


Abstract
The selective dopamine D3-receptor antagonist S33084 dose dependently attenuated induction of hypothermia by 7-hydroxy-2-dipropylaminotetralin (7-OH-DPAT) and PD128,907. S33084 also dose dependently reduced 7-OH-DPAT-induced penile erections (PEs) but had little effect on 7-OH-DPAT-induced yawning and hypophagia, and it did not block contralateral rotation elicited by the preferential D3 agonist quinpirole in unilateral substantia nigra-lesioned rats. In models of potential antipsychotic activity, S33084 had little effect on conditioned avoidance behavior and the locomotor response to amphetamine and cocaine in rats, and weakly inhibited apomorphine-induced climbing in mice. Moreover, S33084 was inactive in models of potential extrapyramidal activity in rats: induction of catalepsy and prolactin secretion and inhibition of methylphenidate-induced gnawing. Another selective D3antagonist, GR218,231, mimicked S33084 in inhibiting 7-OH-DPAT-induced PEs and hypothermia but neither hypophagia nor yawning behavior. Similarly, it was inactive in models of potential antipsychotic and extrapyramidal activity. In distinction to S33084 and GR218,231, the preferential D2 antagonist L741,626 inhibited all responses elicited by 7-OH-DPAT. Furthermore, it displayed robust activity in models of antipsychotic and, at slightly higher doses, extrapyramidal activity. In summary, S33084 was inactive in models of potential antipsychotic and extrapyramidal activity and failed to modify spontaneous locomotor behavior. Furthermore, it did not affect hypophagia or yawns, but attenuated hypothermia and PEs, elicited by 7-OH-DPAT. This profile was shared by GR218,231, whereas L741,626 was effective in all models. Thus, D2-receptors are principally involved in these paradigms, although D3-receptors may contribute to induction of hypothermia and PEs. S33084 should comprise a useful tool for further exploration of the pathophysiological significance of D3- versus D2-receptors.
InMillan et al. (2000b), we describe a novel benzopyranopyrrole, S33084, that behaves as a potent, competitive, and selective (>100-fold) antagonist at cloned, human (h)D3- versus hD2-receptors (Table1), and which shows only negligible (>200-fold) lower affinity for all other sites (>30) examined to date. S33084 also interacts selectively with cloned and native rat D3- versus D2-receptors (Cussac et al., 2000). Thus, S33084 provides, a potentially important tool for exploration of the functional significance of D3- compared with D2-receptors. Indeed, the use of S33084, in parallel with a further selective D3 antagonist GR218,231 (Murray et al., 1996) and the preferential D2 antagonist L741,626 (Kulagowski et al., 1996) indicated that D2- compared with D3-receptors are predominantly implicated in the tonic modulation of ascending dopaminergic transmission in rats. However, D3-receptors may contribute to the phasic control of dopamine (DA) release in frontal cortex (Millan et al., 2000b).
Summary of the interaction of S33084 compared with GR218,231 and L741,626, at D3- versus D2 receptors
In this study, we addressed two complementary and interdependent issues. First, we characterized the in vivo actions of S33084 in several models involving dopaminergic activity in rodents, in particular paradigms considered predictive of antipsychotic or extrapyramidal properties. Second, using S33084 together with GR218,231 and L741,626, we examined the role of D3- compared with D2-receptors in the diverse functional models used. These may be summarized as follows. First, we examined the decrease in core temperature (CT) elicited by the agonists PD128,907 and 7-hydroxy-2-dipropylaminotetralin (7-OH-DPAT). Although considered “preferential” D3- versus D2-receptor ligands, PD128,907 and 7-OH-DPAT may elicit actions via the D2 and/or D3 sites in vivo (see Discussion;Pugsley et al., 1995; Bristow et al., 1996; Shafer and Levant, 1998). Second, we evaluated several other dopaminergic behaviors elicited by 7-OH-DPAT that are assumed to be mediated by D2-receptors, i.e., yawning, penile erections (PEs), and hypophagia in rats (Leibowitz, 1980; Melis et al., 1987;Ladurelle et al., 1991; Argiolas and Melis, 1997). Third, in view of the possibility that D3-receptors may be implicated in psychotic states and their treatment (Shafer and Levant, 1998; Harrison, 1999; Kerwin and Owen, 1999), we examined several models of potential antipsychotic activity. These included “classic” procedures in which dopaminergic mechanisms are strongly implicated: induction of hyperlocomotion in rats by the psychostimulants amphetamine and cocaine, conditioned avoidance responses (CARs) in rats, and apomorphine-induced climbing behavior in mice (Millan et al., 1998a,b; Moore, 1999). In addition, inasmuch as glutamatergic deficits may contribute to the etiology of psychotic states (Tamminga, 1998), we also examined the hyperlocomotion response to the open channel blocker atN-methyl-d-aspartate (NMDA) receptors, phencyclidine (PCP) (Gorelick and Balster, 1995; Millan et al., 1999). This response is of particular note because serotonergic, as well as dopaminergic, mechanisms are implicated in the actions of PCP (Svensson et al., 1995; Martin et al., 1997; Millan et al., 1999; Moore, 1999). Fourth, D3-receptors, which are preferentially localized in limbic structures (Hall et al., 1996; Shafer and Levant, 1998; Suzuki et al., 1998; Gurevich and Joyce, 1999), may play a role different to that of D2 sites in the control of motor behavior (Baik et al., 1995; Millan et al., 1995; Kelly et al., 1998; Shafer and Levant, 1998). Thus, we likewise investigated models involving interruption of dopaminergic transmission in the striatum and predictive of extrapyramidal motor side effects: induction of catalepsy and blockade of the stereotyped gnawing evoked by methylphenidate in rats (Cunningham-Owens, 1996; Keks, 1996; Millan et al., 1998b). In complementary studies, we examined both spontaneous locomotor activity and contralateral rotation elicited by the dopaminergic D2/D3 agonist quinpirole in rats sustaining a unilateral lesion of the substantia nigra, pars compacta (SNPC; Millan et al., 1998a,b). Finally, as a measure of potential endocrine, extrapyramidal side effects (Cunningham-Owens, 1996; Keks, 1996), circulating levels of prolactin (PRL) were examined.
Materials and Methods
Animals.
In vivo studies used male Wistar rats weighing 220 to 250 g, unless otherwise specified (Iffa-Credo, L'Arbresle, France) and male CD1 mice weighing 22 to 25 g (Charles River, Saint-Aubin-les-Elbeuf, France). They were maintained in sawdust-lined cages with unrestricted access to food and water. The laboratory temperature was held at 21 ± 1°C and humidity was controlled at 60 ± 5%. There was a 12-h light/dark cycle, with lights on from 7:30 AM to 9:30 PM. Before experimentation, all animals were adapted for at least 1 week to laboratory conditions.
Induction of Hypothermia by PD128,907 and 7-OH-DPAT.
In lightly restrained rats, CT was determined with a (rectal) digital thermistoprobe as described in Millan et al. (1995). Basal CT was determined; S33084, GR218,231, L741,626, or vehicle was administered; and 30 min later, vehicle, PD128,907 (0.63 mg/kg s.c.), or 7-OH-DPAT (0.16 mg/kg s.c) was injected. Thirty minutes later, CT was redetermined and the difference to pretreatment values calculated. Data were analyzed by ANOVA and ID50 values plus 95% CL were calculated. The percentage of inhibition was computed as follows: 100 × [1 − [(antagonist + agonist) − (vehicle + vehicle)]]/[(vehicle + agonist) − (vehicle + vehicle)].
Induction of PE and Yawning by 7-OH-DPAT.
Rats weighing 120 to 140 g were individually placed in transparent, Plexiglas observation cages (11 × 26 × 30.5 cm). A mirror was situated behind the cage to facilitate observation of behavior. Thirty minutes after S33084, GR218,231, L741,626, or vehicle administration, 7-OH-DPAT (0.04 mg/kg s.c.) or vehicle was administered, and PE and yawns measured over a 30-min period. Data were analyzed by ANOVA followed by Dunnett's test and ID50 values (95% CL) were calculated.
Induction of Hypophagia by 7-OH-DPAT.
Individually housed rats were food deprived for 24 h before the experiment. S33084, GR218,231, or L741,626 followed by 7-OH-DPAT (0.16 mg/kg s.c.) or vehicle was administered at 45 and 30 min, respectively, before reaccess to a preweighed quantity of food (standard chow). One hour later, chow was reweighed and food intake (FI) calculated. Data were analyzed by ANOVA followed by Dunnett's test and ID50 values (95% CL) were calculated. The percentage of inhibition was computed as follows: 100 × [1 − [(antagonist + 7-OH-DPAT) − (vehicle + vehicle)]]/[(vehicle + 7-OH-DPAT) − (vehicle + vehicle)].
Influence on Rotation in Unilateral, SNPC-Lesioned Rats.
As detailed in Millan et al. (1998a,b), rats with a unilateral 6-hydroxydopamine (8 μg/4 μl) lesion of the left SNPC were allowed 3 weeks after surgery to recover before testing. Only animals that showed a pronounced response (>100 contralateral rotations), first to apomorphine (0.04 mg/kg s.c.; two sessions, 1 week apart) and subsequently to quinpirole (0.02 mg/kg, s.c.; two further sessions, 1 week apart), were selected for the study. The number of contralateral rotations effected 0 to 60 min after administration of apomorphine, or 20 to 50 min after administration of quinpirole, were monitored automatically with rats coupled to a harness connected to a Rotacount 8 (Columbus Instruments, Columbus, OH) apparatus. After selection of the animals, sessions were performed once a week, with an ABACADA design, with A corresponding to control and B, C, and D to test sessions. Rats were administered with vehicle (control session) or S33084, GR218,231, or L741,626 (test session) 25 min before quinpirole. Drug actions were expressed as a function of the mean of the preceding and subsequent control sessions (defined as 100%). The influence of S33084, GR218,231, and L741,626 alone was evaluated in test sessions where rats were first administered with vehicle and, 25 min later, with the above-mentioned compounds instead of quinpirole. Data were analyzed by paired Student's t test and ID50values (95% CL) were calculated for antagonism of quinpirole.
Conditioned Avoidance Procedure in Rats.
As detailed inMillan et al. (1998a), rats were trained to avoid an electric shock (560 μA, 5-s maximal duration) by switching compartments of a shuttle-box (Letica, Barcelona, Spain) on the appearance of a light. Each trial (10 daily) consisted of a 10-s period with the light on, followed or not by a period with shock, dependent on the response to the light. The trial terminated once the rat had moved into the other compartment, either during the light-on period (CAR), or during the shock period (escape response). The maximal duration of the trial was 15 s (if the animal did not respond to the shock: escape failure or nonresponse). The number of CARs (maximum of 10) was determined. S33084, GR218,231, L741,626, or vehicle was administered 30 min before testing. Data were analyzed by a paired Wilcoxon signed rank test and ID50 values (95% CL) were calculated.
Apomorphine-Induced Climbing in Mice.
The induction of climbing behavior was examined in mice individually placed in steel cylinders (14 cm in diameter) possessing walls (14 cm in height) constructed of vertical bars (1 cm apart and 2 mm in diameter). Climbing behavior was assessed as previously (Millan et al., 1998a) according to a rating scale (0–4). The mean of two measures made 10 and 20 min following apomorphine (0.75 mg/kg s.c.) was determined. S33084, GR218,231, L741,626, or vehicle was administered s.c. 30 min before apomorphine. Data were analyzed by ANOVA followed by Dunnett's test and ID50 values (95% CL) were calculated.
Spontaneous, Cocaine-, Amphetamine-, PCP-, and Dizocilpine-Induced Locomotion in Rats.
As described in Millan et al. (1998a), rats were administered with S33084, GR218,231, L741,626, or vehicle and individually placed for a 30-min habituation period in transparent polycarbonate cages (45 × 30 × 20 cm) located in activity chambers (Lablinc System; Coulbourn, Lehigh Valley, PA). Then, the animals received vehicle (spontaneous locomotion), cocaine (20 mg/kg i.p.), amphetamine (2.5 mg/kg i.p.), PCP (20.0 mg/kg s.c.), or dizocilpine (0.16 mg/kg s.c.) and locomotion was monitored for 60 min. A movement corresponded to the consecutive interruption within 2 s of two infrared beams situated 24 cm apart and 4 cm above the cage floor. Data were analyzed by ANOVA followed by Dunnett's test and ID50 values (95% CL) were calculated.
PRL Levels in Rats.
PRL levels were determined essentially as described in Millan et al. (1998a), in systemic plasma 30 min after application of S33084, GR218,231, L741,626, or vehicle. Levels of PRL were determined by radioimmunoassay with a highly selective antibody directed against rat PRL, which displayed <0.1% cross-reactivity to all other hormones tested (RPQ553; Amersham, Buckingham, England). Data were analyzed by ANOVA followed by Dunnett's test.
Catalepsy in Rats.
As detailed in Millan et al. (1998a), the left and right hindpaws of rats were placed over the corresponding forepaw and the duration of this position was determined, with a maximum possible duration of 30 s. Three independent measures were made, separated by 1-min intervals. S33084, GR218,231, L741,626, or vehicle was administered 30 min before testing. The mean of the three determinations was calculated. Data were analyzed by ANOVA followed by Dunnett's test. Active dose (AD)50 values (95% CL) were calculated.
Methylphenidate-Induced Gnawing in Rats.
As described inMillan et al. (1998a), rats were administered with methylphenidate (40.0 mg/kg i.p.) or vehicle and 30 min later, the number of gnawing periods recorded over 10 min (one 10-s observation period per minute) in observation cages (33.5 × 23.5 × 19 cm). S33084, GR218,231, L741,626, or vehicle was injected 30 min before methylphenidate or vehicle. Data were analyzed by ANOVA followed by Dunnett's test and ID50 values (95% CL) were calculated.
Spontaneous Locomotion in Mice Not Habituated to Observation Chamber.
As described in Millan et al. (1998b), 30 min after S33084, GR218,231, L741,626, or vehicle administration, mice were placed for 10 min in individual, white Plexiglas chambers (27 × 27 × 27 cm) furnished with two rows of four photocells, located 2 cm above the floor and 6 cm apart. These photocells were connected via an interface to a computer with software written by Osys/Orga System (Changé, France). The interruption of two adjacent beams corresponded to a locomotion count. Data were analyzed by ANOVA followed by Dunnett's test and ID50 values (95% CL) were calculated.
Spontaneous Locomotion in Rats Not Habituated to Observation Chamber.
Locomotor behavior was evaluated in rats placed in transparent polycarbonate cages (45 × 30 × 20 cm) located in activity chambers (Lablinc System, Coulbourn). Locomotion was monitored for 12 min. Drugs were given 30 min before testing. A movement corresponds to the consecutive interruption within 2 s of two infrared beams situated 24 cm apart and 4 cm above the cage floor. Data were analyzed by ANOVA followed by Dunnett's test and ID50 values (95% CL) were calculated.
Drug Doses, Solution, Salts, and Sources.
In all procedures, full dose-response relationships were evaluated for S33084, GR218,231, and L741,626. The maximal dose evaluated for L741,626 was 40.0 mg/kg s.c., and as exemplified in Results, fully permitted the expression of its actions in the in vivo protocols examined herein. For S33084 and GR218,231, maximal doses of 10.0 mg/kg were defined. These maximum doses were determined by the absolute limit of their solubility. Furthermore, relative to their very high affinities at D3-receptors (Millan et al., 2000b), they are higher than those required for the demonstration of any potential D3-receptor-mediated actions in vivo (seeDiscussion). All drug doses are in terms of the base. Drugs were dissolved in sterile water. For S33084, GR218,231, and L741,626, a few drops of lactic acid were added and the pH adjusted to as close to neutrality as possible (>5.0). Drugs were injected s.c. in an injection volume of 1 ml/kg (rats) or 10 ml/kg (mice). Drug structures, sources, and salts were as follows: d-amphetamine sulfate (Calaire Chimie, Calais, France); cocaine HCl (Coopérative Pharmaceutique Française, Melun, France); PD128,907 {(+)-(4aR,10bR)-3,4,4a,10b-tetrahydro-4-propyl-2H,5H-[1]benzopyrano-[4,3-b]-1,4-oxazin-9-ol) HCl}, quinpirole HCl, dizocilpine maleate, and 7-OH-DPAT HCl (Research Biochemicals International, Natick, MA); apomorphine HCl and phencyclidine HCl (Sigma Chemical Co., St Quentin-Fallavier, France); L741,626 base (Tocris Cookson, Bristol, UK); and methylphenidate HCl (Ciba-Geigy, Basel, Switzerland). S33084 and GR218,231 were synthesized by G. Lavielle and J.-L. Peglion (Institut de Recherches Servier), respectively.
Results
Modulation of PD128,907- and 7-OH-DPAT-Induced Hypothermia (Fig.1).
Both PD128,907 and 7-OH-DPAT elicited a pronounced reduction in CT in rats: vehicle = +0.74 ± 0.04°C versus PD128,907 = −1.44 ± 0.08°C and versus 7-OH-DPAT = −1.49 ± 0.07°C, P< .001 (in unpaired t test) in each case. Their actions were dose dependently and significantly attenuated by S33084 with ID50 values (95% CL) of 3.1 (1.1–9.2) and 8.8 (5.6–13.8) mg/kg s.c., respectively. Alone, administered over an identical dose range, S33084 slightly (and significantly) modified CT only at the highest dose (10.0) tested:F4,26 = 34.3, P < .001; vehicle = +0.73 ± 0.06°C versus S33084 (10.0) = +0.03 ± 0.26°C, P < .01 in Dunnett's test. GR218,231 also dose dependently attenuated the hypothermic actions of PD128,907 and 7-OH-DPAT with ID50 values (95% CL) of 7.3 (2.2–24.3) and 1.7 (0.9–3.3) mg/kg s.c., respectively. Alone, administered over an identical dose range, GR218,231 slightly (and significantly) reduced CT at the highest dose (10.0) tested:F4,24 = 10.2, P < .001; vehicle = +0.67 ± 0.08°C versus GR218,231 (10.0) = +0.28 ± 0.06°C, P < .01 in Dunnett's test. L741,626 similarly dose dependently inhibited the reduction in CT provoked by PD128,907 and 7-OH-DPAT with ID50 values (95% CL) of 3.4 (1.6–7.1) and 3.3 (0.8–13.4) mg/kg s.c., respectively. It slightly and significantly reduced CT itself at high doses: F5,32= 31.1, P < .001; vehicle = +0.81 ± 0.08°C versus L741,626 (10.0) = +0.40 ± 0.12°C and L741,626 (20.0) = +0.10 ± 0.23°C, P < .05 (in Dunnett's test) in each case.

Influence of S33084, GR218,231, and L741,626 on 7-OH-DPAT- and PD128,907-induced hypothermia in rats. Data are means ± S.E (n = 5 per value). ANOVAs for the influence of S33084, GR218,231, and L741,626 versus 7-OH-DPAT and PD128,907 are as follows. 7-OH-DPAT: S33084,F3,28 = 38.5, P < .001; GR218,231, F3,26 = 138.0,P < .001; and L741, 626,F5,34 = 105.9, P < .001. PD128,907: S33084, F4,18 = 138.3,P < .001; GR218,231,F3,18 = 55.5, P < .001; and L741,626, F3,18 = 43.4,P < .001. Asterisks indicate significance of drug/PD128,907 or drug/7-OH-DPAT versus vehicle/PD128,907 or vehicle/7-OH-DPAT values, respectively, in Dunnett's test. *P < .05.
Modulation of Induction by 7-OH-DPAT of PE and Yawning Behavior (Fig. 2).
7-OH-DPAT elicited both PE and yawning behavior. S33084 dose dependently attenuated, although it did not abolish, PE with an ID50 (95% CL) of 2.8 (1.0–7.7) mg/kg s.c., whereas it did not significantly reduce yawning behavior. GR218,231 also significantly attenuated the induction of PE by 7-OH-DPAT with an ID50 (95% CL) of 8.7 (4.0–18.7) mg/kg s.c., whereas it did not significantly modify the yawning response. L741,626 dose dependently reduced the induction of PE by 7-OH-DPAT with an ID50 (95% CL) of 2.8 (1.7–4.8) mg/kg s.c., although this response was not abolished. Similarly, it dose dependently reduced yawning behavior elicited by 7-OH-DPAT with an ID50 (95% CL) of 1.1 (0.5–2.4) mg/kg s.c. In the presence of S33084, GR218,231 and L741,626 alone, PE and yawns were not observed (data not shown).

Influence of S33084, GR218,231, and L741,626 on 7-OH-DPAT-induced PEs and yawning behavior in rats. Data are means ± S.E. (n = 7 per value). ANOVAs for the influence of S33084, GR218,231, and L741,626 are as follows. PE: S33084,F6,59 = 4.6, P < .001; GR218,231, F4,44 = 3.9,P < .01; and L741,626,F6,53 = 7.1, P < .001. Yawns: S33084, F6,59 = 1.8,P > .05; GR218,231,F4,44 = 0.5, P > .05; and L741,626, F6,53 = 13.9,P < .001. Asterisks indicate significance of drug/7-OH-DPAT versus vehicle/7-OH-DPAT values in Dunnett's test. *P < .05.
Modulation of 7-OH-DPAT-Induced Hypophagia (Fig.3).
7-OH-DPAT elicited a marked reduction in FI: vehicle = 7.3 ± 0.2 g versus 7-OH-DPAT = 1.9 ± 0.2 g, P < .05 in unpaired t test. This action was unaffected by S33084 and GR218,231, which did not themselves affect FI (data not shown). In contrast, L741,626 dose dependently blocked the hypophagic action of 7-OH-DPAT (Fig. 3) with an ID50 (95% CL) of 0.7 (0.3–1.6) mg/kg s.c., without itself significantly modifying FI (data not shown).

Influence of S33084, GR218,231, and L741,626 on 7-OH-DPAT-induced hypophagia. Data are means ± S.E. (n = 5 per value). ANOVAs for the influence of S33084, GR218,231, and L741,626 versus 7-OH-DPAT are as follows. S33084, F4,34 = 0.9,P > .05; GR218,231,F4,27 = 0.2, P > .05; and L741,626, F3,26 = 5.8,P < .01. The asterisk indicates significance of drug/7-OH-DPAT versus vehicle/7-OH-DPAT values in Dunnett's test. *P < .05.
Influence on Quinpirole-Induced Rotation in Unilateral SNPC-Lesioned Rats (Fig. 4).
The dopamine D2/D3 agonist quinpirole elicited a marked contralateral rotation in rats sustaining a unilateral lesion of the SNPC: vehicle + vehicle = 58 ± 12 rotations versus vehicle + quinpirole = 373 ± 44 rotations,P < .001 in an unpaired t test. This action of quinpirole was not significantly inhibited by S33084 and GR218,231, but it was dose dependently blocked by L741,626 with an ID50 (95% CL) of 1.0 (0.4–2.3) mg/kg s.c. S33084, GR218,231, and L741,626, administered alone, did not elicit either contralateral (Fig. 4) or ipsilateral (data not shown) rotations.

Influence of S33084, GR218,231, and L741,626 on quinpirole-induced rotation in rats sustaining a unilateral lesion of the SNPC. Data are means ± S.E. (n = 5 per value). Asterisks indicate significance of drug/quinpirole versus vehicle/quinpirole values in paired Student's t test. *P < .05.
Activity in Models of Potential Antipsychotic Activity (Fig.5).
S33084 and GR218,231 weakly but significantly inhibited the induction of climbing behavior in mice by the dopaminergic agonist apomorphine. Their ID50values (95% CL) were 7.5 (4.2–13.5) and 6.4 (4.6–8.8) mg/kg s.c., respectively. However, this response was dose dependently and potently abolished by L741,626 with an ID50 (95% CL) of 0.3 (0.2–0.6) mg/kg s.c. In a CAR procedure in rats, over a higher dose range, L741,626 also showed robust, dose-dependent activity with an ID50 (95% CL) of 6.1 (3.1–12.0) mg/kg s.c. In contrast to L741,626, neither S33084 nor GR218,231 were inactive in this model. At the highest dose tested (20 mg/kg s.c.), L741,626 significantly enhanced escape failures (vehicle = 0 ± 0 versus L741,626 = 5.6 ± 1.4, P < .01), whereas S33084 (10 mg/kg s.c.) and GR218,231 (10 mg/kg s.c.) did not significantly modify this parameter (1.2 ± 0.1 and 0 ± 0, respectively; P > .05). The psychostimulants amphetamine and cocaine elicited pronounced increase in locomotion in rats: vehicle = 51 ± 7 locomotor counts versus amphetamine = 253 ± 30 counts and versus cocaine = 303 ± 46 counts, with P < .001 (in unpairedt test) in each case. Their actions were dose dependently blocked by L741,626 with ID50 values (95% CL) of 1.0 (0.4–2.6) and 0.5 (0.1–3.5) mg/kg s.c., respectively. In contrast, across a broad dose range, S33084 and GR218,231 did not significantly reduce the actions of amphetamine or cocaine. Finally, the locomotor response to PCP (vehicle = 51 ± 7 counts versus PCP = 230 ± 35 counts, P < .001 in unpaired t test) was not significantly affected by S33084, GR218,231, or L741,626 (not shown).
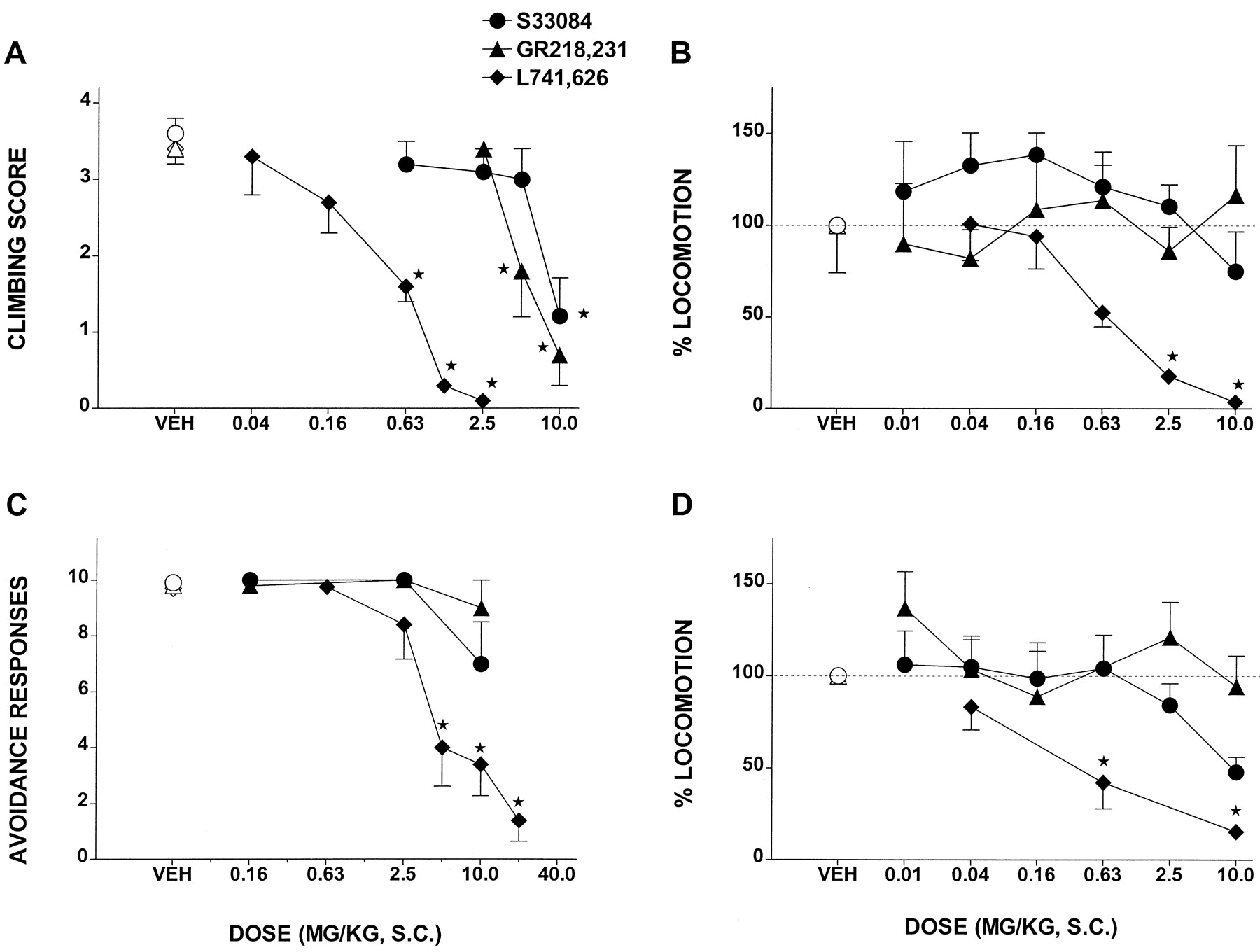
Activity of S33084, GR218,231, and L741,626 in models of potential antipsychotic activity. A, inhibition of apomorphine (0.75 mg/kg s.c.)-induced climbing in mice. B, inhibition of amphetamine (2.5 mg/kg i.p.)-induced locomotion in rats. C, inhibition of conditioned avoidance responses in rats. D, inhibition of cocaine (20.0 mg/kg i.p.)-induced locomotion in rats. Data are means ± S.E. (n = 5 per value). ANOVAs are as follows. A, S33084, F4,34 = 8.6,P < .001; GR218,231,F3,34 = 17.1, P < .001; and L741,626, F5,41 = 31.7,P < .001. B, S33084,F6,43 = 0.8, P > .001; GR218,231, F6,41 = 0.2,P > .05; and L741,626,F5,30 = 8.6, P < .001. D, S 33084, F6,34 = 0.8,P > .05; GR218,231,F5,32 = 0.5, P > .05; and L741,626, F3,17 = 6.9,P < .01. Asterisks indicate significance of differences to vehicle values in Dunnett's test. *P < .05. In C, asterisks indicate significance of differences to vehicle values in the Wilcoxon signed rank test. *P < .05.
Activity in Models of Potential Extrapyramidal Activity (Fig.6).
Although S33084 and GR218,231 did not elicit catalepsy, L741,626 dose dependently elicited a marked and robust catalepsy response activity with an AD50 (95% CL) of 7.0 (4.5–10.9) mg/kg s.c. S33084 and GR218,231 also failed to significantly inhibit the induction of stereotyped gnawing by methylphenidate. In contrast, L741,626 dose dependently blocked this behavior with an ID50 (95% CL) of 2.4 (0.9–6.7) mg/kg s.c. Administered alone (0.16–10.0 mg/kg s.c.), S33084, GR218,231, and L741,626 all failed to elicit gnawing or other stereotyped behaviors such as rearing and sniffing (data not shown). Circulating levels of PRL were not significantly increased by S33084 and GR218,231, whereas they were dose dependently elevated by L741,626.

Influence of S33084, GR218,231, and L741,626 in models predictive of extrapyramidal motor and endocrine side effects. A, induction of catalepsy. B, inhibition of stereotyped gnawing elicited by the DA releaser methylphenidate (40.0 mg/kg i.p.). C, elevation in circulating levels of PRL. Data are means ± S.E. (n = 5 per value). ANOVAs are as follows. A, S33084, F2,15 = 3.2,P > .05; GR218,231,F2,10 = 2.5, P > .05; and L741,626, F3,15 = 54.6,P < .001. B, S33084,F2,11 = 0.1, P > .05; GR218,231, F2,11 = 0.1,P > .05; and L741,626,F3,14 = 17.3, P < .001. C, S33084, F6,28 =1.3,P > .05; GR218,231,F5,24 = 1.1, P > .05; and L741,626, F5,24 = 12.0,P < .001. Asterisks indicate significance of differences to vehicle values in Dunnett's test. *P < .05.
Influence on Spontaneous Locomotor Activity (Fig.7).
In rats habituated to observation chambers for 30 min, i.e., under conditions in which the hyperlocomotion elicited by amphetamine and other psychostimulants was examined (Fig. 5), S33084 and GR218,231 elicited a mild but significant reduction in locomotor activity with ID50 values (95% CL) of 1.6 (0.3–7.4) and 4.9 (1.5–15.9) mg/kg s.c., respectively. L741,626 also dose dependently (0.16–20.0 mg/kg) reduced locomotor activity with an ID50 (95% CL) of 8.1 (3.5–18.9) mg/kg s.c. On reanalysis of these data over the six independent 10-min observation periods, a similar pattern of data was obtained (data not shown). In rats that had not previously been exposed to the observation chambers, likewise over a broad range of doses, S33084 and GR218,231 did not significantly enhance locomotor activity. Furthermore, at the highest doses examined, they significantly diminished locomotor activity [ID50 values (95% CL) of 2.6 (0.5–14.6) and 1.5 (0.3–8.6) mg/kg s.c., respectively]. Under these conditions, L741,626 also dose dependently reduced locomotor activity with an ID50 (95% CL) of 2.3 (0.6–8.3) mg/kg s.c. On reanalysis of these data for the four independent 3-min observation sessions, a similar pattern of data was acquired (data not shown). In mice that had not been habituated to observation chambers, S33084 and GR218,231 did not significantly modify locomotor activity over a broad range of doses. L741,626 (0.63–20.0 mg/kg), however, dose dependently diminished locomotor activity with an ID50 (95% CL) of 5.6 (2.1–15.2) mg/kg s.c.

Influence of S33084, GR218,231, and L741,626 on spontaneous motor behavior in mice and rats. Top, spontaneous locomotor activity in rats habituated to observation chambers. Middle, spontaneous locomotor activity in rats nonhabituated to observation chambers. Bottom, spontaneous locomotor activity in mice nonhabituated to observation chambers. Data are means ± S.E. (n = 5 per value). ANOVAs are as follows. Top, S33084, F7,41 = 5.7,P < .05; GR218,231,F6,43 = 2.5, P < .05; and L741,626, F6,39 = 9.6,P < .001. Middle, S33084,F7,47 = 3.5, P < .01; GR218,231, F6,25 = 11.4,P < .001; and L741,626,F5,23 = 15.4, P < .001. Bottom, S33084, F7,48 = 3.5,P < .05; GR218,231,F5,52 = 2.1, P > .05; and L741,626, F7,40 = 9.3,P < .001. Asterisks indicate significance of differences to vehicle values in Dunnett's test. *P < .05.
Discussion
Active Doses of S33084.
S33084 and GR218,231 show 100-fold higher affinity at D3- versus D2 sites, and their affinity for the latter is ∼10-fold lower than that of L741,626 (Table 1). In the absence of in vivo models attributable exclusively to D3-receptors, these relative potencies permit estimation of doses at which D3- versus D2-receptor-mediated actions of S33084 should be expressed, an approach previously exploited for selective D4-receptor antagonists, for which functional models in vivo are similarly unavailable (Bristow et al., 1997; Millan et al., 1998a). Thus, in models involving D2-receptors, one would expect residual activity of S33084 and GR218,231 to be exerted at doses ∼10-fold higher than those of L741,626. Compared with such D2-receptor-mediated responses, models involving only D3-receptors should reveal actions of S33084 and GR218,231 at 100-fold lower doses. Furthermore, in such models, L741,626 should be effective only at 100-fold higher doses than those of S33084 and GR218,231. Although such calculations are approximate, they provide an instructive framework for cautious interpretation of functional actions of S33084 in vivo, the effects of which should not be automatically attributed to D3-receptors.
Hypothermia.
S33084 attenuated induction of hypothermia by PD128,907 and 7-OH-DPAT, an action mimicked by GR218,231. These observations are indicative of a role of D3-receptors, in particular because the hypothermic potency of D3/D2 agonists correlates with affinities at hD3-receptors (Millan et al., 1995; Varty and Higgins, 1998), whereas a further D3-receptor antagonist, S14297, stereospecifically blocks hypothermia (Millan et al., 1995; Audinot et al., 1998). Furthermore, microinjection studies (Barik and De Beaurepaire, 1998) suggest that the Isles of Calleja, a region enriched in D3-receptors (Shafer and Levant, 1998), mediate 7-OH-DPAT hypothermia. However, as outlined above, active doses of S33084 and GR218,231 are too high to be accounted for by an exclusive role of D3-receptors. Furthermore, L741,626 inhibited hypothermia at doses similar to that of S33084 and GR218,231. This implicates a role of D2-receptors, in line with findings that transgenic mice lacking D2- but not D3-receptors display an attenuation of PD128,907-induced hypothermia (Boulay et al., 1999a,b; Xu et al., 1999). Nevertheless, the possibility that S33084 and GR218,231 interfere with hypothermia via blockade of D2-receptors is unlikely because 1) doses of S33084 and GR218,231 ∼10- to 20-fold higher than that of L741,626 should have been required, and 2) at doses blocking hypothermia, S33084 and GR218,231 were ineffective in other neurochemical (Millan et al., 2000b) and behavioral paradigms (see Results) reflecting D2-receptor blockade.
Thus, in analogy to the ability of S33084 and GR218,231 to prevent suppression of mesocortical DA release by PD128,907 (Millan et al., 2000b), doses blocking hypothermia are intermediate between those considered selective for D3-receptors and those reflecting residual activity at D2 sites. Recent studies of dopaminergic and other classes of receptor suggest several possible underlying mechanisms. First, both D2- and D3-receptors may be involved. It might be countered that, by analogy to binding isotherms for drugs displaying differential affinity at two radiolabeled sites, dose-response curves for S33084 and GR218,231 should be biphasic. However, this would be difficult to detect in vivo. Second, colocalized D3- and D2-receptors (Surmeier et al., 1992) interact via multiple transduction mechanisms (Shafer and Levant, 1998; Missale et al., 1998). Such cooperative effects could explain drug potencies at cerebral sites differing from those at cloned, heterologously expressed, pure populations of hD3- or hD2-receptor. Third, a further possibility in this light would be the formation of D3/D2-receptor heterodimers, in analogy to μ/δ-opioid and muscarinic (M2/M3) receptors (Jordan and Devi, 1999; Maggio et al., 1999). And fourth, in analogy to 5-hydroxytryptamine2C receptors (Fitzgerald et al., 1999), specific (hetero- or monomeric) cerebral isoforms of rat D3-receptor (Jung et al., 1999) with pharmacological profiles differing from cloned D3-sites may be involved. Thus, although D2-receptors are involved in hypothermia, mechanisms underlying the influence of S33084 on hypothermic actions of dopaminergic agonists, and the role of D3-receptors in thermoregulatory mechanisms, require further examination.
Yawning, PE, and FI.
The resistance of 7-OH-DPAT-induced yawning behavior (Ahlenius and Larsson, 1995; Kurashima et al., 1995) to S33084 and GR218,231 suggests that this behavior is not mediated by D3-receptors. Rather, its blockade by L741,626 implicates D2 sites, probably postsynaptic to dopaminergic neurons and localized in the hypothalamic paraventricular nucleus (PVN) (Melis et al., 1987; Bristow et al., 1996; Argiolas and Melis, 1997). Although the PVN is likewise implicated in dopaminergic processes eliciting PE, underlying mechanisms differ from yawning (Melis et al., 1987). In line with this supposition, S33084 and GR218,231 interfered with induction of PE by 7-OH-DPAT, suggesting a possible role of D3-receptors. Because similar doses of 7-OH-DPAT elicit yawns (D2) and PE (D3), this observation underpins studies questioning its selectivity for D3 sites in vivo (see the Introduction; Shafer and Levant, 1998). However, active doses of S33084 and GR218,231 are too high to be attributable to a selective role of D3-receptors (vide supra). Furthermore, although the inhibitory action of L741,626 also implicates D2-receptors in mediation of PE, doses of S33084 and GR218,231 likewise do not correspond to those blocking D2-receptors. Thus, in analogy to hypothermia (see above), alternative mechanisms involving D3-receptors may underlie inhibition of PE by S33084. Postsynaptic D2-receptors in perifornical hypothalamus are implicated in anorexic actions of DA (Leibowitz, 1980;Ladurelle et al., 1991), and the resistance of 7-OH-DPAT-induced hypophagia to S33084 and GR218,231 versus L741,626 suggests involvement of D2-receptors. Anatomical observations support a major role of D2-receptors in mediation of hypophagia, yawns, and PE. Thus, the density of D3-receptors in the PVN, and in the perifornical and other hypothalamic nuclei is low relative to D2-receptors (Bouthenet et al., 1991; Hall et al., 1996; Gurevich and Joyce, 1999).
Spontaneous Locomotor Behavior.
In three complementary paradigms, S33084 (and GR218,231) did not enhance spontaneous locomotor activity. These observations contradict the hypothesis that postsynaptic D3-receptors in nucleus accumbens exert a major inhibitory influence on locomotor behavior. This suggestion was based on findings that D3antagonists of lesser selectivity, U99194 and nafadotride, enhance locomotion in rodents habituated to an open-field (Haadsma-Svensson and Svensson, 1998), whereas 7-OH-DPAT decreased locomotion independently of DA release (De Boer et al., 1997; Haadsma-Svensson and Svensson, 1998). Furthermore, antisense neutralization of D3-receptors prolonged locomotor behavior (Ekman et al., 1998) and early studies of D3-receptor deficient mice revealed a mild and transient hyperactivity (Baik et al., 1995; Accili et al., 1996; Xu et al., 1997; Kelly et al., 1998). In line with the present data, however, recent studies of knockout mice (Boulay et al., 1999a,b; Xu et al., 1999) showed that lack of D2- but not D3-receptors eliminates hypolocomotor actions of 7-OH-DPAT. Moreover, there are no differences in motor-stimulant actions of U99194 and nafadotride between wild-type and D3 mutant animals. Finally, like S33084, S14297 does not increase spontaneous locomotion (Millan et al., 1995; M.B., unpublished observation). Inactivation of D3-receptors does not, thus, consistently enhance locomotor activity. Indeed, S33084 and GR218,231 mimicked the inhibitory influence of L741,626 on locomotor behavior in rats, in line with findings of Jung et al. (1999) that deletion of D3- and D2-receptors synergistically reduces locomotor behavior in mice.
Rotation.
In line with the above-mentioned comments, neither S33084 nor GR218,231 elicited rotation in unilateral SNPC-lesioned rats, and a microinjection study concluded that D3-receptors do not initiate turning behavior from nucleus accumbens (Koshikawa et al., 1996). Notably, induction of rotation by quinpirole was inhibited by L741,626 but unaffected by S33084 and GR218,231. These observations support the view that D2-receptor activation is the principal mechanism underlying antiparkinsonian properties of dopaminergic agonists (Baik et al. 1995; Newman-Tancredi et al., 1997; Kelly et al., 1998; Shafer and Levant, 1998). However, the possible relevance of striatal D3-receptors in interaction with colocalized D1-receptors justifies further investigation (Xu et al., 1997; Perachon et al., 1999; Pilla et al., 1999).
Potential Antipsychotic Activity.
Of various models examined, S33084 only attenuated climbing behavior elicited by apomorphine. This model is highly sensitive to D2-receptor blockade (Millan et al., 1998b, 2000a; Moore, 1999), and active dose ranges of S33084 (and GR218,231) in this paradigm suggest that their effects reflect residual activity at D2-receptors (vide supra). In a further, “classic” model, CAR, S33084, and GR218,231 were, in contrast, inactive. Furthermore, L741,626 was effective only over a higher dose range, consistent with studies showing that only elevated doses of antipsychotics are active in this procedure (Millan et al., 1998b, 2000a; Moore, 1999). The present data are, thus, in line with correlation analyses with diverse antipsychotics suggesting a key role of D2-receptors in both the apomorphine-climbing and CAR models (Millan et al., 1995, 1998a, 2000a;Moore, 1999). S33084 only weakly inhibited the locomotor response to the psychostimulants amphetamine and cocaine compared with GR218,231, whereas L741,626 showed robust activity. These observations also correspond to the marked correlation between antipsychotic affinity at D2-receptors and blockade of amphetamine-locomotion (Millan et al., 1995, 1998a, 1999, 2000a). Furthermore, in D3-receptor-deficient mice, the hyperlocomotion elicited by amphetamine is not attenuated (Xu et al., 1997). Finally, serotonergic mechanisms predominantly mediate the locomotor response to PCP (Martin et al., 1997; Svensson et al., 1995), which was resistant to S33084 as well as GR218,231 and L741,626.
This inactivity of S33084 is consistent with analyses suggesting (with the exception of PCP locomotion) a predominant role of D2-receptors (Millan et al., 1995, 1998a, 1999,2000a; Moore, 1999). Moreover, Ralph et al. (1999) suggested that D2 sites are involved in disruption of prepulse inhibition by amphetamine. Despite the interest of these observations, no antipsychotic currently used clinically differentiates D2- from D3-receptors. Thus, evaluation of drugs interacting selectively with D3- or D2-receptors in schizophrenic patients may be necessary to definitively establish whether D3- and/or D2-receptors are involved in the pathogenesis and treatment of schizophrenia. Regarding the potential significance of D3-receptors, it is worth recalling alterations in D3-receptor expression after chronic antipsychotic treatment in rodents (Harrison, 1999); changes of D3-receptor density in psychotic patients, normalized by antipsychotic treatment (Gurevich et al., 1997); an association of D3-receptor gene polymorphisms with the occurrence of schizophrenia (Kerwin and Owen, 1999); and a role of D3-receptors in the modulation of reward (Pilla et al., 1999).
Extrapyramidal Activity.
In models reflecting perturbation of striatally integrated motor performance, induction of catalepsy and inhibition of methylphenidate-induced gnawing, S33084 and GR218,231 were inactive, whereas L741,626 was effective. These data suggest that blockade of D2-receptors elicits extrapyramidal, motor side effects. In distinction, selective blockade of D3-receptors, scarce in the striatum (Shafer and Levant, 1998), is ineffective. These findings are of note inasmuch as correlation analyses failed to differentiate the role of D3- versus D2-receptors in induction of extrapyramidal effects (Millan et al., 1995, 1998a). Furthermore, there are contradictory data from D2-receptor-deficient mice as concerns the occurrence of “catalepsy” and parkinsonian symptoms (Baik et al., 1995; Kelly et al., 1998; Boulay et al., 1999a; Jung et al., 1999; Xu et al., 1999). The lack of influence of S33084 and GR218,231 compared with L741,626 on PRL levels corresponds to the high density of (tonically active) D2-receptors on lactotrophs, whereas D3-receptors are absent from the pituitary and the tubero-infundibular dopaminergic pathway (Durham et al., 1997).
Summary and Conclusions
S33084 was inactive in diverse models of potential antipsychotic and extrapyramidal activity, and it neither enhanced spontaneous locomotor behavior nor modified induction of yawning behavior and hypophagia by dopaminergic agonists. However, it attenuated induction of hypothermia and PE by 7-OH-DPAT (and PD128,907). A similar profile was acquired for GR218,231. As discussed above, then, further exploration of the mechanisms of action of S33084 in attenuating hypothermia and PE, and of the involvement of D3-receptors in these responses, would be of interest. Notably, in no model could an exclusive role of D3-receptors be revealed. In contrast, D2-receptors were implicated in all paradigms as revealed by the robust activity of L741,626, which was effective in models predictive of antipsychotic actions at doses only slightly lower than those predictive of extrapyramidal actions. Nevertheless, the genuine significance of D2- versus D3-receptors in the etiology and treatment of schizophrenia may only become apparent on evaluation of selective ligands in human. Finally, as concerns experimental studies, S33084 should comprise an instructive tool for further elucidation of the pathophysiological significance of D3-receptors.
Acknowledgments
We thank C. Langaney for secretarial assistance and B. Denorme, S. Girardon, H. Gressier, L. Iob, and S. Veiga for technical assistance.
Footnotes
-
Send reprint requests to: Dr. Mark J. Millan, Institut de Recherches Servier, Centre de Recherches de Croissy, Psychopharmacology Department, 125 Chemin de Ronde, 78290 - Croissy-sur-Seine, France.
- Abbreviations:
- S33084
- (3aR,9bS)-N-[4-(8-cyano-1,3a,4,9b-tetrahydro-3H-benzopyrano[3,4-c]pyrrole-2-yl)-butyl]-(4-phenyl) benzamide
- GR218,231
- 2(R,S)-(di-n-propylamino)-6-(4-methoxyphenylsulfonylmethyl)-1,2,3,4-tetrahydronaphthalene
- L741,626
- 4-(4-chlorophenyl)-1-(1H-indol-3-ylmethyl)piperidin-4-ol
- DA
- dopamine
- CT
- core temperature
- PD128,907
- (+)-(4aR,10bR)-3,4,4a,10b-tetrahydro-4-propyl-2H,5H-[1]benzopyrano-[4,3-b]-1,4-oxazin-9-ol) HCl
- 7-OH-DPAT
- 7-hydroxy-2-dipropylaminotetralin
- PE
- penile erection
- CAR
- conditioned avoidance response
- NMDA
- N-methyl-d-aspartate
- PCP
- phencyclidine
- SNPC
- substantia nigra, pars compacta
- PRL
- prolactin
- FI
- food intake
- AD
- active dose
- PVN
- paraventricular nucleus
- Received October 25, 1999.
- Accepted February 21, 2000.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}