


Abstract
We previously demonstrated that sustained depletion of methylguanine DNA methyltransferase (MGMT) activity is required for optimal reversal of chloroethylnitrosourea resistance in tumor cells. The purpose of this study was to design O6-benzylguanine (BG) treatments that deplete MGMT activity in tumor cells and xenograft tumors in a prolonged manner. When SF767 cells were treated with a bolus dose of BG (25 μM for 1 h), >95% of MGMT activity was depleted but 33% of the activity recovered within 24 h. In contrast, MGMT activity was completely depleted for 24 h when cells were pretreated with a low dose of BG (2.5 μM) for 24 h, followed by the bolus dose and same low-dose treatment for 24 h. This combination regimen of pre- and post-treatments with a bolus dose sensitized cellsN,N′-bis(2-chloroethyl)-N-nitrosourea in vitro by ∼2-fold more than the bolus dose alone. Similar BG treatment with Alzet micro-osmotic pumps produced sustained inhibition of MGMT activity in vivo. In xenograft SF767 tumors, low-dose pre- and post-treatments (8 mg/kg over 24 h) combined with an i.p. bolus dose (80 mg/kg) of BG inhibited >95% of MGMT activity for 24 h after the bolus. The bolus dose alone did not deplete MGMT for 24 h. These results demonstrate that combination low-dose and bolus BG treatment is superior to the bolus dose alone in depleting MGMT activity in a sustained manner in vitro and in vivo. When combined withN,N′-bis(2-chloroethyl)-N-nitrosourea treatment, this BG regimen also should also produce greater antitumor activity than the single bolus dose evaluated clinically.
The chloroethylnitrosourea (CENU) class of antitumor agents is clinically used to treat a number of tumors, including brain neoplasms, mailgnant melanoma, advanced lymphomas, and gastrointestinal carcinomas (Berger, 1993). However, these agents are not considered to be curative therapies. A primary reason for the suboptimal efficacy of CENUs is the inherent resistance of many tumors to these agents. The cytotoxic mechanism of CENUs involves a rapid chloroethylation at theO6 position of guanine DNA residues, followed by a slow intramolecular cyclization to produce anO6-ethanoguanine intermediate. The cytosine of the opposite strand then attacks the ethanoguanine, producing a lethal interstrand cross-link (Kohn, 1977; Tong et al., 1982). This secondary reaction takes place over several hours, so that a temporal window exists in which the chloroethyl adduct can be repaired (Lemoine et al., 1991).
The DNA repair proteinO6-methylguanine DNA methyltransferase (MGMT) efficiently removes alkyl groups from guanine before cross-link formation, and also can react with the ethanoguanine intermediate (Brent and Remack, 1988; Gonzaga et al., 1992). The former mechanism involves a stoichiometric reaction in which the chloroethyl moiety is transferred from the guanine to cysteine residue 145 of MGMT, forming a covalent bond. The guanine is thus restored, and the MGMT molecule is irreversibly inactivated. New repair activity, therefore, requires de novo protein synthesis (Kroes and Erickson, 1995). In various tumor cell lines and human tumors, MGMT has been shown to function in CENU resistance (Robins et al., 1983; Dolan et al., 1986, 1988; Pegg, 1990). Enhanced MGMT expression by gene transfer in murine hemopoietic cells also increased resistance to CENUs (Allay et al., 1995; Jelinek et al., 1996; Maze et al., 1996, 1997). Collectively, these data suggest depletion of MGMT in tumors before CENU therapy might enhance chemotherapy against neoplasms normally resistant to these agents.
Low-molecular weight compounds such as the DNA-methylating agents streptozotocin (STZ) and dacarbazine have been used to potentiate CENU cytotoxicity in vitro and in vivo. For instance, STZ depleted MGMT activity by 75% and produced a 2- to 3-log increase in cell killing byN,N′-bis(2-chloroethyl)-N-nitrosourea (BCNU) in vitro (Futscher et al., 1989). In human trials, three injections of STZ (500 mg/m2) reduced MGMT activity in lymphocytes by 75% (Gerson, 1989). However, in phase II clinical trials directed at refractory melanoma (Smith et al., 1996) and metastatic colon cancer (Wilson et al., 1995), the combination of STZ and BCNU did not significantly improve the therapeutic efficacy. In the latter study, STZ also was found to deplete MGMT levels in lymphocytes but not the target metastatic colon cancer cells. Clinical trials of combination methylating and chloroethylating agent therapy indicated the dose-limiting toxicity to be hematological, although significant hepatic and pulmonary toxicities also were observed (Micetich et al., 1992; Gerard et al., 1993; Wilson et al., 1995).
The free base O6-methylguanine (MG) inactivates MGMT by acting as a direct substrate, thereby circumventing many of the genotoxic and mutagenic properties of DNA-methylating agents. In vitro, MG significantly reduced cellular MGMT activity and sensitized tumor cells to CENUs (Dolan et al., 1986, 1988; Gerson et al., 1988). In mice, MG reduced MGMT levels to 35 and 25% of control levels in the liver and xenograft tumors, respectively. However, this treatment did not enhance of the therapeutic effectiveness of CENUs against xenograft HT29 human colon tumors (Dolan et al., 1989). In addition, MG exhibited poor solubility characteristics, requiring very high doses to achieve therapeutic efficacy. Therefore, this agent was not suitable for human clinical trials.
In recent years, numerous studies have demonstrated depletion of MGMT and sensitization to CENU in human tumor cells by the free baseO6-benzylguanine (BG). For example, micromolar concentrations of BG inhibited >90% of cellular MGMT within 10 min and completely depleted MGMT within 1 h in HT29 cells (Dolan et al., 1990a). Treatments with BG also markedly potentiated CENU toxicity in HT29 and other human tumor cells normally resistant to CENU (Dolan et al., 1991; Baer et al., 1993;Magull-Seltenreich and Zeller, 1995). Furthermore, growth rates of xenograft human brain and colon tumors in nude mice were significantly inhibited when mice were treated with BG before BCNU compared with BCNU alone (Dolan et al., 1990b, 1993; Friedman et al., 1992; Mitchell et al., 1992; Felker et al., 1993; Gerson et al., 1993; Wedge and Newlands, 1996; Kurpad et al., 1997; Phillips et al., 1997). All of these studies used a single bolus injection of BG before BCNU therapy. Despite promising results, partial but significant recovery of MGMT activity was observed in the tumors within 24 h of BCNU treatment. The recovery in MGMT activity at 24 h after BCNU treatment ranged from 16 to 33% of the initial tumor levels (Gerson et al., 1993; Wedge and Newlands, 1996; Phillips et al., 1997; Wedge et al., 1997). In some cases, substantial recovery was evident within 15 h of BCNU treatment (Mitchell et al., 1992; Gerson et al., 1993). This dosing regimen also has recently been used in clinical trials (Friedman et al., 1998; Spiro et al., 1999). It has previously been suggested byMarathi et al. (1994) that optimal reversal of BCNU resistance requires complete inactivation of MGMT for at least 24 h after BCNU administration. Therefore, we have compared various BG treatments in vitro and report a combination regimen that completely inactivates MGMT for 24 h in xenograft human glioma tumors.
Experimental Procedures
Materials.
BCNU was obtained from the Developmental Therapeutics Branch, National Cancer Institute (Bethesda, MD). BCNU was dissolved in 100% ethanol, and diluted at least 100-fold in culture medium for experiments. BG was purchased from Sigma Chemical Co. (St. Louis, MO). For cell culture experiments, BG was dissolved in dimethyl sulfoxide and diluted at least 100-fold in the medium. For in vivo administration, BG was added to polyethylene glycol-400 prewarmed to 37°C and subsequently diluted to 40% polyethylene glycol-400 (v/v) in PBS.
Cell Culture.
The human glioma cell line SF767 was provided by The Brain Tumor Research Center (University of California-San Francisco). SF767 cells were cultured in Eagle's minimum essential medium supplemented with 10% bovine calf serum (Hyclone Laboratories Inc., Logan UT), 1% l-glutamine, HEPES buffer, glutamine, and 2% penicillin-streptomycin (Life Technologies, Inc, Grand Island, NY). Cells were maintained in logarithmic growth phase at 37°C in 5% CO2 atmosphere.
In Vitro BG Treatments.
For MGMT inactivation studies, 2 × 105 SF767 cells were seeded for 24 h in 5 ml of normal culture medium per 25-cm2 flask. The cells were then exposed to various BG treatments under normal culture conditions. For the bolus dose treatment, cells were treated with 25 μM BG for 1 h, and then washed four times with complete medium before replacing with fresh, complete medium. Cells also were administered a BG pretreatment with or without the bolus dose, which consisted of treatment with 2.5 μM BG continuously for 24 h before the time at which the bolus dose of BG was administered. A BG post-treatment also was used in conjunction with the pretreatments and bolus treatments. The post-treatment consisted of exposure to 2.5 μM BG for 24 h immediately following the bolus BG dose. Cellular extracts to measure MGMT activity were prepared at the end of the 24-h pretreatment and at 1, 8, and 24 h after the bolus dose treatment. All control samples were treated with a corresponding dose of the respective vehicle. To analyze possible potentiation of BCNU cytotoxicity, 3 × 105 SF767 cells were seeded for 24 h, then treated with same above-mentioned BG treatments. At the end of these respective treatments, the cells were trypsinized and seeded in triplicate at a density of 2 × 102 cells/10-cm dish in 10 ml of complete medium. Cells were incubated for 10 to 12 days under normal culture conditions, and then colonies were fixed in methanol and stained with methylene blue in phosphate buffer. Colonies were enumerated and expressed as the mean and standard error of triplicate dishes for two independent experiments.
Xenograft Tumor Studies.
Animal protocols were approved by the Animal Use Committee of Indiana University School of Medicine. Naturally obese diabetic mice with severe combined immunodeficiency (NOD/SCID) were maintained in microisolator cages with sterile bedding, food, and water. Male and female NOD/SCID mice at 9 to 12 weeks of age were inoculated s.c. in the flank with 10 × 106 SF767 cells suspended in 0.1 ml of Hanks' balanced saline solution containing 1 mM HEPES. When tumors were palpable (∼3 weeks later), the mice were divided into treatment groups. The four groups were comprised of control animals and those implanted s.c. with one, two, or three Alzet pumps (Alza Corp., Palo Alto, CA) per mouse. Each pump was designed to deliver 1 μl of BG, or 0.003 mg/h, for 72 h. Thus, mice bearing one, two, or three pumps received 24-h cumulative doses of 0.07, 0.13, and 0.2 mg of BG, respectively. With the average weight of 25 g/mouse, the respective cumulative 24-h doses were ∼2.8, 5.2, and 8.0 mg/kg. Mice were sacrificed from each group at 24-h postimplantation, and the tumors were excised, frozen in liquid nitrogen, and stored at −80°C. The remaining mice received a bolus i.p. injection of BG (80 mg/kg) at 24-h after pump implantation; control mice received an equivalent dose of vehicle. Tumors were then harvested at 1, 12, and 24 h after the bolus injection.
Measurement of MGMT Activity.
Cultured cells (1.5 × 106) were resuspended in 400 μl of assay buffer (50 mM Tris, pH 8.0, 1 mM dithiothreitol, 1 mM EDTA, 5% glycerol). Cell extracts were prepared by sonication on ice followed by centrifugation to clarify the supernatant. Tumor extracts were prepared by thawing frozen tumors and resuspending each sample in ∼3 ml of assay buffer per milligram of tumor weight. Tumors were then homogenized on ice three times for 30 s each, sonicated, and centrifuged. Protein content was quantitated with the Bradford protein assay. The MGMT assay was performed as described previously (Futscher et al., 1989) with some modification. Briefly, an 18-bp oligomer was synthesized to contain theO6-methylguanine lesion within aPvuII restriction site. This oligo was radiolabeled by filling in the 3′ recessed end of the complementary 16-bp strand with [α-32P]thymidine 5′-triphosphate (NEN, Boston, MA). MGMT activity was measured by incubating 0.2 pmol of the radiolabeled probe with 25 μg of total cellular protein at 37°C for 2 h, followed by phenol-chloroform extraction to remove cellular protein and ethanol precipitation of the probe. The probe was then digested withPvuII (Boehringer Mannheim, Indianapolis, IN) and electrophoresed on a 20% denaturing polyacrylamide gel. MGMT activity is proportional to the amount of radiolabeled 8-bp fragment produced. Results were quantitated on a Storm 860 PhosphorImager (Molecular Dynamics, Sunnyvale, CA).
Statistical Analysis.
Single-value treatments with BG were compared with their respective control (vehicle) treatment using Student's t test to determine the significance of differences. Multiple value BG treatments were compared with control and each other by repeated-measures ANOVA. Differences between treatments were considered to be significant at P< .05 unless otherwise stated. LC50 and LC90values were determined from the semilogarithmic survival curves. BG sensitivity was compared between treatments by ANOVA. All data are presented as the means ± S.E. from at least three independent measurements unless otherwise stated.
Results
Prolonged Depletion of MGMT Activity by Continuous, Low-Dose BG Treatment in SF767 Cells.
Untreated SF767 cells displayed moderately high MGMT activity (Fig. 1, lane 1). When the cells were treated with only a high dose (25 μM) of BG for 1 h (bolus dose), cellular MGMT activity was completely inhibited (>95% ± 0.3%) 1 h later (lane 2). This depletion was maintained at 8 h (lane 3), but MGMT levels had recovered to 33% ± 1.5% of the initial level at 24 h (lane 4). Alternatively, in cells pretreated with 2.5 μM BG for 24 h without the bolus dose, MGMT activity was depleted by only 87% ± 1.3% (lane 10) with ∼26% ± 1.6% recovery 24 h later (lane 11). The combination of these two treatments, 2.5 μM BG for 24 h before the bolus dose, depleted >95% ± 0.4% of cellular MGMT activity at 1 h after the bolus (lane 5). However, MGMT activity recovered faster following this combination treatment than the bolus alone, with 15% ± 1.4% recovery at 8 h (lane 6) and 47% ± 2.2% at 24 h (lane 7). In contrast, when a 24-h post-treatment with 2.5 μM BG was added following the pretreatment and bolus dose, the MGMT recovery was blocked and >95% of MGMT activity was inactivated at 8 and 24 h after the bolus dose (lanes 8 and 9, respectively). Hence, these results demonstrate that the combination of pre- and post-treatments for 24 h with a low concentration (2.5 μM) of BG in conjunction with a 1-h bolus dose (25 μM) of BG significantly enhanced the extent and duration of MGMT inactivation compared with the bolus dose alone. Most importantly, this combination treatment depleted >95% of cellular MGMT activity for 24 h.

Combination regimen of continuous low-dose and high-dose BG treatments produces prolonged depletion of MGMT activity in SF767 human glioma cells. SF767 cells were exposed to various BG treatments and harvested at the times indicated for MGMT measurement. MGMT activity in untreated cells (lane 1) was used as a reference for MGMT recovery following BG treatment. In lanes 2 to 4, cells were treated with 25 μM BG for 1 h and then harvested at 1, 8, and 24 h after the onset of treatment, respectively. In lanes 10 and 11, cells were treated for 24 h with 2.5 μM BG and then harvested at 1 and 24 h after removal of the BG, respectively. In lanes 5 to 7, cells were pretreated for 24 h with 2.5 μM BG and then 25 μM BG for 1 h, and harvested at 1, 8, and 24 h after the onset of the bolus treatment, respectively. In lanes 8 and 9, cells were pre- and post-treated with 2.5 μM BG for 24 h in conjunction with the bolus dose (25 μM BG for 1 h), and cells were harvested at 8 and 24 h after onset of the bolus treatment, respectively. Two independent experiments were performed with consistent results.
Potentiation of BCNU Cell Killing In Vitro by Continuous, Low-Dose and High-Dose BG Treatments.
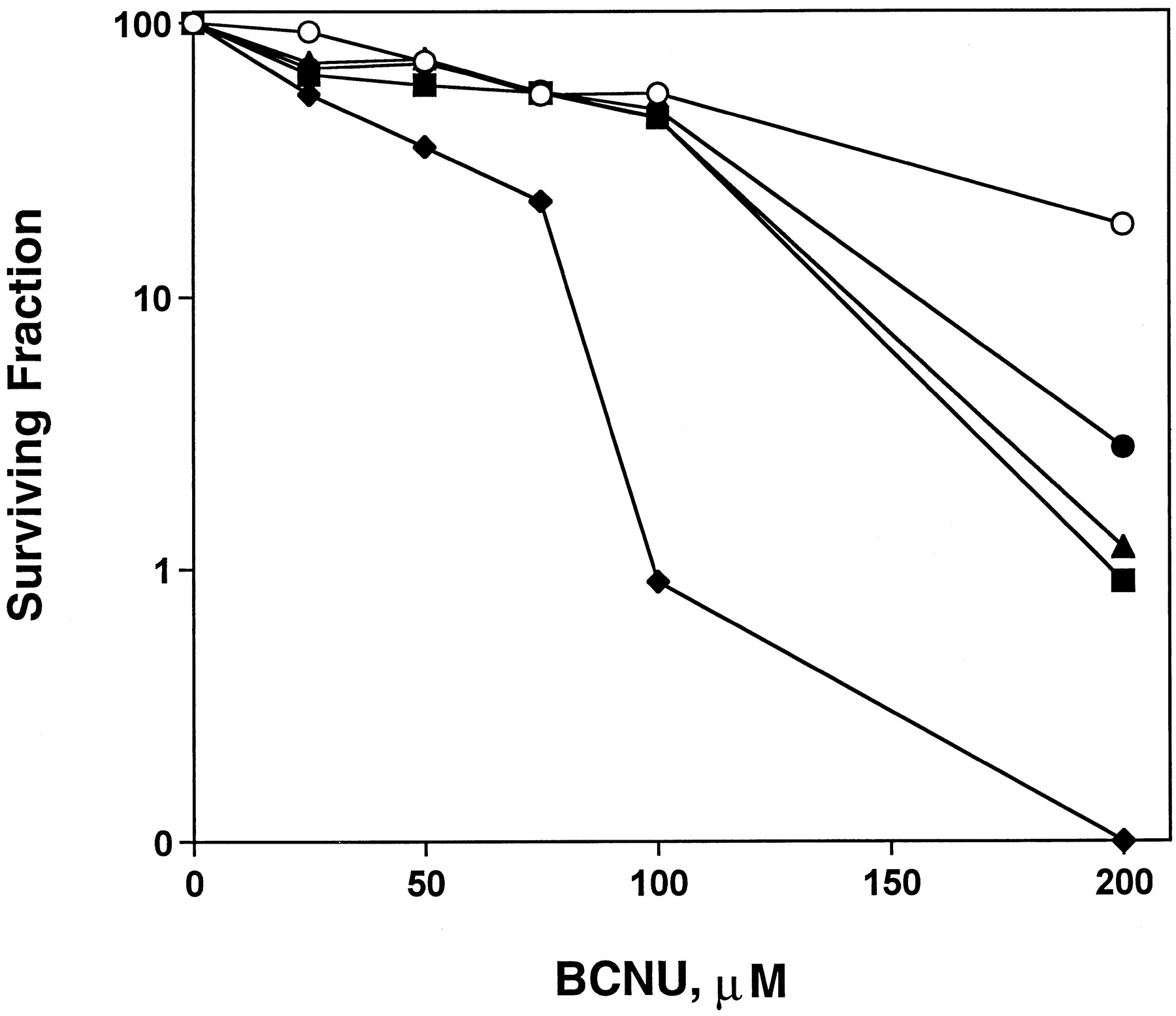
Survival of colony-forming cells was used to compare BCNU sensitivity in SF767 cells treated with various BG treatments (Fig. 2). Cells treated with only BCNU (open circles) exhibited an LC50 value of ∼100 μM and an LC90 value of ∼250 μM, as shown in Table1. The 24-h continuous pretreatment with 2.5 μM BG, which depleted ∼87% of cellular MGMT activity, potentiated BCNU cell killing by only 1.6-fold, as determined by the LC90 value (closed circles). When the bolus high-dose (25 μM) of BG, which inhibited >95% of MGMT activity, was administered immediately before BCNU treatment (triangles), BCNU toxicity was potentiated by ∼1.8-fold. When the pretreatment and bolus BG treatments were combined (squares), BCNU sensitivity was not further enhanced compared with the bolus alone. However, when cells were pretreated with 2.5 μM BG for 24 h in addition to the bolus BG treatment before BCNU exposure, then post-treated with 2.5 μM BG for 24 h subsequent to BCNU exposure (diamonds), BCNU cytotoxicity was potentiated by 2.8-fold, as indicated by the LC90 value. Thus, the combination dosing regimen of 24-h continuous pre- and post-treatments with a low concentration of BG and 1-h bolus dose was significantly more effective in reversing BCNU resistance than the low-dose pretreatment or bolus high-dose BG treatment alone, as determined by ANOVA (P <.05).

Continuous, low-dose BG treatment with or without high-dose BG treatment increases cell killing by BCNU. Colony formation assays were performed on SF767 cells treated with various concentrations of BCNU for 1 h (○). Cells pretreated with 2.5 μM BG for 24 h (●) or 25 μM BG for 1 h (▴) immediately before BCNU treatment were only slightly sensitized to BCNU. Combination treatment with 2.5 μM BG for 24 h and 25 μM BG for 1 h (▪) immediately before BCNU treatment did not enhance the BCNU sensitization compared with the bolus treatment alone. Post-treatment with 2.5 μM BG for 24 h after pretreatment with 2.5 μM BG for 24 h and 25 μM BG for 1 h (♦) significantly sensitized the cells to BCNU. Survival curves represent the mean of triplicate flasks from two experiments; all standard error values were <10% of the respective mean. The LC50 and LC90 values are summarized in Table 1.
Comparison of BCNU LC50 and LC90 values among BG treatments in SF767 cells
Prolonged Depletion of MGMT Activity by Combination Continuous and Bolus BG Treatment in SF767 Xenograft Tumors In Vivo.
Mice bearing SF767 xenograft tumors were treated with a continuous, low dose of BG with surgically implanted osmotic pumps throughout the experiments. The cumulative doses of BG administered over 24 h to mice implanted with one, two, and three pumps were 2.8, 5.2, or 8.0 mg/kg, respectively. After these 24-h continuous, low-dose pretreatments, mice received a bolus BG injection of 80 mg/kg. Tumor MGMT activity was measured at 1, 12, and 24 h after the bolus injection, during which time the mice continued to receive the low doses of BG administered by the pumps. The untreated tumors exhibited comparable MGMT activity to SF767 cells in vitro (Fig.3, lane 1). The bolus injection alone (80 mg/kg) depleted 86% ± 2.9% and 92% ± 1.4% of the MGMT activity in the tumors at 1- and 12-h postinjection, respectively (lanes 14 and 15). At 24 h, however, the tumor MGMT activity had recovered to ∼33% ± 1.2% of the initial level (lane 16). After 24 h of continuous pretreatment with one pump at 0.12 mg of BG/kg/h (2.8 mg/kg in 24 h, or 3.5% of the bolus dose), only 29% ± 2.4% of the tumor MGMT activity was inhibited compared with control (lane 10 versus lane 1). This continuous pre- and post-treatment only slightly enhanced the depletion of tumor MGMT activity at 1, 12, and 24 h after the bolus compared with the bolus alone (lanes 11–13 versus lanes 14–16, respectively). Pretreatment for 24 h with two pumps at 0.22 mg of BG/kg/h (5.2 mg/kg in 24 h, or 6.5% of the bolus dose) depleted 37% ± 3.1% of the tumor MGMT activity (lane 6 versus lane 1). The pre- and post-treatment at this dose also depleted tumor MGMT activity only slightly more than the bolus alone at 1, 12, and 24 h after the bolus (lanes 7–9 versus lanes 14–16, respectively). About 23% ± 1.8% of MGMT activity had recovered at 24 h (lane 9) compared with 33% with the bolus alone (lane 16). Animals implanted with three pumps received 0.33 mg of BG/kg/h (8.0 mg/kg in 24 h, or one-tenth of the bolus dose) for 24 h before and after the bolus dose (80 mg/kg). In these animals, >90% ± 1.3% of the tumor MGMT activity was inhibited after the 24-h continuous pretreatment alone (lane 2 versus lane 1). The bolus injection (80 mg/kg) further depleted tumor MGMT levels to nearly undetectable levels at 1, 12, and 24 h after the bolus (lanes 3–5, respectively). These results are summarized in Table 2 and demonstrate that continuous, 24-h pre- and post-treatment with 0.33 mg of BG/kg/h significantly potentiates inactivation of MGMT in xenograft tumors compared with a single bolus BG injection of 80 mg/kg. Furthermore, the combination regimen of 24-h pre- and post-treatments at a cumulative dose of one-tenth of the bolus (8 mg/kg) and the single bolus injection (80 mg/kg) resulted in >95% depletion of tumor MGMT activity for 24 h after the bolus.

Combination continuous, low-dose and bolus BG treatments produces prolonged depletion of MGMT activity in xenograft SF767 tumors. Mice bearing SF767 xenograft tumors were continuously treated with various low doses of BG for 48 h by surgically implanted osmotic pumps. Each pump delivered BG at 0.12 mg/kg/h, or 2.8 mg/kg over 24 h. Mice were implanted with one (lanes 2–5), two (lanes 6–9), or three (lanes 10–13) pumps. At 24-h postimplantation, mice were administered a bolus i.p. injection of BG at 80 mg/kg. The bolus also was administered to mice without any pumps (bolus only, lanes 14–16). Mice were euthanized and tumors were harvested for MGMT measurement 24 h after pump implantation (lanes 2, 6, and 10) and at 1 (lanes 3, 7, and 11), 12 (lanes 4, 8, and 12), and 24 h (lanes 5, 9, and 13) after the bolus treatment. MGMT activity was compared in tumors of untreated control mice (lane 1). The data shown are from one representative experiment with one animal per treatment; two independent experiments were performed with reproducible results. The results were quantitated by phosphorimage analysis and are shown in Table 2.
MGMT depletion in xenograft SF767 tumors by various BG treatments
Discussion
Numerous studies have successfully used BG to reverse BCNU resistance in human tumor cells in vitro (Futscher et al., 1989; Dolan et al., 1990a; Baer et al., 1993; Magull-Seltenreich and Zeller, 1995) and in vivo (Dolan et al., 1990b, 1993; Friedman et al., 1992; Mitchell et al., 1992; Felker et al., 1993; Gerson et al., 1993; Wedge and Newlands, 1996; Kurpad et al., 1997; Phillips et al., 1997). In all but one of these studies, MGMT activity in the xenograft tumors partially recovered within 24 h of the BCNU treatment. We previously demonstrated that maximal sensitization of CENU-resistant cells requires prolonged and complete depletion of cellular MGMT following BCNU treatment (Marathi et al., 1994). In that study, we found BG regimens producing sustained and complete (>95%) inhibition of MGMT activity for 24 h following BCNU treatment provided the greatest enhancement of BCNU cytotoxicity. This is probably due to the delayed formation of lethal cross-links following BCNU exposure. Hence, it is necessary to maintain minimal MGMT activity until the maximal number of cross-links is formed to attain the greatest increase in BCNU sensitivity. The objective of the current study was therefore to examine optimal BG regimens that inactivate >95% of MGMT activity for 24 h in xenograft SF767 tumors. We first examined several BG treatments in vitro for MGMT depletion and potentiation of BCNU cytotoxicity.
Two combination BG regimens were compared versus a high-dose, or bolus, BG treatment alone for depletion of MGMT in SF767 cells. The BG concentration of 25 μM was chosen for the bolus dose because it has been suggested that the maximally tolerated plasma concentration of BG in humans is ∼30 μM (Friedman et al., 1998). This 1-h treatment inhibited >95% of the MGMT activity immediately after exposure, but a typical pattern of MGMT recovery occurred within 24 h, with about one-third of the initial MGMT activity observed at 24 h. This extent of MGMT recovery is comparable to that observed in xenograft tumors following a single bolus injection of BG. For example, 30% of MGMT activity recovered within 24 h in human glioblastoma U87 MG tumors (Wedge and Newlands, 1996). To identify alternative dosing regimens that deplete >95% of tumor MGMT activity for 24 h, we first examined MGMT inactivation and recovery in cells treated continuously for 24 h with a low dose (2.5 μM) of BG. This treatment failed to deplete >95% of MGMT activity at 1- or 24-h post-treatment. When the bolus dose was administered immediately after this 24-h pretreatment, >95% of the tumor MGMT activity was inhibited immediately after the bolus dose. Surprisingly, this combination treatment did not attenuate the MGMT recovery compared with the bolus dose alone. Instead, the rate of MGMT recovery was enhanced by the combination treatment compared with the bolus dose alone, with 47 and 33% of the MGMT activity recovered at 24 h, respectively. This observation suggested that the continuous low-dose treatment may up-regulate MGMT activity, which could potentially be an important factor in designing BG-dosing regimens for future human trials. To block the MGMT recovery observed following the bolus dose with and without pretreatment, a 24-h continuous post-treatment with 2.5 μM BG was added after the pre- and bolus treatments. This combination regimen produced the desired result of >95% depletion of MGMT activity for 24 h. Therefore, these results indicate that 24-h treatments with 2.5 μM BG, or ∼10% of the maximally tolerated plasma BG concentration in humans, administered both before and after a single bolus dose of BG, produce sustained depletion of >95% of MGMT activity in SF767 cells in vitro.
We next examined the ability of these BG regimens to reverse resistance to BCNU in vitro. SF767 cells are resistant to BCNU, with an LC90 value of ∼250 μM, which is comparable to many other human tumor cells inherently resistant to CENUs. Treating the cells for 1 h with 25 μM BG before BCNU treatment sensitized the cells by ∼1.8-fold, as indicated by LC90values. Interestingly, pretreating the cells for 24 h with 2.5 μM BG alone also significantly sensitized these cells (Table 1). This pretreatment inhibited only 87% of the MGMT activity compared with >95% depletion by the high dose alone. However, the levels of MGMT activity at 24 h after BG treatment were comparable between these two treatments, i.e., 33 and 26% after the bolus and pretreatment, respectively. Hence, this observation supports the idea that the level of MGMT activity in tumor cells during the 24-h period following BCNU treatment is more important in modulating BCNU sensitivity than the initial level of MGMT activity. When the pretreatment and bolus dose were combined, BCNU sensitivity was not enhanced compared with the bolus treatment alone despite enhanced recovery in cellular MGMT activity following this regimen. Thus, the extent of recovery in MGMT levels apparently does not correlate directly with the degree of BCNU sensitization among these three treatments, suggesting that a threshold in MGMT levels exists above which BCNU sensitization may not be further attenuated despite higher MGMT levels. The combination regimen that successfully depleted >95% of MGMT activity for 24 h achieved the greatest potentiation of BCNU sensitivity. Continuous treatment for 24 h with a low dose (2.5 μM) of BG both before and after the bolus dose increased the BCNU cytotoxicity by almost 3-fold compared with no BG treatment or the bolus dose alone, as measured by the LC90 values. This result is in agreement with our previous report that complete inactivation of MGMT for 24 h produced the greatest enhancement in BCNU cytotoxicity (Marathi et al., 1994).
These in vitro studies provided a basis for developing a BG treatment regimen to deplete MGMT activity in xenograft SF767 tumors in a prolonged manner in vivo. To mimic the continuous low-dose pre- and post-treatments, surgically implanted Alzet osmotic pumps were used. These pumps deliver a fixed volume of 1 μl, or ∼0.003 mg of BG, per hour for 72 h. The resulting doses of BG administered to mice implanted with one, two, or three pumps were 2.6, 5.2, and 8.0 mg of BG/kg over a 24-h period. A bolus BG dose of 80 mg/kg also was administered 24 h after the pumps were implanted, in parallel with our in vitro experiments. The in vivo results were remarkably similar to the results in vitro. For example, the bolus BG dose alone markedly inhibited the tumor MGMT activity immediately after treatment, but ∼33% of the activity had recovered within 24 h. Also, only the pre- and post-treatments with 8.0 mg of BG/kg, or one-tenth of the bolus dose, inactivated >95% of the tumor MGMT for 24 h after the bolus. Pre- and post-treatments with BG at 2.6 mg/kg had little effect on MGMT depletion compared with the bolus alone, whereas the 5.2-mg/kg treatment only partially attenuated the MGMT recovery compared with the bolus alone. These results demonstrate that MGMT activity in xenograft tumors can be depleted by >95% for 24 h by novel BG-dosing regimens such as the one described in this study. Furthermore, this BG regimen also would be expected to be more effective at potentiating the cytotoxicity of BCNU and other CENU in vivo than the single bolus dose of BG evaluated to date. Therefore, future clinical trials should be aimed at using BG-dosing schedules that deplete tumor MGMT activity in a sustained manner. In addition, our studies indicate that BG regimens that effectively deplete MGMT activity in vitro generally produce similar results in xenograft tumors in vivo. Future studies will examine the ability of various BG treatments to modulate BCNU cytotoxicity in vivo and the relative toxicities of these treatments.
Acknowledgments
We gratefully acknowledge Fengying Luo for assistance in caring for and experimental handling of the NOD/SCID mice.
Footnotes
-
Send reprint requests to: Dr. Leonard C. Erickson, Indiana University Cancer Center, 1044 W. Walnut St., Bldg. R4, Rm. 168, Indianapolis, IN 46268. E-mail: lcericks{at}iupui.edu
-
↵1 This research was supported by National Cancer Institute Grants CA 75426 (to D.A.W.) and CA 45628 (to L.C.E.).
- Abbreviations:
- CENU
- chloroethylnitrosourea
- MGMT
- methylguanine DNA methyltransferase
- STZ
- streptozotocin
- BCNU
- N,N′-bis(2-chloroethyl)-N-nitrosourea
- MG
- O6-methylguanine
- BG
- O6-benzylguanine
- NOD/SCID
- naturally obese diabetic mice with severe combined immunodeficiency
- Received June 10, 1999.
- Accepted August 24, 1999.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}