


Abstract
Polyspecific organic cation transporters in epithelia play an important role in the elimination of many endogenous bioactive amines and therapeutically important drugs. Recently, the first human organic cation transporter (hOCT1) was cloned from liver. The purpose of the current study was to determine the effect of molecular size and hydrophobicity on the transport of organic cations by hOCT1. We studied the interaction of a series of n-tetraalkylammonium (n-TAA) compounds (alkyl chain length, N, ranging from 1 to 6 carbons) with hOCT1 in a transiently transfected human cell line, HeLa. [14C]tetraethylammonium (TEA) uptake was measured under different experimental conditions. Bothcis-inhibition and trans-stimulation studies were carried out. With the exception of tetramethylammonium, all of the n-TAAs significantly inhibited [14C]TEA uptake. A reversed correlation of IC50 values (range, 3.0–260 μM) with alkyl chain lengths or partition coefficients (LogP) was observed.trans-Stimulation studies revealed that TEA, tetrapropylammonium, tetrabutylammonium, as well as tributylmethylammonium trans-stimulated TEA uptake mediated by hOCT1. In contrast, tetramethylammonium and tetrapentylammonium did not trans-stimulate [14C]TEA uptake, and tetrahexylammonium demonstrated an apparent “trans-inhibition” effect. These data indicate that with increasing alkyl chain lengths (N ≥ 2),n-TAA compounds are more poorly translocated by hOCT1 although their potency of inhibition increases. Similar findings were obtained with nonaliphatic hydrocarbons. These data suggest that a balance between hydrophobic and hydrophilic properties is necessary for binding and subsequent translocation by hOCT1.
It is widely recognized that an array of organic cations with diverse chemical structures undergo hepatobiliary secretion (Meijer et al., 1990; Oude Elferink et al., 1995; Groothuis and Meijer, 1996). Distinct transporters for small molecular weight, hydrophilic (type I) and large molecular weight, bulkier (type II) organic cations appear to be involved in the uptake of organic cations across the sinusoidal membrane of the hepatocyte (Mol et al., 1988; Meijer et al., 1990;Steen et al., 1991; Moseley et al., 1992; Oude Elferink et al., 1995;Moseley et al., 1996). Previous studies demonstrated that lipophilicity was a major determinant for the hepatobiliary transport of a series of small molecular mass monoquaternary compounds (<200 Da; type I compounds) in the rat. That is, increasing lipophilicity was associated with increasing hepatic clearances of these compounds (Neef and Meijer, 1984). However, because the studies were carried out in vivo, the specific transporters involved in the hepatic clearance of these compounds were not identified. Furthermore, the structure activity relationships established in these studies were limited to hepatobiliary secretion in the rat. It is not known whether such relationships also describe the hepatobiliary transport of organic cations in the human.
Recently, the first human organic cation transporter (hOCT1) was cloned (Gorboulev et al., 1997; Zhang et al., 1997b). Northern blot analysis demonstrated that hOCT1 is expressed primarily in human liver. Functional studies carried out in Xenopus laevis oocytes suggest that hOCT1 represents an organic cation transporter located on the sinusoidal side of the hepatocyte (Zhang et al., 1997b). To study the functional properties of hOCT1, we developed a transiently transfected cell line, HeLa (Zhang et al., 1998b). We determined the effect of various organic cations and other compounds on the transport of the model organic cation, tetraethylammonium (TEA). Our data suggest that a number of organic cations with diverse structures inhibited TEA uptake in hOCT1 DNA-transfected HeLa cells (Zhang et al., 1998b). In addition, we observed that TEA and 1-methyl-4-phenylpyridinium, known substrates of hOCT1, trans-stimulated the uptake of [14C]TEA. Namely, a high concentration of unlabeled TEA or 1-methyl-4-phenylpyridinium inside the cells stimulated the uptake of [14C]TEA (Zhang et al., 1998b). These data suggest that trans-stimulation studies may be useful in identifying substrates of hOCT1 in the HeLa cell expression system.
Although our previous study demonstrated that structurally diverse organic cations interact with hOCT1, systematic studies ascertaining structure activity relationships were not performed. To obtain insight into the relationship between physicochemical properties, particularly hydrophobicity, and transport by hOCT1, we studied a series ofn-tetraalkylammonium (n-TAA) compounds in hOCT1 transfected HeLa cells. By performing both trans-stimulation and cis-inhibition studies, we determined the effect of hydrophobicity on inhibition potency and translocation by hOCT1. Forn-TAA compounds with molecular weights greater than or equal to 130, we observed a reverse correlation between IC50 and partition coefficient (octanol per water). A reverse correlation was also observed between rate of influx and partition coefficient, indicating that factors other than binding affinity contribute to the overall transport rate of these compounds by hOCT1. These data indicate that a balance between hydrophobic and hydrophilic properties is required for interaction and subsequent translocation by hOCT1.
Experimental Procedures
DNA Isolation.
hOCT1 DNA was subcloned into the mammalian expression vector pTargeT (Promega, Madison, WI) as described previously (Zhang et al., 1998b). DNA for transfection studies was isolated with the Qiagen Endo-free DNA isolation kit (Qiagen, Inc., Valencia, CA). The DNA was resuspended in endotoxin-free TE buffer and its concentration was determined by UV spectroscopy.
Reverse Transcription-Polymerase Chain Reaction (RT-PCR).
The first-strand cDNA for PCR amplification was synthesized from mRNA isolated from various tissues or Caco-2 cells with oligo(dT) primers using the SuperScript preamplification system (Gibco-BRL, Gaithersburg, MD) (Zhang et al., 1997a). The primers used in PCR were designed from the hOCT1 cDNA sequence (5′ and 3′ end primers in Zhang et al., 1997b). PCR was performed in a thermal cycler (Perkin-Elmer, Foster City, CA) using the cycle as described previously (Zhang et al., 1997b). The PCR products were electrophoresed through 1% agarose gel, eluted, and then subcloned into the pGEM-T vector (Promega Biotech). The vector inserts were sequenced by the Biomolecular Resource Center at University of California-San Francisco (Zhang et al., 1997a,b).
Maintenance of Cell Culture and Transfection.
HeLa cells were maintained in growth medium in 175-ml cell culture flasks (Nalge Nunc International, Napierville, IL) at 37°C in a humidified 5% CO2/95% air atmosphere as described previously (Zhang et al., 1998b). All studies were performed in cells of passages 3 to 19. Cells were seeded at a density of 1.6 × 105 cells/well in 12-well tissue culture plates (Corning Costar Corp., Cambridge, MA) 24 h before transfection. Lipofectamine reagent (Gibco-BRL) was used to deliver DNA to the cells after a modified protocol from Gibco-BRL (Zhang et al., 1998b). Briefly, for each well, 1 μg of the purified plasmid DNA was added to 100 μl of Opti-MEM (Gibco-BRL) and 3.25 μl of lipid (2 mg/ml) was added to another 100 μl of media. The two solutions were then mixed and incubated for 30 min at room temperature. After incubation, 800 μl of Opti-MEM was added to the 200-μl mixture. The final volume of 1-ml mixture was applied to each well after rinsing the cells with 1 ml of the Opti-MEM. The cells were incubated for 18 h before the transfection mixture was removed by aspiration and replaced with standard complete growth medium.
Uptake Measurements.
In general, uptake studies were carried out 24 to 44 h post-transfection as described previously (Zhang et al., 1998b). Briefly, the growth medium was gently aspirated and each well was rinsed with 1 ml of PBS. To initiate uptake, 0.5 ml of PBS containing 10 μM [14C]TEA was added to each well. Inhibition and IC50 studies were carried out by adding various concentrations of unlabeled compounds to the reaction mixture. The uptake was carried out at room temperature for 20 min and stopped by aspiration of the uptake medium. The cell monolayers of each well were immediately washed with 2 ml of ice-cold PBS buffer once and 1 ml of the buffer twice. The cells were then solubilized with 1 ml 0.5% Triton X-100 and 0.5 ml of sample was assayed using liquid scintillation counting (Beckman Instruments, Palo Alto, CA).
In trans-stimulation studies, each well of cells was preincubated with either 0.5 ml of PBS buffer (control) or 0.5 ml of PBS buffer plus the indicated amount of unlabeled compound at 37°C for 1 h. Cells were then rinsed once with 2 ml of ice-cold PBS buffer and once with 1 ml of the buffer before the uptake studies were performed as described above.
The protein concentration was determined with the Bio-Rad protein assay kit (Bio-Rad, Hercules, CA), using BSA as the standard as described previously (Zhang et al., 1998b).
Efflux Studies.
Efflux studies were carried out 24 to 44 h post-transfection. The cells were washed with 1 ml of PBS before preincubation with 10 μM [14C]TEA for 1 h at 37°C. The cells were then washed twice with 1 ml of ice-cold PBS. For the time course we added only PBS to the wells, and determined the concentration of [14C]TEA in 0.5-ml samples of the media at various times. In thetrans-efflux studies, either 1 ml of PBS (control) or 1 ml of PBS with the indicated amount of unlabeled compound was added to each well of cells. After 10 min at room temperature, a 0.5-ml aliquot of incubation media was sampled and the concentration of [14C]TEA determined in the aliquot.
Partition Coefficient Determinations.
Octanol-water partition coefficients were determined from an n-octanol and water system at pH 7.4 (Neef and Meijer, 1984). Briefly, 5 ml of 4 mM procainamide, quinine, and quinidine water solutions were prepared. Then each water solution was mixed with 5 ml of n-octanol by vortexing. The mixture was rotated for 2 h at room temperature. The layers were separated by centrifugation at 2500 rpm for 15 min. Aliquots (100 μl) from each layer were diluted to 5 ml of water or octanol. The concentration ratio of octanol to water was determined as the ratio of UV absorbance from octanol to water solutions at λmax. The λmax value for procainamide is 278 nm; for quinine and quinidine the value is 330 nm.
Data Analysis.
In general, uptake values are expressed as mean ± S.E. or mean ± S.D. as indicated in the figurelegends. A minimum of two wells was used to generate a data point in each experiment. All experiments were repeated at least once on a different day using a different cell passage.
For IC50 studies, data were fit to the equation V = V0/[1+(I/IC50)n] where V is the uptake of [14C]TEA in the presence of inhibitor, V0 is the uptake of [14C]TEA in the absence of inhibitor, I is the inhibitor concentration, and n is the Hill coefficient. The KaleidaGraph fitting program (Abelbeck Software) was used to fit the data by nonlinear regression.
Statistical analysis was carried out by comparing the tested compounds with the controls from the same experiments using the unpaired Student’s t test (Primer of Biostatistics software, Version 3, written by Stanton A. Glantz, McGraw-Hill Companies, 1991). Results were considered statistically different with a probability ofp <.05. Analysis using one-way ANOVA produced pvalues that were not different from those obtained using Student’st test.
Materials.
HeLa cells and all of the media and buffers used to maintain the cells were obtained from the University of California San Francisco Cell Culture Facility. Original stocks of HeLa cells were from American Type Culture Collection (Rockville, MD). Lipofectamine and Opti-MEM were purchased from Gibco-BRL (Gaithersburg, MD). All chemicals were obtained from Sigma (St. Louis, MO) and Fisher (Pittsburgh, PA) or as indicated. [14C]TEA (55 mCi/mmol) was purchased from American Radiolabeled Chemicals, Inc. (St. Louis, MO). The n-TAA compounds (i.e., M-methyl, E-ethyl, Pr-propyl, Bu-butyl, Pe-pentyl, and H-hexyl) and n-octanol were purchased from Aldrich Chemicals (Milwaukee, WI) and tributylmethylammonium (TBuMA) was purchased from Fluka Chemical Corp., (Ronkonkoma, NY).
Results
Tissue Distribution of hOCT1.
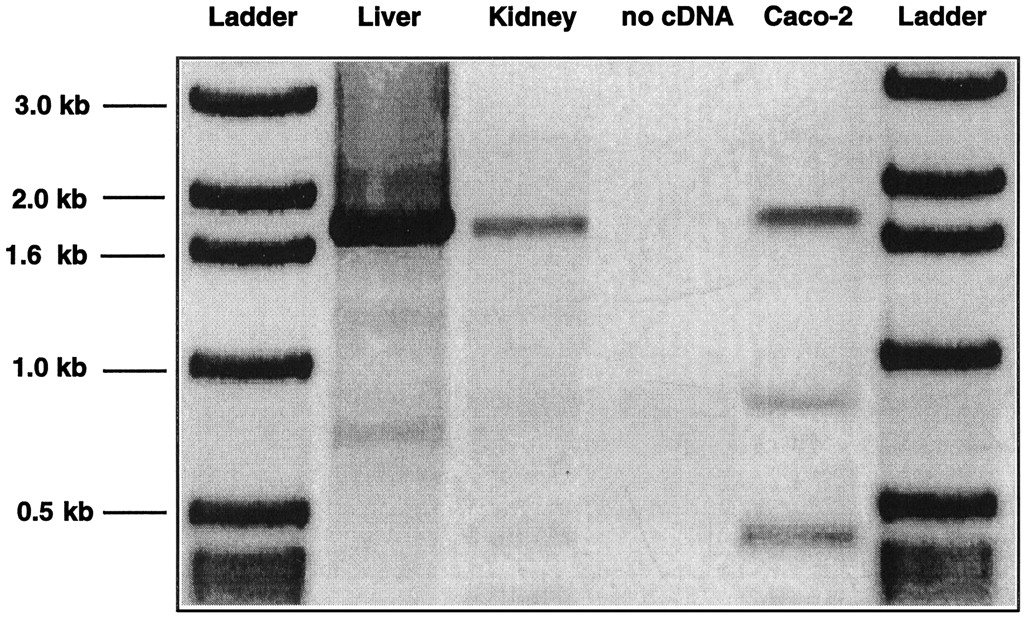
The tissue distribution of hOCT1 was determined by RT-PCR using primers derived from the hOCT1 cDNA sequence (Zhang et al., 1997b, Gorboulev et al., 1997). Bands corresponding to full-length hOCT1 were detected in human liver, kidney, small intestine (data not shown), and Caco-2 cells (Fig.1). The band from hOCT1 mRNA was strongest in the liver (Fig. 1), indicating that hOCT1 mRNA transcripts are expressed in abundance in human liver. Sequence analysis demonstrated that the sequences of these PCR products were identical with that of hOCT1.

RT-PCR analysis of hOCT1 mRNA transcript expression in various tissues. mRNA from human liver, kidney, and Caco-2 cells was subjected to RT-PCR using 5′ end primer and 3′ end primer (see Zhang et al., 1997b) to obtain a full-length DNA of the hOCT1 open reading frame. Water was used as the “no cDNA” control in PCR reactions. The PCR products were analyzed by 1% agarose gel and stained with ethidium bromide. The gel is shown as a negative version. A 1-kb DNA ladder (Gibco-BRL) was used to determine the band size.
Expression of hOCT1 in HeLa Cells.
Previous studies in this laboratory demonstrated that hOCT1 can be transiently expressed in HeLa cells (Zhang et al., 1998b). In this study, we halved the amount of DNA (i.e., 1 μg of DNA/well) used in transfection. With this modification, we used less lipid while maintaining a lipid/DNA ratio of 6.5:1. Under these conditions, we observed an enhancement in TEA uptake (data not shown) similar in magnitude to that observed in our previous studies (Zhang et al., 1998b).
Inhibition Studies.
cis-Inhibition studies were carried out to determine whethern-TAA compounds inhibit the uptake of [14C]TEA in hOCT1 DNA-transfected HeLa cells. Tetramethylammonium (TMA) (50 μM or 10 mM) did not inhibit [14C]TEA uptake (data not shown), whereas (50 μM) unlabeled tetrapropylammonium (TPrA), TBA, tetrapentylammonium (TPeA), and tetrahexylammonium (THA) significantly inhibited [14C]TEA uptake with various degrees of inhibition (Fig. 2).

Inhibition of [14C]TEA uptake byn-TAA compounds. The uptake of 10 μM [14C]TEA (at 20 min) was measured in pTargeT-hOCT1-transfected HeLa cells in the presence of 50 μM of the given compounds. Controls represent uptake of [14C]TEA in the pTargeT-hOCT1-transfected HeLa cells in the absence of inhibitors. Uptake of [14C]TEA in the empty vector-transfected cells in the absence of inhibitors is shown as well (indicated as Empty Vector). Data represent mean ± S.E. (n = 6–8) obtained from three to four separate experiments. *p <.05.
The concentration dependence of the various n-TAA compounds (e.g., TEA, TPrA, TBA, TPeA, and THA) in inhibiting [14C]TEA uptake was determined. To obtain IC50 values, data from multiple experiments were fit to an equation as described in Experimental Procedures. Figure 3 depicts a representative inhibition curve for TPeA. As shown in Table1, IC50 values decreased with increasing alkyl chain lengths (or molecular weights) from 260 ± 61 μM for TEA to 3.0 ± 1.6 μM for THA.

Concentration-dependent inhibition of [14C]TEA uptake by TPeA in HeLa cells transfected with pTargeT-hOCT1. Initial rates of transport were determined in the presence of various concentrations of TPeA. Data represent determinations from two experiments and were fitted by nonlinear regression as described in Experimental Procedures. The IC50 value of TPeA obtained from the fit was 8.6 ± 2.2 μM.
P values and IC50 values of n-TAA compounds
A semilogarithmic plot of IC50 values versus alkyl chain length (N) resulted in a straight line [i.e., log(IC50) = −0.490 · N + 3.46, where N = alkyl chain length, r2 = 0.990] (Fig. 4), suggesting that there is a good correlation between IC50 values and alkyl chain. Since alkyl chain length is related to the hydrophobicity of these compounds, we further determined whether there was a correlation between IC50 and partition coefficient (P) because hydrophobicity (or lipophilicity) can be expressed as the partition coefficient between octanol and an aqueous solution.

Relationship between IC50 values in inhibiting [14C]TEA uptake in HeLa cells expressing hOCT1 and alkyl chain length (N) of n-TAA compounds. Each IC50 value represents the mean ± S.E. fitted with data from two to four separate experiments by nonlinear regression as described in Experimental Procedures. The fitted equation is log(IC50) = −0.490 · N + 3.46,r2 = 0.990.
Previous studies have demonstrated that the molecular weights (MW) and partition coefficients (P, octanol/water) of “homologous” monoquaternary ammonium compounds are highly correlated (Neef and Meijer, 1984). Based on data in the literature (Neef and Meijer, 1984), we plotted MW versus log(P) (Fig. 5), and obtained a linear relationship, i.e., MW = 45.6 · log(P) + 247,r2 = 0.935. Using this equation, we derived P for the various n-TAA compounds used in this study (Table 1). As a consequence, a linear relationship was observed from a double-logarithmic plot of IC50 versus P, i.e., log(IC50) = −0.398 · log(P) + 1.44,r2 = 0.975 (Fig.6).

Relationship between MW of quaternary ammonium compounds and P values determined from octanol per water solution (P values cited from Neef and Meijer, 1984: TMA, .002;n-BuTMA, .0031; TEMA, .006; and TBuMA, .0744). The fitted equation is MW = 45.6 · log(P) + 247,r2 = 0.935.

Relationship between IC50 values in inhibiting [14C]TEA uptake in HeLa cells expressing hOCT1 and P values of n-TAA compounds estimated from Fig. 5. The fitted equation is log(IC50) = −0.393 · log(P) + 1.44, r2 = 0.974, when data for TBuMA (open circle) was included in the correlation curve. Data for procainamide, vecuronium, quinine, and quinidine are shown as triangles. When these data were included, the fitted equation is log(IC50) = −0.383 · log(P) + 1.49, r2 = 0.859.
To determine whether such a trend is also apparent for quaternaryn-alkylammonium compounds with different side chain lengths, we determined the IC50 value of TBuMA in inhibiting [14C]TEA uptake in HeLa cells expressing hOCT1. The IC50 for TBuMA (66 ± 24 μM) was plotted against P of TBuMA (0.0744). As shown in Fig. 6, TBuMA also fit well to the straight line (i.e., log(IC50) = −0.393 · log(P) + 1.43,r2 = 0.974 (Fig. 6).
We also examined the relationship of IC50 values and partition coefficients for nonaliphatic organic cations. Octanol-water partition coefficient values of procainamide, quinine, and quinidine were determined. As listed in Table2, P for procainamide is .009, for quinine is 2.32, and for quinidine is 2.45. When the nonaliphatic compounds (procainamide, vecuronium, quinine, and quinidine) were included, the relationship was log(IC50) = −0.383 · log(P) + 1.49, r2 = 0.859 (Fig. 6).
P values and IC50 values of non-aliphatic organic cations
trans-Stimulation Studies.
trans-Stimulation studies have been used previously to investigate whether an inhibitor might also be a substrate of hOCT1, i.e., translocated by hOCT1 (Zhang et al., 1998b). Preincubated hOCT1 DNA-transfected HeLa cells with various concentrations of unlabeled TEA exhibited trans-stimulation of [14C]TEA influx. We observed that the magnitude of the trans-stimulation effect increased with increasingtrans-TEA concentrations, and it was saturable at high concentrations. When we incubated the cells with a concentration representing 10 times the Km orKi value, i.e., 2 mM TEA, thetrans-stimulation effect was maximal (data not shown). Therefore, we preincubated cells with concentrations approximately 10 times the Km orKi value of unlabeled test compounds under the assumption that this condition would produce a maximumtrans-stimulation effect. As shown in Fig.7, after preincubating hOCT1 cDNA-transfected HeLa cells with TEA (2 mM), TPrA (1 mM), TBuMA (1 mM), or TBA (0.5 mM) for 1 h at 37°C, [14C]TEA uptake was significantly enhanced (p <.05) and the enhanced effect decreased with increasing alkyl chain length. This trans-stimulation effect was not observed in empty vector-transfected HeLa cells. Preincubating hOCT1 DNA-transfected cells with TMA (10 mM) or TPeA (0.2 mM) did not result in a significant change in [14C]TEA uptake, whereas preincubation of cells with 0.1 mM THA resulted in a significant decrease (apparent trans-inhibition) of [14C]TEA uptake (p <.05).

trans-Stimulation of [14C]TEA uptake in pTargeT-hOCT1 transfected HeLa cells. The uptake (at 20 min) of [14C]TEA (10 μM) was measured after a 60-min preincubation (followed by washing) of the hOCT1 DNA-transfected cells (dark bars) and empty vector-transfected cells (open bars) with PBS (control) or PBS containing 10 mM TMA, 2 mM TEA, 1 mM TPrA, 1 mM TBuMA, 0.5 mM TBA, 0.2 mM TPeA, or 0.1 mM THA, respectively, at 37°C. Data represent the mean ± S.E. (n = 4–10) obtained from two to five separate experiments. *p <.05.
trans-Stimulation studies were also performed with various organic cations, i.e., procainamide, vecuronium, quinine, and quinidine, which were previously shown to inhibit [14C]TEA uptake via hOCT1 (Zhang et al., 1998b). Figure 8 demonstrates that preincubating hOCT1 DNA-transfected cells with procainamide (1 mM)trans-stimulated TEA uptake, whereas preincubating cells with vecuronium (2 mM), quinine (0.2 mM), and quinidine (0.2 mM) resulted in a significant decrease (apparenttrans-inhibition) in [14C]TEA uptake (p <.05).

trans-Stimulation of [14C]TEA uptake in pTargeT-hOCT1-transfected HeLa cells. The uptake (at 20 min) of [14C]TEA (10 μM) was measured after a 60-min preincubation of the hOCT1 DNA-transfected cells (dark bars) and empty vector-transfected cells (open bars) with PBS (control) or PBS containing 2 mM TEA, 1 mM procainamide, 2 mM vecuronium, 0.2 mM quinine, or 0.2 mM quinidine, respectively, at 37°C. Data represent the mean ± S.E. (n = 4–8) obtained from two to four separate experiments. *p <.05.
Efflux Studies.
In further experiments we investigated the effect of the various n-TAA compounds on the efflux of [14C]TEA from cells preloaded with [14C]TEA. The efflux of [14C]TEA from HeLa cells transfected with hOCT1 plasmid DNA increased with time and was linear up to 40 min (data not shown). Therefore, further studies were carried out at 10 min. As shown in Fig. 9, after incubating the [14C]TEA preloaded cells with unlabeled TEA (2 mM), TPrA (1 mM), TBA (0.5 mM), and TBuMA (1 mM) for 10 min, efflux of [14C]TEA was significantly enhanced. Incubating with TMA (10 mM) and TPeA (0.2 mM) did not result in significant change in [14C]TEA efflux, whereas incubating the cells with THA (0.1 mM) resulted in a significant decrease of efflux (p <.05).

Efflux of [14C]TEA from hOCT1 DNA-transfected HeLa cells. Cells were preloaded by incubating with 10 μM [14C]TEA for 1 h at 37°C. The efflux of [14C]TEA was measured after incubating with either PBS buffer (control) or 10 mM TMA, 2 mM TEA, 1 mM TPrA, 1 mM TBuMA, 0.5 mM TBA, 0.2 mM TPeA, or 0.1 mM THA for 10 min. Empty vector efflux is subtracted from the efflux measured in hOCT1 transfected cells. Data represent the mean ± S.E. (n = 8) obtained from two separate experiments. *p <.05.
Discussion
Secretory transporters in the liver, kidney, and intestine play a role in the elimination of organic cations from the body (Turnheim and Lauterbach, 1977a,b; Miyamoto et al., 1988; Pritchard and Miller, 1993;Oude Elferink et al., 1995; Tomita et al., 1997; Zhang et al., 1998a). hOCT1 appears to be predominantly involved in the uptake of organic cations in the liver; however, data obtained in this study as well as in our previous study (Zhang et al., 1997b) demonstrate that hOCT1 mRNA transcripts are present, although in much lower abundance, in kidney and Caco-2 cells, a human colon carcinoma cell line (Fig. 1).
Functional studies of hOCT1 utilizing heterologous expression systems (e.g., X. laevis oocytes and transfected HeLa cells) have demonstrated that various organic cations as well as some neutral and anionic compounds inhibit the transport of model organic cations by hOCT1 (Zhang et al., 1997b; 1998b). The goal of the current study was to use a series of n-TAA compounds to systematically determine the effect of hydrophobicity on inhibition potencies of compounds in interacting with hOCT1. Furthermore, the effect of hydrophobicity on the rate of transport (e.g., influx and efflux) by hOCT1 was determined.
Initially, a good correlation between IC50 values and alkyl chain length was found. Namely, we observed thatn-TAA compounds with the longer alkyl chain lengths were the more potent inhibitors of hOCT1. This observation is consistent with what is generally found for these compounds in inhibiting organic cation transport (Dantzler et al., 1991; Groves et al., 1994; Ullrich et al., 1991; Wright et al., 1995; Wright and Wunz, 1998). In the literature, a linear relationship between P values of the quaternary ammonium cations (including n-TAAs) and MWs has been reported (Neef and Meijer, 1984). We used this relationship to estimate the P values of several n-TAAs (Table 1). A good correlation between IC50 and P in a double-logarithmic plot was generated for homologous n-TAAs (i.e., log(IC50) = −0.398 · log(P) + 1.44,r2 = 0.975 (Fig. 6). The data indicate that for inhibition, binding affinity is increased with increasing hydrophobicity. This correlation also applies to TBuMA for which the four alkyl chains are not of equal length (Fig. 6). Althoughr2 was slightly reduced from 0.974 to 0.859 when data for nonaliphatic compounds were included in the same plot (Fig. 6), a good relationship between IC50and P value is still apparent; this suggests that a compound’s hydrophobicity is a major determinant of its potency of interaction with hOCT1.
Inhibition does not necessarily imply that a compound is also translocated by a transporter. Since radiolabeled n-TAA compounds are not available except for TEA, we usedtrans-stimulation studies to investigate whether then-TAA compounds might also be substrates of hOCT1.trans-Stimulation is a frequently used method to test whether two molecules share a common transport pathway (Holohan and Ross, 1980; Miyamoto et al., 1988; Dantzler et al., 1991). If the presence of the test compound on the opposite side (trans) of the membrane results in an enhanced flux of the radiolabeled probe, the test compound is considered to be a substrate for the same transporter as the probe, and vice versa. However if a compound does not trans-stimulate or does trans-inhibit it may or may not be a substrate (Busch et al., 1998). Tracer flux experiments are required to determine whether such compounds are in fact substrates. trans-Stimulation has been used previously in the hOCT1 DNA-transfected cell line to investigate whether an inhibitor might also be a substrate (Zhang et al., 1998b).
TMA did not trans-stimulate TEA influx or efflux; this is consistent with our inhibition study suggesting that TMA does not interact with hOCT1. For all other n-TAA compounds, we observed that compounds with shorter alkyl chain lengths (or smaller P, i.e., less lipophilic), TEA, TPrA, TBuMA, and TBA, producedtrans-stimulation effects. However, the magnitude of the effect decreased with increasing alkyl chain length, in the [14C]TEA influx experiments, suggesting that the transport rate was decreased (Fig. 7). In contrast, compounds with a longer alkyl chain length (or larger P, i.e., more lipophilic), TPeA and THA, did not demonstrate a trans-stimulation effect. Moreover, THA demonstrated an apparent trans-inhibition effect, suggesting that the transporter loaded with THA has a slower turnover rate than that of the unloaded transporter. The compounds which trans-stimulated [14C]TEA influx (Fig. 7) were also shown to trans-stimulate [14C]TEA efflux from cells preloaded with [14C]TEA (Fig. 9). However, notable differences in the trans-stimulation effect of TBuMA on [14C]TEA influx versus efflux were observed, suggesting that hOCT1 may have asymmetrical binding sites.
An alternative explanation of the apparent trans-stimulation effect could be that n-TAA compounds might interact with a site not related directly to the transporter (e.g., an ion channel in the cells) and alter the membrane potential. Since hOCT1 is a potential-dependent transporter, an effect on membrane potential would alter [14C]TEA influx or efflux. Because of the low activity of the expressed transporter in the transfected cells, we were unable to perform studies under voltage-clamped conditions. Therefore, we cannot exclude indirect effects of the n-TAA compounds on membrane potential. However, unpublished data from this laboratory indicate that n-TAA compoundstrans-stimulate [14C]TEA uptake by hOCT1 and by rOCT1 differently. If the trans-stimulation effects of the n-TAA compounds were solely due to indirect effects on membrane potential, the compounds should have produced similar trans-stimulation of [14C]TEA uptake in cells transfected with either hOCT1 or rOCT1.
Previously, it was shown that increasing lipophilicity is associated with an increase in hepatic clearance of various compounds in the rat (Neef and Meijer, 1984; Proost et al., 1997). These results are in contrast to our data, which suggest that with increasing lipophilicity, the efflux via hOCT1 decreases (Fig. 7). The discrepancies between our data and the previous studies in the intact liver may be explained in several ways. First, diffusional pathways as well as multiple transporters on both the canalicular and sinusoidal membrane contribute to hepatobiliary secretion in the intact organ or animal. Second, hOCT1 is a transporter cloned from human liver; species differences between the structure activity relationships of the rat and human transporters may be present. Previously, notable species differences in the effect of hydrophobicity on transport of organic cations in rat and rabbit kidney preparations were observed (Ullrich et al., 1991; Groves et al., 1994).
In summary, our observations suggest that the longer the alkyl chain length (i.e., the more hydrophobic and bulkier), the higher the affinity of the tetraalkylammonium compounds for hOCT1, but the slower the rate of transport (i.e., poorer substrate) by hOCT1. hOCT1 may represent a human liver transporter which accepts smaller organic cations as its substrates. The correlation observed between IC50 and P values could be used to estimate the IC50 values of various n-alkylammonium compounds in interacting with hOCT1. A balance between hydrophobic and hydrophilic properties is required for efficient transport of an organic cation by hOCT1.
Footnotes
-
Send reprint requests to: Kathleen M. Giacomini, Ph.D., Department of Biopharmaceutical Sciences, University of California, San Francisco, 513 Parnassus, S-926, San Francisco, CA 94143-1936. E-mail:kmg{at}itsa.ucsf.edu
-
1 This study was supported by National Institutes of Health Grant GM-57656. L.Z. was supported in part by the University of California San Francisco Chancellor’s Graduate Research Fellowship.
- Abbreviations:
- hOCT1
- human organic cation transporter
- TEA
- tetraethylammonium
- TPrA
- tetrapropylammonium
- TBuMA
- tributylmethylammonium
- TPeA
- tetrapentylammonium
- THA
- tetrahexylammonium
- n-TAA
- n-tetraalkylammonium
- RT-PCR
- reverse transcription-polymerase chain reaction
- MW
- molecular weight
- Received June 24, 1998.
- Accepted October 19, 1998.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}