted on the front wall. Three square translucent response keys (6.4 × 6.4 cm) were arranged 2.54 cm apart in a horizontal row 3.2 cm from the top of the operant panel. Each key could be transilluminated by red, green, or yellow stimulus lights. In addition, three circular translucent panels (1.9 cm in diameter) were located in a vertical column below the center response key and could be transilluminated by red, green or yellow stimulus lights. The operant panel also supported an externally mounted pellet dispenser (model G5210; Gerbrands, Arlington, MA) that delivered 1 g of fruit-flavored food pellets (P.J. Noyes Co., Lancaster, NH) to a food receptacle mounted on the cage beneath the operant response panel. The panel was controlled by a MED-PC interface and an IBM-compatible computer programmed in MED-STATE Notation (MED Associates, East Fairfield, VT).
Experiments used a multiple-cycle procedure. Each of five cycles was 15 min in length and consisted of two components: a 10-min time-out period followed by a 5-min response period. During the time-out period, no stimulus lights were illuminated and responding had no scheduled consequences. During the response period, the center key was transilluminated yellow, and the subjects could respond for up to 10 food pellets under a fixed ratio 30 schedule of reinforcement. If all 10 food pellets were earned before 5 min had elapsed, the lights were turned off, and responding had no scheduled consequences for the remainder of that response period. All monkeys were trained until they responded at rates greater than 1.0 response/s during all five cycles for 10 consecutive days, and we have shown previously that monkeys respond at relatively stable rates across successive response periods in this procedure (Negus et al., 1993a).
Sessions were conducted 5 days a week. Test sessions were usually conducted on Tuesdays and Fridays, and training sessions were conducted on Mondays, Wednesdays, and Thursdays. In addition, test sessions were conducted only after a training session during which the monkeys responded at rates greater than 1.0 response/s for all five cycles. Test sessions were identical to training sessions except that test compounds were administered using a cumulative dosing procedure, in which doses of the test drug or drug mixture were administered at the beginning of each time-out period, and each dose increased the total cumulative dose by one-fourth or one-half log unit.
Initially, complete dose-effect curves were determined for each drug administered alone, and each drug was tested twice. Tests of μ and κ agonists alone were separated by at least three days, and tests of SNC80 alone were separated by at least 7 days. Subsequently, three mixtures of SNC80 in combination with each μ agonist and each κ agonist were examined, and the proportions of each drug in the mixtures were based on the relative potency of the drugs (see “Data Analysis” for details). Thus, each cumulative dose for each mixture contained a given proportion of SNC80 and a given proportion of μ or κ agonist. Each mixture was tested once, and 1 week separated each test. Most studies were conducted in a group of five monkeys. Methadone was studied in a group of four monkeys, because one monkey was ill at the time the others were tested. Nalbuphine was also tested in a group of only four monkeys, because nalbuphine alone did not reliably decrease rates of responding in one monkey. All agonists and mixtures were tested up to doses that eliminated responding in most or all monkeys.
Assay of Thermal Nociception. The monkeys were seated in acrylic restraint chairs so that their tails hung down freely. The bottom 10 cm of each monkey's shaved tail was immersed in a thermal container of warm water. If the subject did not withdraw its tail within 20 s, the tail was removed from the water by the experimenter, and a latency of 20 s was assigned to that measurement. During each cycle of measurements, tail-withdrawal latencies were measured from four different temperatures. Usually, these temperatures were 38, 42, 46, and 50°C, and we have reported previously that tail-withdrawal latencies are inversely related to water temperature across this range of temperatures (Gatch et al., 1995). When high drug doses were tested, 54°C was used and 38°C was omitted. Temperatures above 54°C were not tested to avoid any possibility of tissue damage. The order in which the temperatures were presented varied from one set of measurements to the next according to a modified Latin square sequence. Experiments were conducted no more than twice a week. A stopwatch was used to measure and record time intervals.
Each test session consisted of multiple 30 min cycles. Before the first cycle, baseline latencies to tail withdrawal from 38, 42, 46, and 50°C water were determined. During cumulative dosing experiments, a single drug dose was administered at the start of each of five sequential 30 min cycles, and each dose increased the total cumulative dose by one-fourth or one-half log unit. Starting 20 min after each injection, tail-withdrawal latencies from different water temperatures were recorded as described above. Initially, complete dose-effect curves were determined for SNC80, methadone, fentanyl, morphine, nalbuphine, U69,593, and bremazocine administered alone, and each drug was tested twice. Tests of μ and κ agonists alone were separated by at least 3 days, and tests of SNC80 alone were separated by at least 7 days. Subsequently, three mixtures of SNC80 in combination with each μ agonist and each κ agonist were examined, and the proportions of each drug in the mixtures were identical to those examined in the assay of schedule-controlled behavior described above. Each drug mixture was tested once, and 1 week separated each test.
These studies revealed that the combination of SNC80 with μ agonists produced leftward shifts in μ agonist dose-effect curves. To assess the role of delta receptors in mediating SNC80-induced left-ward shifts in μ agonist dose-effect curves, a final study examined the effects of pretreatment with the δ-selective antagonist naltrindole (1.0 mg/kg) on the antinociceptive effects produced by methadone alone and by one mixture of SNC80 + methadone. Naltrindole was administered 30 min before the first cumulative dose of methadone or of the SNC80 + methadone mixture. We have reported previously that this dose and pretreatment time for naltrindole produces δ-selective antagonist effects in rhesus monkeys (Negus et al., 1998).
Data Analysis. For the assay of schedule-controlled responding, operant response rates from each cycle were converted to percentage of control using the average rate from the previous training day as the control value. The ED50 for each individual agonist was defined as the dose of agonist that produced a 50% decrease in the percentage of control rate of responding. Individual ED50 values were calculated by interpolation when only two data points were available (one below and one above 50% control response rate) or by linear regression when at least three data points were available on the linear portion of the dose-effect curve. For drug mixtures, a related quantity, Zmix, was also calculated for each monkey. Zmix was defined as the total drug dose (i.e., dose SNC80 + dose other drug in mixture) that produced a 50% decrease in responding.
For the assay of thermal nociception, tail-withdrawal data were analyzed as described previously using a method that emphasizes drug-induced shifts in complete temperature-effect curves rather than drug-induced changes in tail-withdrawal latencies at any one temperature (Negus et al., 1993b; Gatch et al., 1996). For each set of tail-withdrawal latency measurements collected during any one cycle of a test session, a temperature-effect curve was constructed that related tail-withdrawal latency to water temperature. A T10 value was determined from each temperature-effect curve by fitting a line to the two points that fell immediately above and below 10 s and by calculating the temperature in degrees centigrade that corresponded to a tail-withdrawal latency of 10 s. For example, if tail-withdrawal latencies from 38, 42, 46, and 50°C were 20, 15, 5, and 0.1 s, respectively, then the equation of the line between (42°C, 15 s) and (46°C, 5 s) would be determined and used to interpolate the temperature associated with a tail-withdrawal latency of 10 s. In this case, the T10 value would be 44°C. On some occasions (i.e., with the highest doses of methadone, fentanyl, morphine, and U69,593 in some monkeys), tail withdrawal latencies were greater than 10 s at all water temperatures up to and including 54°C, and as noted above, temperatures above 54°C were not tested to avoid any possibility of tissue damage. In these cases, the T10 was estimated using the experimentally determined value at 54°C and a value of “0” at 58°C (i.e., the next 4°C increment). Thus, the maximum possible T10 value was 56°C (i.e., a 20-s latency from 54°C water). The T10 from each monkey's baseline test cycle was subtracted from the T10 determined from each subsequent test cycle, which provided a measure of change relative to the baseline of each animal, or ΔT10. Antinociception was operationally defined as rightward shifts in the temperature-effect curves and increases in the ΔT10 values. The dose of each drug that produced a ΔT10 of 5°C (ED5°C) was interpolated from the linear portion of the dose-effect curve for each monkey. For drug mixtures, Zmix was also calculated for each monkey as the total drug dose (i.e., dose SNC80 + dose other drug in mixture) that produced a ΔT10 of 5°C. Because drug doses were incremented on a logarithmic scale, ED50, ED5°C, and Zmix values were converted to their log values for calculation of means and standard errors and for statistical analysis and then converted back to linear values for presentation in tables and text.
Interactions between δ agonists and μ and κ agonists were assessed using both graphical and statistical approaches (Wessinger, 1986; Woolverton, 1987; Tallarida, 2000). Graphically, mean ED50 and ED5°C values (95% confidence limits) for SNC80 administered either alone or as part of a mixture were plotted as a function of the ED50 or ED5°C value of the other drug in the mixture. This data presentation format is known as an isobologram, and the line in an isobologram that connects the data points for each drug alone shows predicted data points for drug mixtures assuming additivity. Points that fall above the line of additivity (away from the origin) are suggestive of subadditivity, whereas points that fall below the line (toward the origin) are suggestive of superadditivity.
Statistical evaluation of drug interactions was accomplished by comparing the experimentally determined ED50 or ED5°C values for each mixture (Zmix) with predicted additive ED50 or ED5°C values (Zadd) as described by Tallarida (2000). Zmix values were determined empirically as described above. For the assay of schedule-controlled responding, in which all drugs were equieffective, Zadd values were calculated individually for each monkey from the following equation: Zadd = fA + (1 – f)B, where A is the ED50 for SNC80 alone, B is the ED50 for the other drug alone, and f is the fractional multiplier of A in the computation of the additive total dose. Any choice of f is related to the proportion of drug A (ρA) in a mixture according to the following equation: ρA = fA/Zadd. The present study examined effects produced by mixtures in which f = 0.25, 0.5, and 0.75. Thus, f = 0.25 leads to ρA = A/(A/3 + B), and the ratio of the SNC80 concentration to the concentration of the other drug is [(A/B) ÷ 3]:1; f = 0.5 leads to ρA = A/(A + B), and the ratio of the SNC80 concentration to the concentration of the other drug is A/B:1; and f = 0.75 leads to ρA = 3A/(3A + B), and the ratio of the SNC80 concentration to the concentration of the other drug is [(A/B) × 3]:1.
For the assay of thermal nociception, SNC80 alone was ineffective, and the hypothesis of additivity predicts that SNC80 would not contribute to the effects of a mixture of SNC80 in combination with a μ or κ agonist. As a result, the equation for Zadd reduces to Zadd = B/ρB. Mean predicted and experimental values were compared by one-way analysis of variance, and the criterion for a significant effect was p < 0.05.
Drugs. SNC80 (free base) and bremazocine HCl were synthesized by Drs. Kenner C. Rice and John E. Folk (National Institutes of Health, Bethesda, MD). Methadone HCl, fentanyl HCl, morphine sulfate, and naltrindole HCl were provided by the National Institute on Drug Abuse (Bethesda, MD). U69,593 and nalbuphine HCl were purchased from Sigma-Aldrich (St. Louis, MO). SNC80 was dissolved in 3% lactic acid to a final concentration of 50 mg/ml, and dilutions were made with sterile water. All other drugs were dissolved in sterile water. All drugs were administered i.m. in the thigh, and doses were determined based on the free base or salt forms listed above.
Results
Assay of Schedule-Controlled Responding
Control Response Rates and Effects of Opioid Agonists Alone. The average control response rate ± S.E.M., determined on the days before test days throughout the study, was 2.27 (±0.21) responses/s. Response rates were similar during each of the five cycles of the multiple cycle sessions (2.21 ± 0.27, 2.27 ± 0.24, 2.29 ± 0.24, 2.26 ± 0.19, and 2.32 ± 0.13 responses/s).
The δ agonist SNC80; the μ agonists methadone, fentanyl, morphine, and nalbuphine; and the κ-agonists U69,593 and bremazocine each produced a dose-dependent decrease in the percentage of control rates of responding (Fig. 1), and the ED50 value for each compound is shown in Table 1. The potency order was bremazocine > fentanyl = U69,593 > nalbuphine = SNC80 > methadone ≥ morphine. Relative potencies for each drug were determined in comparison to SNC80, and these relative potencies were subsequently used to determine the relative proportions of SNC80 and the μ or κ agonist in the subsequent drug mixtures.

Effects of SNC80, methadone, fentanyl, morphine, nalbuphine, U69,593 and bremazocine administered alone on schedule-controlled responding. Abscissa, dose of opioid agonist in milligrams per kilogram (log scale). Ordinate, percentage of control response rate. All points show mean data from four or five monkeys. ED50 values for each drug are shown in Table 1.
ED50 values (mg/kg; 95% CL) for the δ agonist SNC80; the μ agonists, methadone, fentanyl, morphine, and nalbuphine; and the κ agonists, U69,593 and bremazocine, in an assay of schedule-controlled responding for food reinforcement in rhesus monkeys
The relative potency of SNC80 to each agonist is also shown (relative potency = SNC80 ED50 ÷ agonist ED50). This relative potency was used to determine proportions of SNC80 and each agonist used in subsequent experiments with drug mixtures.
Effects of SNC80 in Combination with μ Agonists. The effects of each μ agonist administered alone or in combination with SNC80 are shown in Fig. 2 (top). All SNC80 + μ agonist mixtures produced dose-dependent decreases in response rates. Addition of SNC80 produced leftward shifts in the dose-effect curve for morphine, and the magnitude of these left shifts was dependent on the proportion of SNC80 in the mixture. Across the range of proportions tested, the addition of SNC80 produced much smaller changes in the dose-effect curves for the other μ agonists methadone, fentanyl, or nalbuphine. Isobolographic display of ED50 values for SNC80 + μ agonist mixtures indicated that mixtures of SNC80 + morphine decreased response rates with ED50 values that fell close to the line of additivity (Fig. 2, bottom). Statistical analysis confirmed that experimentally determined ED50 values for SNC80 + morphine mixtures (Zmix) were similar to predicted ED50 values for these mixtures (Zadd; Table 2). Mixtures of SNC80 + methadone, fentanyl, or nalbuphine tended to decrease response rates with ED50 values that fell above the line of additivity on the isobolograms, suggestive of subadditivity. However, statistical comparisons of experimentally determined ED50 values (Zmix) and predicted additive ED50 values (Zadd) indicated that only the 10:1 mixture of SNC80 + fentanyl was significantly less potent than predicted (i.e., Zmix > Zadd).
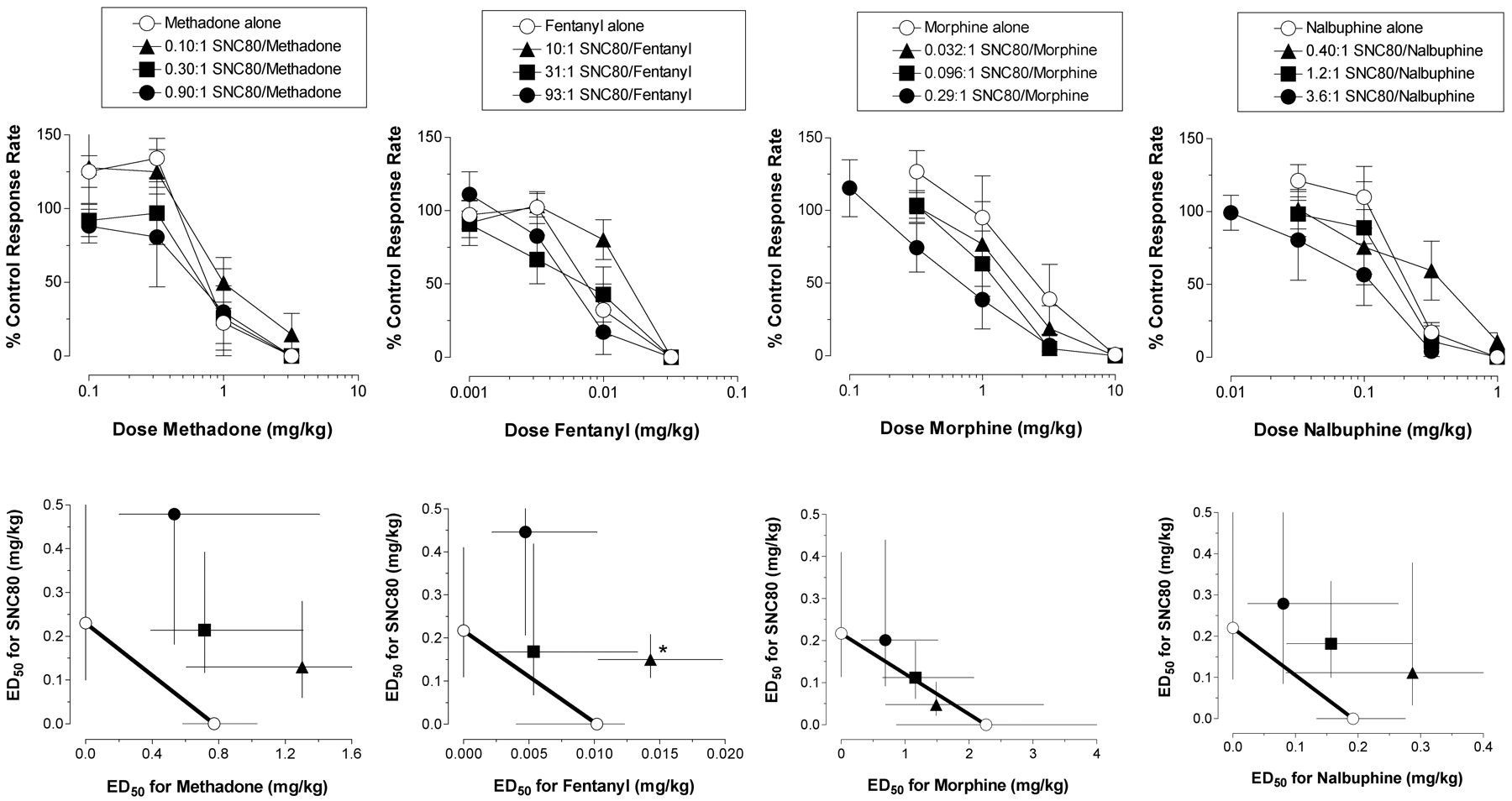
Effects of SNC80 + μ agonist mixtures on schedule-controlled responding. Top, dose-effect curves for each μ agonist alone and in mixtures with SNC80. Abscissae, dose μ agonist in the mixture in milligrams per kilogram (log scale). Ordinates, percentage of control response rate. All points show mean data from four or five monkeys. Open circles show data for the μ agonist alone. Filled symbols show data for SNC80 + μ agonist mixtures, and the proportion of SNC80 in each mixture is shown in the legend boxes at the top of the figure. Bottom, isobolograms for each SNC80 + μ agonist mixture. Abscissae, ED50 value of the μ agonist (milligrams per kilogram). Ordinates, ED50 value of SNC80 (milligrams per kilogram). Each panel shows mean data (95% confidence limits) for four or five monkeys. Open circles show data for SNC80 or the μ agonist alone, and filled symbols show data for SNC80 + μ agonist mixtures. Asterisks indicate experimentally determined values significantly different from additivity (Table 2).
Predicted additive ED50 values (Zadd) and experimentally determined ED50 values (Zmix) of mixtures of SNC80 administered in combination with the μ agonists methadone, fentanyl, morphine, and nalbuphine or the κ agonists U69,593 and bremazocine in an assay of schedule-controlled responding
Zadd and Zmix values show the total dose of SNC80 + agonist in the mixture in milligrams per kilogram (95% CL). The proportion of SNC80 is shown for each mixture.
Effects of SNC80 in Combination with κ Agonists. The effects of the κ agonists U69,593 and bremazocine administered alone or in combination with SNC80 are shown in Fig. 3 (top). All SNC80 + κ agonist mixtures produced dose-dependent decreases in response rates. Moreover, addition of SNC80 produced leftward shifts in the dose-effect curve for both κ agonists, and the magnitude of these left shifts was dependent on the proportion of SNC80 in the mixture. SNC80 + κ agonist mixtures decreased response rates with ED50 values that fell close to the line of additivity on the isobolograms (Fig. 3, bottom). Statistical analysis confirmed that experimentally determined ED50 values (Zmix) and predicted additive ED50 values (Zadd) were similar for all mixtures of SNC80 + U69,593 or bremazocine (Table 2).

Effects of SNC80 + κ agonist mixtures on schedule-controlled responding. Other details as in Fig. 2.
Assay of Thermal Nociception
Baseline T10 Values and Effects of Opioid Agonists Alone. Under baseline conditions, maximum tail-withdrawal latencies (i.e., 20 s) were typically obtained with temperatures of 38 and 42°C. As the temperature was increased to 46 and 50°C, tail-withdrawal latencies for individual monkeys were between 0.32 and 6.05 and 0.30 and 1.62 s, respectively. The average baseline T10 value ± S.E.M. throughout the study was 44.18 ± 0.06°C.
The δ agonist SNC80 was without effect in this assay (i.e., there was no statistically significant change in the ΔT10 across a 100-fold dose range, and an ED5°C could not be determined; Fig. 4). The μ-agonists methadone, fentanyl, morphine, and nalbuphine, and the κ-agonists U69,593 and bremazocine each produced a dose-dependent increase in the ΔT10 (Fig. 4), and the ED5°C value for each compound is shown in Table 3. The potency order, excluding SNC80, was identical to the potency order in the assay of schedule-controlled responding: bremazocine > fentanyl = U69,593 > nalbuphine = methadone > morphine. Given that SNC80 was ineffective in the assay of thermal nociception, the relative potencies from the assay of schedule-controlled responding were used to determine the relative proportions of SNC80 and the μ or κ agonist in the subsequent drug mixtures.
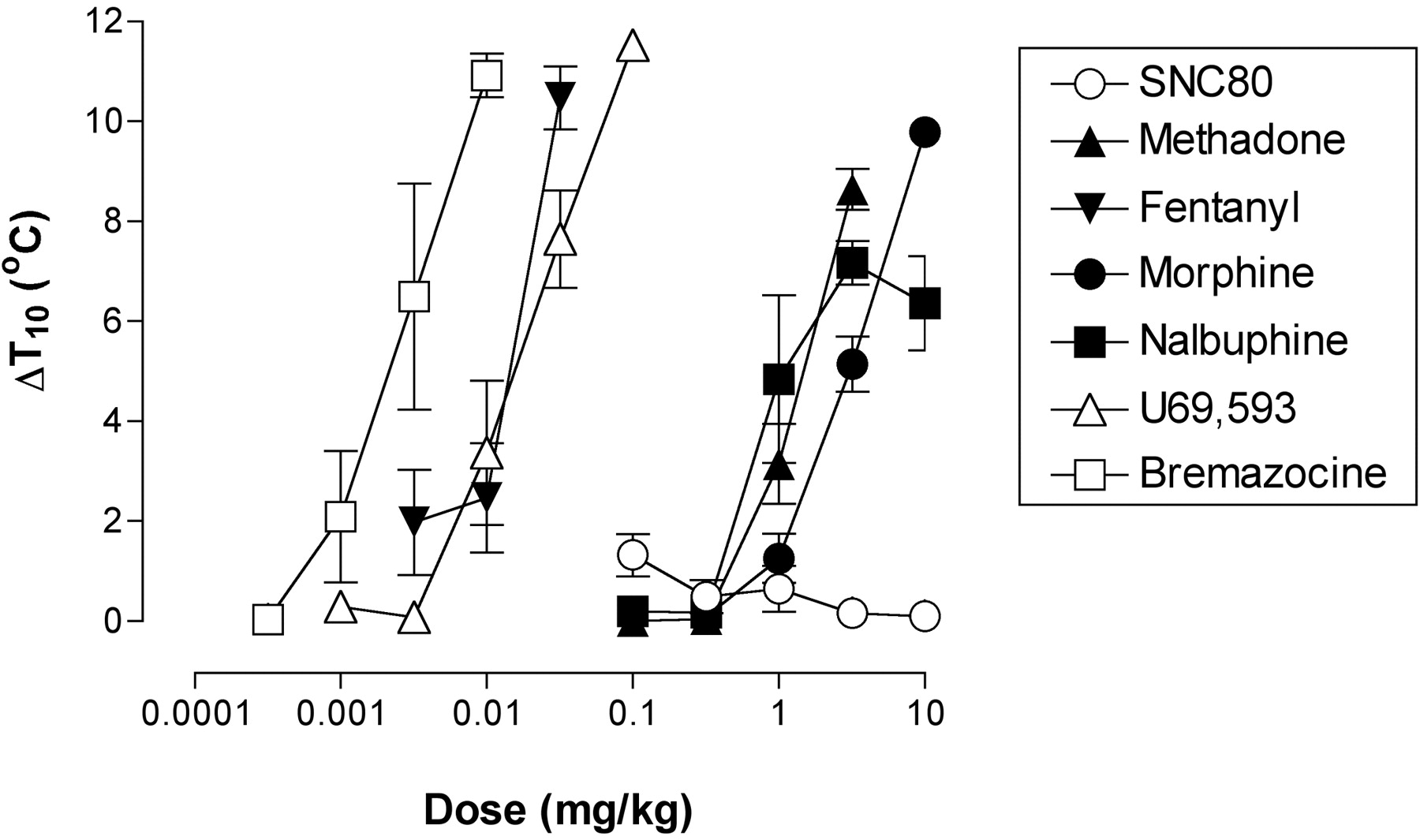
Effects of SNC80, methadone, fentanyl, morphine, nalbuphine, U69,593, and bremazocine administered alone on thermal nociception. Abscissa, dose of opioid agonist in milligrams per kilogram (log scale). Ordinate: Change in threshold for acute thermal pain (Δ T10). All points show mean data from four monkeys. ED5°C values for each drug are shown in Table 3.
ED5°C values in milligrams per kilogram (95% CL) for the μ agonists methadone, fentanyl, morphine, and nalbuphine and the κ agonists U69,593 and bremazocine in an assay of thermal nociception in rhesus monkeys
Note that the δ agonist SNC80 administered alone was ineffective in this procedure.
Effects of SNC80 in Combination with μ Agonists. The effects of each μ agonist administered alone or in combination with SNC80 are shown in Fig. 5 (top). All SNC80 + μ agonist mixtures produced dose-dependent increases in ΔT10 values. Addition of SNC80 produced leftward shifts in the dose-effect curves for methadone, fentanyl, morphine, and nalbuphine, and the magnitude of these left shifts was dependent on the proportion of SNC80 in the mixture. Isobolographic display of ED5°C values for the mixtures indicated that some mixtures of SNC80 + μ agonists increased ΔT10 values with ED5°C values that fell inside the line of additivity (Fig. 5, bottom), suggestive of superadditivity. Statistical analysis confirmed that experimentally determined ED5°C values (Zmix) were significantly lower than predicted additive ED5°C values (Zadd) for each μ agonist in combination with the highest proportion of SNC80 (Table 4). In addition, significant superadditive effects were also observed for methadone, fentanyl, and nalbuphine in combination with the intermediate proportion of SNC80 (Table 4).

Effects of SNC80 + μ agonist mixtures on thermal nociception. Top, dose-effect curves for each μ agonist alone and in mixtures with SNC80. Abscissae, dose μ agonist in the mixture in milligrams per kilogram (log scale). Ordinates, change in threshold for acute thermal pain (Δ T10). All points show mean data from four monkeys. Open circles show data for the μ agonist alone. Filled symbols show data for SNC80 + μ agonist mixtures, and the proportion of SNC80 in each mixture is shown in the legend boxes at the top of the figure. Bottom, isobolograms for each SNC80 + μ agonist mixture. Abscissae, ED5°C value of the μ agonist (milligrams per kilogram). Ordinates, ED5°C value of SNC80 (milligrams per kilogram). Each panel shows mean data (95% confidence limits) for four monkeys. Open circles show data for SNC80 or the μ agonist alone, and filled symbols show data for SNC80 + μ agonist mixtures. Asterisks indicate experimentally determined values significantly different from additivity (Table 4).
Predicted additive ED5°C values (Zadd) and experimentally determined ED5°C values (Zmix) of mixtures of SNC80 administered in combination with the μ agonists methadone, fentanyl, morphine, and nalbuphine or the κ agonists U69,593 and bremazocine in an assay of thermal nociception
Zadd and Zmix values show the total dose of SNC80 + agonist in the mixture in milligrams per kilogram (95% confidence limits). The proportion of SNC80 is shown for each mixture.
Effects of SNC80 in Combination with κ Agonists. The effects of the κ agonists U69,593 and bremazocine administered alone or in combination with SNC80 are shown in Fig. 6 (top). All SNC80 + κ agonist mixtures produced dose-dependent increases in ΔT10 values. The addition of SNC80 did not alter the antinociceptive effects of U69,593. ED5°C values for SNC80 + U69,593 mixtures fell close to the line of additivity on the isobologram (Fig. 6, bottom left), suggestive of additivity. Statistical analysis confirmed that experimentally determined ED5°C values (Zmix) and predicted additive ED5°C values (Zadd) were similar (Table 4). Addition of SNC80 produced leftward shifts in the dose-effect curve for bremazocine, and the magnitude of these left shifts was dependent on the proportion of SNC80 in the mixture. ED5°C values for some SNC80 + bremazocine mixtures fell inside the line of additivity on the isobologram (Fig. 6, bottom right), suggestive of superadditivity. Statistical analysis confirmed that experimentally determined ED5°C values (Zmix) were significantly lower than predicted additive ED5°C values (Zadd) for bremazocine in combination with intermediate and high proportions of SNC80 (Table 4).

Effects of SNC80 + κ agonist mixtures on thermal nociception. Other details as in Fig. 5.
Antagonist Effects of Naltrindole. As noted above, methadone alone produced a dose-dependent increase in ΔT10 values, and addition of SNC80 produced a leftward shift in the methadone dose-effect curve. Figure 7 shows that pretreatment with the δ-selective antagonist naltrindole (1.0 mg/kg) had no effect on the dose-effect curve for methadone alone. However, naltrindole completely blocked the SNC80-induced leftward shift in the methadone dose-effect curve. Thus, after naltrindole pretreatment, the dose-effect curve for the 0.90:1 SNC80 + methadone mixture was similar to the dose-effect curve for methadone alone.

Effects of 1 mg/kg naltrindole pretreatment on 0.9:1 SNC80/methadone mixture and methadone alone. Other details as in Fig. 5, top panels.
Discussion
A major finding of the present study was that SNC80 + μ agonist mixtures produced superadditive effects in the assay of thermal nociception, but only additive or subadditive effects in the assay of schedule-controlled responding. Thus, the nature of the interaction between δ and μ agonists in rhesus monkeys varied as a function of the experimental endpoint. One implication of this finding is that the pharmacological profile of δ + μ agonist mixtures may be different from that of the component drugs. For example, SNC80 was more effective and methadone was more potent in producing rate-decreasing effects than thermal antinociception. However, SNC80 + methadone mixtures were more potent in producing antinociception than rate-decreasing effects. These findings suggest that it may be possible to manipulate opioid agonist mixtures to develop safer and more effective medications that selectively enhance clinically useful effects of the component drugs (e.g., analgesia), while having less effect on or attenuating some undesirable effects (e.g., sedation). In contrast to the results with combinations of δ and μ agonists, combinations of SNC80 and the κ agonist U69,593 produced only additive antinociceptive and rate-decreasing effects. This latter finding demonstrates a degree of pharmacological selectivity in opioid interactions in rhesus monkeys.
δ/μ Interactions and Antinociception. In the present study, some mixtures of SNC80 + μ agonists produced superadditive antinociceptive effects. Antinociceptive interactions were influenced by the relative proportions of the δ and μ agonists, and superadditive effects were obtained only with mixtures that contained an intermediate or high proportion of SNC80. To assess the role of μ agonist efficacy, the effects of SNC80 were examined in combination with four μ agonists that differed in their relative efficacy at μ opioid receptors. SNC80 produced superadditive antinociceptive effects in combination with all four μ agonists. Thus, the present results suggest that, in rhesus monkeys, μ agonist efficacy may not be a critical determinant of the interaction between the antinociceptive effects of δ and μ agonists.
These results in an assay of thermal nociception in rhesus monkeys agree with previous results obtained in an assay of shock titration in squirrel monkeys (Dykstra et al., 1993, 2002). Both SNC80 and BW383U86 produced weak and inconsistent antinociceptive effects when administered alone in this shock titration procedure. However, SNC80 and BW373U86 enhanced the effects of a range of μ agonists as indicated by leftward or upward shifts in μ agonist dose-effect curves. Two factors were cited as potentially important determinants of δ/μ interactions (Dykstra et al., 1993, 2002). First, studies with SNC80 indicated that δ/μ interactions were most evident when SNC80 and the μ agonist were given simultaneously. SNC80 was less effective in enhancing μ agonist effects when it was administered as a pretreatment 20 min before determination of a cumulative μ agonist dose-effect curve. In the present study, SNC80 and μ agonists were administered simultaneously in drug mixtures, and as a result, this approach might be expected to provide the optimal conditions for observing δ/μ interactions. Second, results from studies in the shock titration procedure were interpreted to suggest that δ agonists were more effective in enhancing the antinociceptive effects of intermediate to high-efficacy μ agonists and less effective in combination with low-efficacy μ agonists. In particular, BW373U86 and SNC80 were relatively ineffective in enhancing the effects of the low-efficacy opioid butorphanol. Butorphanol was not examined in the present study, but in agreement with the present results, δ agonists enhanced the effects of methadone, fentanyl, morphine, buprenorphine, and nalbuphine in squirrel monkeys (Dykstra et al., 1993, 2002). Thus, in both squirrel and rhesus monkeys, δ agonists enhanced the antinociceptive effects of μ agonists with a wide range of efficacies at μ receptors.
The ability of δ agonists to enhance the antinociceptive effects of μ agonists under some conditions was first described in studies with rodents (Heyman et al., 1989; Jiang et al., 1990; Porreca et al., 1990; Horan et al., 1992; Adams et al., 1993; O'Neill et al., 1997). For example, combinations of the δ agonists DPDPE or deltorphin with morphine produced superadditive effects in a warm-water tail-flick test in mice (Horan et al., 1992). However, in contrast to results in nonhuman primates, δ opioids modulated the antinociceptive effects of a limited range of μ agonists in rodents. In the warm water tail-withdrawal test in mice, for example, DPDPE enhanced the effects of morphine, normorphine, and codeine but not [d-Ala2,N-Me-Phe4,Gly5-ol]-enkephalin, PLO17, β-endorphin, phenazocine, etorphine, or sufentanil (O'Neill et al., 1997). Moreover, in a warm water tail-withdrawal assay in rats, DPDPE produced only additive or subadditive effects in combination with morphine (Adams et al., 1993). The reason for these discrepancies is not known. Together, these results suggest that there may be important species differences in δ/μ antinociceptive interactions. In particular, δ/μ antinociceptive interactions may be more pronounced in nonhuman primates than in rodents.
The present experiment demonstrated that although the δ antagonist naltrindole had no effect on methadone-induced antinociception alone, it completely blocked the SNC80-induced leftward shift in the methadone dose-effect curve. This suggests that the SNC80-induced increase in methadone antinociceptive potency was δ receptor-mediated. This result is consistent with other reports in monkeys and rodents in which δ agonist-induced increases in μ agonist antinociceptive potency were blocked by δ antagonists (Heyman et al., 1989; Jiang et al., 1990; Adams et al., 1993; Dykstra et al., 1993).
δ/μ Interactions on Other Experimental Endpoints. In contrast to the results in the assay of thermal nociception, the rate-decreasing effects of δ + μ agonist mixtures were only additive or subadditive. These findings confirm and extend previous studies of δ/μ interactions in the assay of shock titration in squirrel monkeys (Dykstra et al., 1993, 2002). In those studies, drugs administered alone or in combination were not tested up to doses that decreased response rates, and a rigorous analysis of drug interactions on rates of responding was not possible; however, there was no evidence to suggest superadditive effects of δ and μ agonists on rates of responding. These results imply that it may be possible to design δ + μ mixtures that serve as strong analgesics with a more favorable profile of undesirable effects than highly selective μ agonists administered alone.
δ/μ interactions have also been examined in a relatively limited set of nonantinociceptive procedures in rodents. In contrast to the synergy often observed in assays of antinociception, δ and μ agonists usually either did not alter or attenuated each other's nonantinociceptive effects (O'Neill et al., 1997; Su et al., 1998). To date, the only endpoint that has shown δ/μ synergism equivalent to that reported for antinociceptive effects has been an inhibition of bladder motility in mice (Sheldon et al., 1989).
δ/κ Interactions. This is the first study to examine δ/κ interactions in nonhuman primates. Mixtures of SNC80 and the highly selective κ agonist U69,593 produced only additive antinociceptive and rate-decreasing effects. This suggests that there is some degree of pharmacological selectivity in opioid agonist interactions in rhesus monkeys. In contrast to the present results, a mixture of DPDPE and the highly selective κ agonist U50,488H produced superadditive effects in an assay of mechanical antinociception in rats (Miaskowski et al., 1990). Furthermore, gestational and hormone-simulated pregnancy antinociception in rats has been shown to require concomitant activation of spinal δ and κ opioid systems, which has been interpreted to reflect a synergistic interaction between δ and κ opioid systems in rodents (Dawson-Basoa and Gintzler, 1998). Further studies will be required to examine the determinants of δ/κ interactions.
Results with the nonselective κ-agonist bremazocine were intermediate between results of SNC80 with selective μ and selective κ agonists. As with mixtures of SNC80 + μ agonists, mixtures of SNC80 + bremazocine produced additive rate-decreasing effects and superadditive antinociceptive effects. However, in contrast to the effects obtained with μ agonists, 1) the effects of SNC80 on bremazocine antinociception were not monotonically related to the proportion of SNC80; 2) the effects of SNC80 on bremazocine antinociception were smaller than its effects on μ-agonist antinociception; and 3) despite statistically significant superadditive effects, the 95% confidence limits of Zadd and Zmix overlapped for all SNC80 + bremazocine combinations. The behavioral profile of SNC80 + bremazocine mixtures may be the result of bremazocine's mixed action at μ and κ receptors. For example, the superadditive antinociceptive effects of the combination may be due to weak bremazocine activity at the μ receptor (Negus et al., 1991) and SNC80 enhancement of those μ agonist effects. Alternatively, bremazocine and U69,593 may act at different κ receptor subtypes in rhesus monkeys (Butelman et al., 1993; Ko et al., 1998), and δ receptors may interact differentially with these κ receptor subtypes.
Mechanisms of Opioid Interactions. The neural mechanisms that underlie antinociceptive interactions between δ and μ agonists in rhesus monkeys are not known. These behavioral interactions could reflect drug interactions at physically coupled δ and μ receptors, intracellular interactions mediated by separate δ and μ receptors located on common neurons, or intercellular interactions mediated by δ and μ receptors located on separate neurons in a common neural circuit (Mansour et al., 1988; Mansour et al., 1995; Cheng et al., 1997; Jordan et al., 2000). The behavioral selectivity of δ/μ interactions suggests that there is some neuroanatomical selectivity in the substrate that mediates those interactions. The results of the present study, for example, suggest that substrates capable of mediating synergistic δ/μ interactions may exist in neural systems underlying opioid-induced antinociception, but not in neural systems underlying opioid-induced decreases in operant responding. Regarding the possibility of physically coupled δ and μ receptors, complexes composed of δ and μ receptors were first proposed on the basis of behavioral studies (Vaught and Takemori, 1979) and evidence from receptor binding assays (Rothman and Westfall, 1981; Rothman and Westfall, 1982). More recently, heterodimers composed of δ and μ receptors have been demonstrated with immunoprecipitation techniques in vitro, and these dimers seem to mediate both cooperative binding of δ and μ ligands and synergistic δ/μ interactions in functional assays (Gomes et al., 2000; Jordan et al., 2000). However, the pharmacology of δ/μ interactions mediated by δ/μ dimers in vitro does not correspond precisely with δ/μ antinociceptive interactions observed in the present study. For example, synergistic δ/μ interactions in vitro could be obtained with some δ antagonists, but not with SNC80 (Gomes et al., 2000). Conversely, the present study found δ/μ antinociceptive interactions with SNC80, but not with the δ antagonist naltrindole. It should be pointed out that δ/κ dimers have also been identified in vitro, and these dimers mediated cooperative binding and synergistic functional effects between the δ agonist DPDPE and the κ agonist U69,593 (Jordan and Devi, 1999). However, in the present study, SNC80 did not produce synergistic behavioral effects in combination with U69,593. Together, these results suggest that opioid receptor dimers identified to date from in vitro studies may not contribute to opioid antinociceptive interactions in rhesus monkeys.
Acknowledgments
We thank Dr. Sharron Kirchain for expert veterinary assistance.
Footnotes
-
This work was supported by Grants R01-DA11460, T32-DA07252, and P01-DA14528 from the National Institute on Drug Abuse, National Institutes of Health.
-
DOI: 10.1124/jpet.103.056515.
-
ABBREVIATIONS: DPDPE, [d-Pen2,d-Pen5]-enkephalin; SNC80, (+)-4-[(αR)-α-((2S,5R)-4-allyl-2,5-dimethyl-1-piperazinyl)-3-methoxy-benzyl]-N,N-diethyl-benzamide; BW373U86, (±)-4-[(R*)-[(2S*,5R*)-2,5-dimethyl-4-(2-propenyl)-1-piperazinyl]-(3-hydroxyphenyl)methyl]-N,N-diethylbenzamide hydrochloride; PL017, [N-MePhe3,d-Pro4]morphiceptin.
- Received July 3, 2003.
- Accepted August 29, 2003.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}