


Abstract
Improved treatment of Alzheimer disease (AD) is a significant unmet medical need that is becoming even more critical given the rise in the number of patients and the substantial economic burden. The current standards of care, acetylcholinesterase inhibitors (AChEIs), are hindered by gastrointestinal side effects owing to their nonselective activation of muscarinic and nicotinic receptors. Recently, the highly selective M1 positive allosteric modulator PQCA (1-((4-cyano-4-(pyridine-2-yl)piperidin-1-yl)methyl-4-oxo-4 H-quinolizine-3-carboxylic acid) has been demonstrated to improve cognition in a variety of rodent and nonhuman primate cognition models without producing significant gastrointestinal side effects. Here we describe the effect of PQCA and the AChEI donepezil on two clinically relevant and highly translatable touchscreen cognition tasks in nonhuman primates: paired-associates learning (PAL) and the continuous-performance task (CPT). Blockade of muscarinic signaling by scopolamine produced significant impairments in both PAL and CPT. PQCA and donepezil attenuated the scopolamine deficits in both tasks, and the action of these two compounds was similar in magnitude. In addition, the combination of subeffective doses of PQCA and donepezil enhanced PAL performance. These results further suggest that M1-positive allosteric modulators, either as monotherapy or as an add-on to current standards of care, have potential to reduce the cognitive deficits associated with AD.
Introduction
Alzheimer's disease (AD) is a profoundly debilitating neurodegenerative disorder characterized by neuronal cell dysfunction and death, particularly in the cholinergic neurons of the basal forebrain (Bartus et al., 1982; Schliebs and Arendt, 2006). Most currently approved symptomatic treatments for AD promote cholinergic function; four of the five drugs approved for symptomatic AD treatment are acetylcholinesterase inhibitors (AChEIs). Unfortunately, AChEIs provide relatively modest cognitive improvement and tend to produce significant gastrointestinal side effects (Raina et al., 2008). As a result of these limitations and the severity of the disease, novel treatments for AD are a significant unmet medical need.
The M1 muscarinic receptor has gained interest as a target for novel treatments because of its high level of expression in brain areas associated with cognitive processing and its limited expression in the periphery (Levey, 1993; Volpicelli and Levey, 2004). Furthermore, M1 knockout mice show a variety of cognitive deficits (Miyakawa et al., 2001; Anagnostaras et al., 2003), and compounds that activate the M1 receptor have demonstrated procognitive effects preclinically (Sedman et al., 1995; Hatcher et al., 1998; Wienrich et al., 2001).
Orthosteric M1 agonists have been developed and, in some cases, tested clinically (Sedman et al., 1995; Hatcher et al., 1998; Wienrich et al., 2001); however, the development of these compounds has been unsuccessful as a result of issues, including insufficient selectivity, gastrointestinal side effects, and modest efficacy potentially because of the use of animal models that lack predictive validity (Fisher et al., 2003; Schenk et al., 2005; Cummings, 2008; Lindner et al., 2008; Conn et al., 2009). It is unclear which of these factors has resulted in the lack of clinical success.
To overcome the issue of selectivity and gastrointestinal tolerability, we have developed M1 positive allosteric modulators (PAMs) that are potent and highly selective (Ma et al., 2009). For example PQCA (1-((4-cyano-4-(pyridine-2-yl)piperidin-1-yl)methyl-4-oxo-4 H-quinolizine-3-carboxylic acid) has an EC50 value of 49 and 135 nM on rhesus and human M1 receptor, respectively, and is inactive up to 30 μM versus the other muscarinic receptors (Kuduk et al., 2011). Previously we have demonstrated that PQCA enhances cognition across preclinical species and cognitive domains while providing a significantly improved therapeutic window in terms of GI effects, relative to the AChEI donepezil and the nonselective M1 orthosteric agonists xanomeline (Uslaner et al., 2013; Vardigan et al., 2014).
In this study, we aimed to provide more convincing translatable cognition data by extending our characterization of PQCA to two rhesus macaque touchscreen cognition tests, paired-associates learning (PAL), and the continuous-performance task (CPT). These tests are particularly valuable preclinically because nearly identical tests can be administered to humans. Results from such efforts show a high degree of cross-species translatability (Sahakian and Owen, 1992; Robbins et al., 1994; Fray and Robbins, 1996; Fray et al., 1996; Taffe et al., 1999; de Jager et al., 2002; Taffe et al., 2002; Blackwell et al., 2004; Buccafusco, 2006; Egerhazi et al., 2007; Lindner et al., 2008).
In the current set of studies, we first characterized donepezil in PAL and CPT in scopolamine-impaired rhesus monkeys. As far as we know, this is the first time an AChEi has been tested in these assays in nonhuman primates and so represents the first pharmacologic validation of these potentially translatable assays. Second, we characterized the effects of PQCA in both PAL and CPT, representing the first M1 PAM characterized in these assays. Finally, we examined whether subeffective doses of PQCA and donepezil could be combined to produce effects in PAL, which could suggest the potential for additivity in the clinic.
Materials and Methods
Animals.
Eighteen adult male rhesus monkeys (Macaca mulatta) were trained on either the PAL (n = 10) or CPT (n = 8) tasks. Sample size was determined using a power analysis based on performance during training and an expected effect of ≥20% on PAL accuracy and CPT hit rate. To achieve >80% power, the sample size was determined to be seven or more animals. Many of these animals had received test compounds in the past; however, all animals received a 2-week washout before the experiments described here, which is many times longer than the half-lives of the compounds previously tested.
Animals were fed their full allotment of dry food (Purina High Protein Monkey Diet no. 5045), a multivitamin, and fresh fruits and vegetables after cognition testing at ∼1500 hours. The amount of dry food for each animal was manipulated to maintain engagement in cognitive testing studies. Dietary restrictions were approved by the veterinary staff, and body condition assessments were made every other week. Body scores were maintained at or above 2 on the Clingerman and Summers (2005) scale. Water was available ad libitum. Temperature and relative humidity were maintained at 21–24°C and 50%–55%, respectively. All experimental protocols described in this study were approved by the Merck and Co., Inc., Institutional Animal Care and Use Committee and conducted in accordance with the Guide for Care and Use of Laboratory Animals. All efforts were made to minimize animal suffering, to reduce the number of animals used, and to use alternatives to in vivo methods where possible.
Apparatus.
Cognitive testing for each animal took place inside a sound attenuated testing chamber (35 × 35 × 49.5 inches) in a room outside of the housing colony. Each chamber was equipped with a touch-screen computer system running the Cambridge Neuropsychological Test Automated Battery software (Cambridge Cognition, Cambridge, UK) designed for use with nonhuman primates. A pellet dispenser provided a food reward (100-mg flavored pellets; Bio-Serv, Flemington, NJ) to reinforce participation and correct responses. Testing chambers were ventilated and also included a stimulus light, house light, and speakers for white noise and auditory stimuli. Monkeys were unrestrained and free to move around the test chamber during testing. All animals were monitored via live video feeds streamed from cameras mounted inside the test chambers.
Paired-Associates Learning.
Ten single- or pair-housed male rhesus monkeys, ages 6-10 years old (5.2-10.5 kg), were tested on the PAL task, a task dependent on both frontal cortical and temporal-hippocampal systems (Gould et al., 2003, 2006; Meltzer and Constable, 2005 ). The PAL task required the subject to learn to associate specific visual stimuli with specific locations on a trial-by-trial basis, as previously described (Taffe et al., 2002). The stimuli for the PAL task consisted of ∼70 distinct colored shapes or patterns. In each test session, animals were presented with 75 trials, which comprised three different trial types: 2-2, 2 samples in two locations; 3-3, three samples in three locations; and 4-4, four samples in four locations. At the beginning of a trial, one sample stimulus was presented in one of the four corners of the touchscreen. The animal had to acknowledge the stimulus by touching it, and no additional stimuli were presented until the animal responded. Once the animal responded, the second sample was presented in its own unique location until the animal responded. Depending on the trial type, two, three, or four sample stimuli were presented sequentially in unique locations. Sample presentations were separated by a 0.5-second blank screen. After all the sample stimuli were presented, followed by a 1-second blank screen, the choice phase began. In the choice phase, one of the sample stimuli was presented simultaneously in each sample phase location. The animal was then required to touch the correct sample location for that particular stimulus. After a 0.5-second blank screen, the next stimulus was presented in each of the sample locations, and the animal was again required to choose the correct location for that particular stimulus. The animal had to successfully complete a trial by responding correctly to all choices to obtain a food reward. Food reward was accompanied by a 1000-Hz, 85-d B tone and illumination of the food magazine. If the animal made an error, the trial ended, accompanied by a 40-Hz, 85-d B tone, and the animal received a short timeout (5 s) before the next trial began. Animals were trained on PAL until performance on the three trial types was stable at >80% correct on 2-2, 40%–60% correct on 3-3, and <40% correct on 4-4. The percent correct measure was calculated for each trial type as the number of successfully completed trials divided by the number of trials presented. Figure 1 further illustrates the task.

Diagram of paired-associates learning-task trials. Two to four sample stimuli are presented sequentially during the sample phase of each trial. After the animal touches each sample stimulus, the stimuli are redisplayed in all the locations used during the sample phase. During this choice phase, the animal must remember the stimulus-location pairing for each stimulus. The order of the stimuli in the choice phase is randomized. A trial is successfully completed if the animal correctly identifies the sample location of each stimulus.
Subjects received four test sessions weekly (Mondays, Tuesdays, Thursdays, and Fridays). Mondays and Thursdays were typically baseline sessions. Tuesdays and Fridays were possible drug treatment days, which allowed at least 3 days for compound washout. Drug administration occurred if each subject’s performance on vehicle (i.m. saline in the case of scopolamine, p.o. saline for donepezil, and p.o. 20% vitamin E TPGS for PQCA) was stable. Animals were considered stable if their standard error for the percent of correct trials was less than 7.5 for each trial type over the previous three vehicle sessions. Once stability was attained, animals began scopolamine (Sigma Aldrich, 5–30 μg/kg administered i.m.) titration. Scopolamine dosing was individualized for each subject, as behavioral sensitivity to impairment agents in monkeys varies greatly from subject to subject (Cannon et al., 2013). Each subject’s scopolamine dose was titrated to identify a dose that produced a significant decrease in trial success while still completing at least 80% of trials. The scopolamine impairment was also assessed for stability over multiple sessions using the preceding criteria. Once stable performance for both vehicle and scopolamine was achieved, donepezil (0.3–3.0 mg/kg, administered p.o.; Sequoia Research Products, Pangbourne, UK), validation began. Using a Latin square study design, scopolamine (or vehicle) and donepezil (or vehicle) were administered 30 minutes and 4–5 hours before testing, respectively. Finally, after donepezil validation and after at least a 1-week washout period, characterization of PQCA (3, 10, or 30 mg/kg, administered p.o.) began. Doses of PQCA were based on Uslaner et al. (2013), which demonstrated that doses within this range reliably attenuated a scopolamine-induced deficit in other nonhuman primate (NHP) cognition tasks. Using a Latin square study design, scopolamine (or vehicle) and PQCA (or vehicle) were administered 30 minutes and 4 hours before testing, respectively. Finally, a combination study was performed using subeffective doses of donepezil and PQCA to assess possible additivity or synergy between the compounds. Vehicle performance was monitored and remained stable throughout the drug studies.
Continuous Performance Tasks.
Eight single- or pair-housed male rhesus monkeys, aged 6–10 years (4.9–6.8 kg), were tested on the CPT task; a test of sustained attention and impulsivity. This task was a modified version of the CPT previously described by Zeamer et al. (2011). Each CPT session consisted of rapidly presented identically shaped stimuli, which varied only by color [white (RGB: 255 255 255), light yellow (RGB: 255 255 20–100), and yellow (RGB: 255 255 0)]. Yellow was set as the target stimulus. Touching the target was rewarded with a food pellet, accompanied by a 1000-Hz tone. These responses were recorded as hits. Failing to touch the target stimulus was recorded as a miss. The white stimulus was used as an easy distractor to confirm that each subject was responding selectively to the target stimulus; this allowed for exclusion of animals who responded indiscriminately to all stimuli. The light yellow stimulus was used as a difficult distractor. The RGB value of the light yellow color was titrated for each animal to closely resemble the target and to establish a baseline level of false alarms. Because drug treatments have the potential to impact general levels of responding, these incorrect responses provided a way to dissociate a decrease in accuracy from a general decrease in responsivity. Touching either distractor stimulus resulted in a 10-second timeout during which the stimulus light was extinguished and a 40-Hz tone was presented. These responses were recorded as false alarms. Correctly rejecting either type of distractor did not result in food reward but was accompanied by a 1000-Hz tone. Animals received 100 trials per session, with 60% of the stimuli presented as targets, and the other 40% split evenly between easy and difficult distractors. Stimuli were presented for 1 second with a 2-second intertrial interval. The measures recorded for each testing session were (1) hits—the number of correct touches to the target, (2) misses—the number of missed targets, (3) false alarms—the number of incorrect touches to distractors, (4) correct rejections—the number of correctly avoided distractors, and (5) reaction time—the time between appearance and response (hit or false alarm). Animals were trained until they reached an asymptotic performance level of ∼80% correct responses to hits and ∼30% false alarms to the difficult distractor.
In addition to CPT, a color-discrimination (CD) task was given to control for potential drug effects on color vision. The CD task used the same stimuli as CPT. Impairment in both CD and CPT may reflect impairment in color vision, whereas an impairment in only CPT performance with a sparing of CD accuracy suggests an effect on attention. In this task, the target yellow stimulus was presented concurrently with either the easy or difficult distractor. Animals received 40 randomly presented trials immediately after the CPT task was completed; thus, all compound treatments were identical to the CPT task as described herein. Animals completed CPT/CD testing in 15–20 minutes. Considering the relatively long half-lives of the drugs described in these studies, we expect plasma drug concentrations to be similar between the CPT and CD tasks. The CD trials used the same timing parameters as CPT except that each discrimination trial was presented for up to 5 seconds to reduce the attentional strain. The measures recorded for each testing session were (1) correct responses and (2) omissions. Figure 2 further illustrates CPT and the CD task.
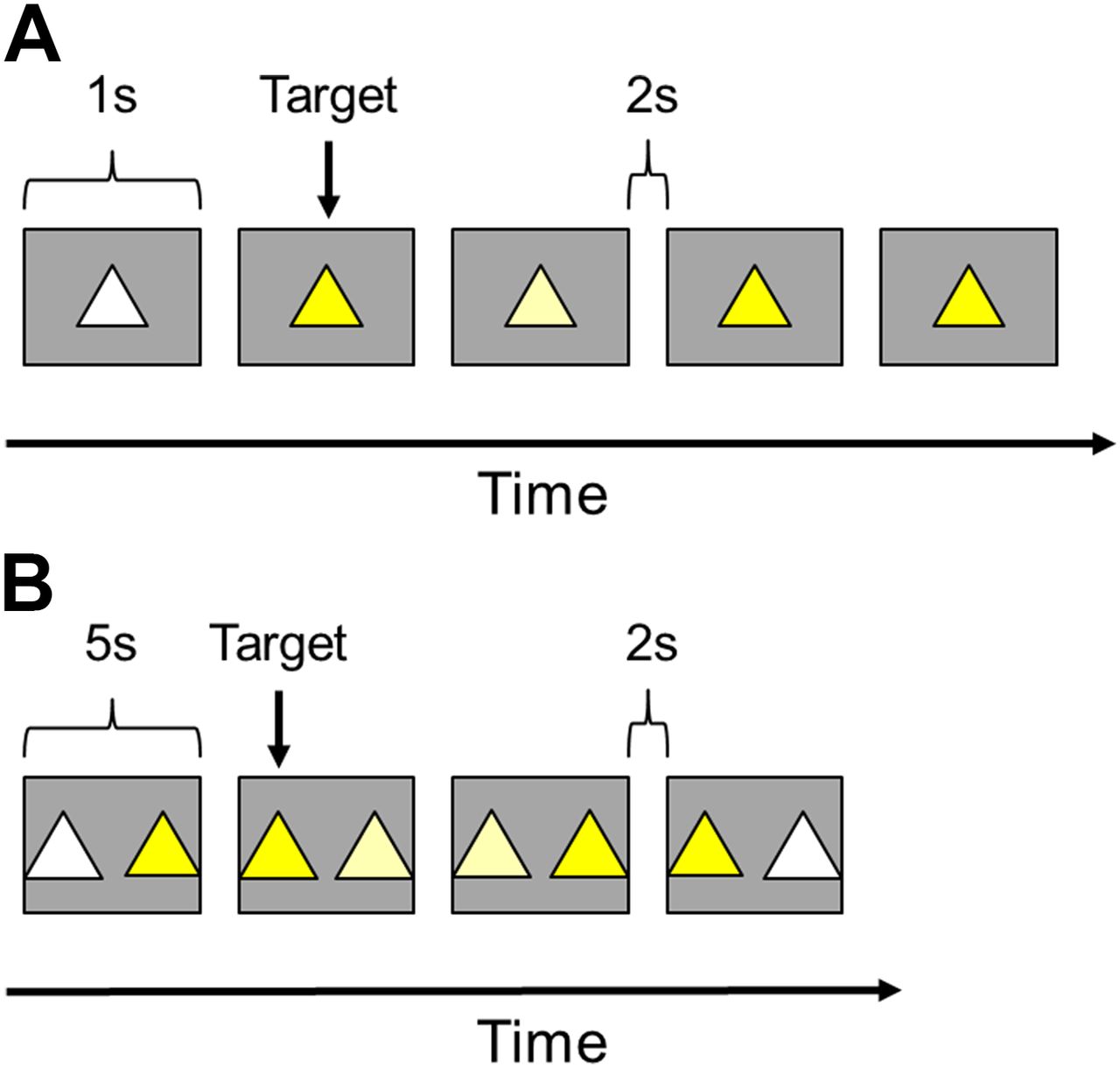
Diagram of trials for the CPT and the CD task. In CPT (A), identically shaped stimuli, which varied by color, were presented rapidly in the center of the screen. Animals were trained to respond to yellow stimuli and ignore white and light yellow distractors. Touches to the yellow stimulus were recoded as hits, and touches to the distractors were recorded as false alarms. The RGB value of the light yellow color was titrated for each animal to establish a baseline level of false alarms, which provided a way to separate a decrease in accuracy from a general decrease in responsivity. Animals received 100 trials per session with 60% of the stimuli presented as targets, and the other 40% split evenly between easy and difficult distractors. Stimuli were presented for 1 second with a 2-second intertrial interval. In CD (B), one target stimulus and one type of distractor were presented concurrently. Animals were trained to respond to the yellow stimuli and ignore white and light yellow distractors. Animals received 40 trials immediately after completion of each CPT session. Stimuli were presented for 5 seconds with a 2-second intertrial interval.
Subjects received four test sessions weekly (Mondays, Tuesdays, Thursdays, and Fridays), with drug administration typically occurring on Tuesdays and Fridays. Drug administration occurred only if each subject’s vehicle performance (i.m. saline for all compounds) was stable, as described already. For CPT, subjects’ scopolamine doses were titrated to identify a dose that reliably decreased hits (>20%) with minimal effect on false alarms. Once a dose that met these parameters was identified, it was repeated at least once. After stable performance for both vehicle and scopolamine was achieved, donepezil (0.1–0.25 mg/kg, i.m.) validation and PQCA (0.3–1.0 mg/kg, i.m.) characterization began. The CPT animals were dosed i.m. because of scheduling issues that prevented us from testing CPT at pretreatment times required for p.o. dosing with these compounds (Tmax ∼4 hours when given p.o.). As shown in Table 1, we chose i.m. doses to match plasma exposures from our previous studies (Uslaner et al., 2013; Vardigan et al., 2014), in which compounds were given p.o. Using a Latin square study design, scopolamine (or saline) and donepezil (or saline) were administered 60 minutes and 30 minutes before testing, respectively. In the PQCA study, scopolamine (or saline) and PQCA (or saline) were administered 60 minutes and 30 minutes before testing, respectively.
Comparison of drug plasma concentrations between dosing routes
Data are mean ± S.D., n = 3/group.
Data Analysis.
Each dependent measure was analyzed via one-way repeated measures analysis of variance (ANOVA).
Within-subjects t tests were performed to compare differences between scopolamine and all other treatments. In addition, to characterizing sustained attention, CPT data were divided into the first and second half of each session and analyzed using a two-way repeated measures ANOVA, with treatment and testing phase as within subject factors. Finally, a two-way repeated measures ANOVA was used to assess CD with treatment and distractor type as within subject factors.
Results
Paired-Associates Learning.
The effect of scopolamine and donepezil on paired-associate learning performance is presented in Fig. 3. A one-way repeated measures ANOVA revealed a main effect of treatment on 2-2 (F(4, 36) = 4.136, P = 0.007), 3-3 (F(4, 36) = 15.937, P < 0.001), and 4-4 (F(4, 36) = 5.506, P = 0.001) trial types. Post hoc analysis demonstrated that scopolamine impaired performance relative to vehicle on all three trial types. For 3-3 trials, 1.0 and 3.0 mg/kg donepezil significantly attenuated the scopolamine effect (P = 0.047 and P = 0.001, respectively); 3.0 mg/kg donepezil showed a trend to reduce the scopolamine effect (P = 0.067) on 4-4 trials.

Donepezil improves accuracy on scopolamine-impaired paired-associates learning in rhesus monkeys. Data shown are percent successful trials for each trial type. *P < 0.05, **P < 0.01, and ***P < 0.001 compared with scopolamine treatment (n = 10).
Figure 4 shows the effect of PQCA on PAL. One-way repeated measures ANOVA revealed a main effect of treatment on 2-2 (F(4, 32) = 11.012, P < 0.001), 3-3 (F(4, 32) = 12.165, P < 0.001), and 4-4 (F(4, 32) = 4.607, P = 0.005) trial types. Similarly to the donepezil study, scopolamine significantly impaired performance for all three trial types; 10 and 30 mg/kg PQCA attenuated the scopolamine effect on 2-2 (P = 0.003 and P = 0.05, respectively) and 3-3 trials (P = 0.009 and P = 0.015, respectively).

PQCA improves accuracy on scopolamine impaired paired-associates learning in rhesus monkeys. Data shown are percent successful trials for each trial type. *P < 0.05, **P < 0.01, and ***P < 0.001 compared with scopolamine treatment (n = 10).
The results from a combination study characterizing ineffective doses of donepezil (0.3 mg/kg) and PQCA (3.0 mg/kg) are shown in Fig. 5. A repeated-measures ANOVA revealed a main effect of treatment of 2-2 (F(2, 14) = 10.171, P = 0.002), 3-3 (F(2, 14) = 16.355, P < 0.001), and 4-4 (F(2, 14) = 8.046, P = 0.005) trial types. Again, scopolamine produced a robust and significant effect for all trial types. The combination of donepezil and PQCA significantly attenuated the scopolamine effect for 2-2 trials (P = 0.05) and trended toward reducing the scopolamine effect for both 3-3 (P = 0.094) and 4-4 (P = 0.073) trial types. When given alone, neither of these doses of donepezil or PQCA attenuated the scopolamine impairment.
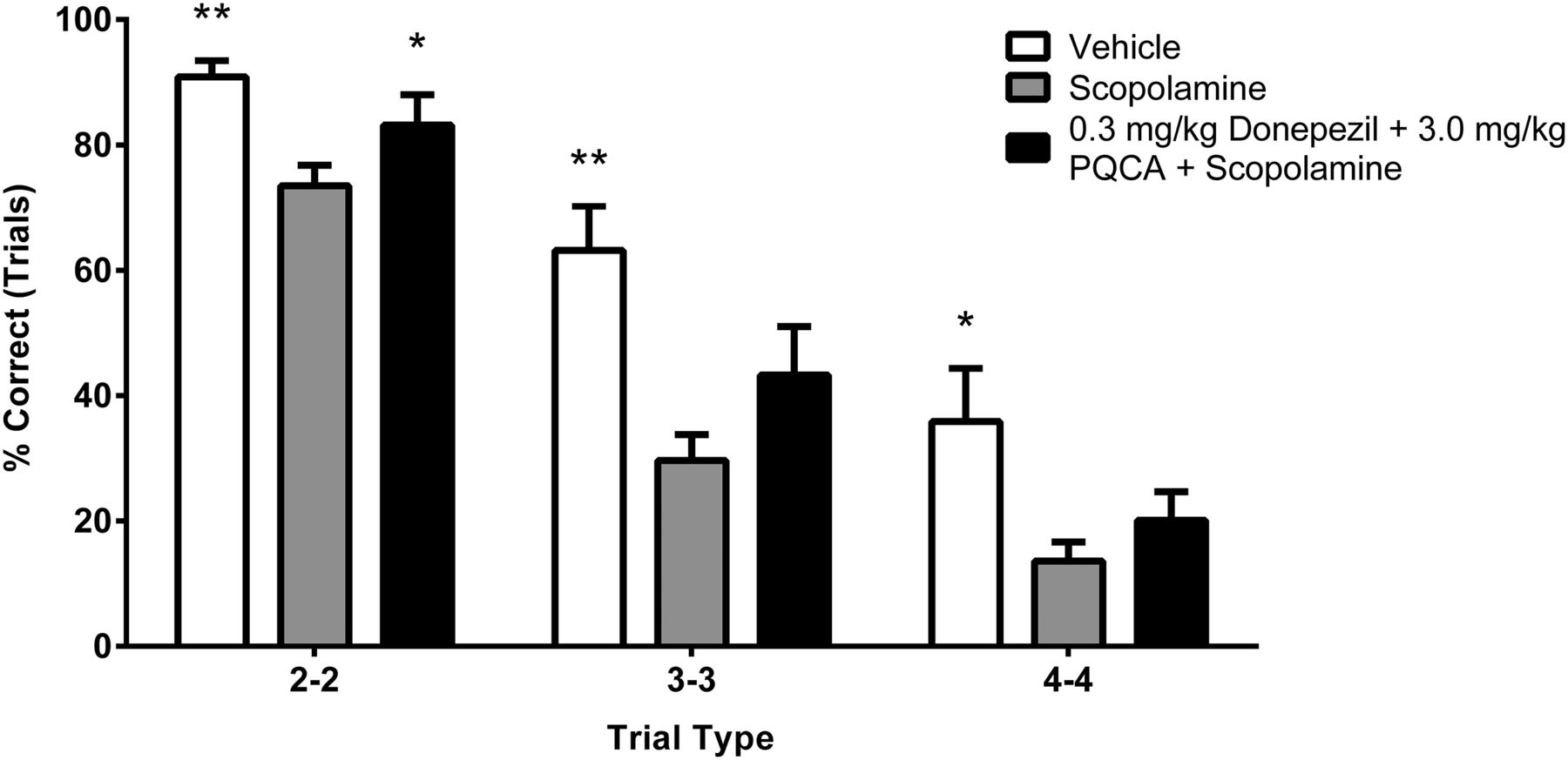
The combination of subeffective doses of donepezil and PQCA improves accuracy on scopolamine-impaired paired-associates learning in rhesus monkeys. Data shown are percent successful trials for each trial type. *P < 0.05 and **P < 0.01 compared with scopolamine treatment (n = 10).
Continuous-Performance Test.
Table 2 and Fig. 6 show the effect of scopolamine and donepezil on the CPT. A one-way repeated-measures ANOVA revealed a main effect of treatment on hit rate (F(3, 21) = 7.843, P = 0.001) (Fig. 6). Scopolamine treatment significantly impaired hit rate (P = 0.001), and the high dose of donepezil (0.25 mg/kg) attenuated the scopolamine effect (P = 0.018). Difficult distractors produced significantly more false alarms than easy distractors (F(1, 7) = 41.924, P < 0.001), but neither type of false alarm was impacted by compound treatment (difficult: F(3, 21) = 1.645, P = 0.209; easy: F(3, 21) = 0.982, P = 0.420). Reaction time for targets was also affected by compound treatment (F(3, 21) = 6.559, P = 0.003). Scopolamine significantly increased reaction time (P = 0.021), and 0.25 mg/kg donepezil reduced the scopolamine effect (P = 0.042). A two-way repeated measures ANOVA revealed main effects of testing phase (F(1, 7) = 21.595, P = 0.002) and treatment (F(3, 21) = 8.433, P = 0.001) but not a phase x treatment interaction (F(3, 21) = 0.888, P = 0.463) for the number of missed targets. The number of missed targets significantly increased during the second half of testing (P = 0.002), suggesting difficulty sustaining attention. A one-way ANOVA on the first half of testing revealed a main effect of treatment on missed targets (F(3, 21) = 4.799, P = 0.011). Scopolamine significantly increased missed targets during the first phase of testing (P = 0.004), and the high dose of donepezil failed to significantly impact the effect of scopolamine, although a trend was noted (P = 0.095). A one-way ANOVA on the second half of testing revealed a main effect of treatment on missed targets (F(3, 21) = 7.840, P = 0.001). Scopolamine significantly increased missed targets during the second half of testing (P = 0.002), and the high dose of donepezil significantly reduced the scopolamine effect (P = 0.018).
Effect of donepezil on the scopolamine-impaired CPT rhesus monkeys
Data are mean ± S.E.M, n = 8.

Donepezil improves accuracy on the scopolamine-impaired continuous-performance task in rhesus monkeys. Scopolamine significantly impaired the hit rate, and donepezil (0.25 mg/kg) attenuated this impairment. *P < 0.05 and ***P < 0.001 compared with scopolamine treatment (n = 8).
In the CD control task, total accuracy (F(3, 21) = 0.609, P = 0.616) and total omissions (F(3,21) = 2.215, P = 0.116) were not affected by scopolamine or donepezil dosing. Furthermore, a two-way repeated-measures ANOVA for accuracy revealed a main effect of distractor type (F(1, 7) = 15.257, P = 0.006), with difficult distractors producing more errors, but no main effect of treatment (F(3, 21) = 0.747, P = 0.536). The treatment x distractor type interaction trended toward significance (F(3, 21) = 2.810, P = 0.064). In addition, the two-way repeated measures ANOVA for omissions was not significant for distractor type (F(1, 7) = 0.089, P = 0.775), treatment (F(3, 21) = 1.997, P = 0.145), or the interaction (F(3, 21) = 0.529, P = 0.667).
Table 3 and Fig. 7 show the effect of PQCA on scopolamine-impaired CPT. A repeated-measures ANOVA revealed a main effect of treatment on the hit rate (F(3, 21) = 17.157, P < 0.001). Scopolamine significantly impaired the hit rate (P < 0.001), and 0.3 and 1.0 mg/kg PQCA significantly attenuated the scopolamine effect (P = 0.021 and P = 0.041, respectively) (Fig. 7). The false-alarm rate was not affected by treatment of difficult (F(3, 21) = 0.983, P = 0.420) or easy (F(3, 21) = 0.011, P = 0.998) distractors. Difficult distractors produced significantly more false alarms than did easy distractors (F(1, 7) = 98.882, P < 0.001). Reaction time for targets was also influenced by treatment (F(3, 21) = 9.917, P < 0.001). Scopolamine significantly increased reaction time (P = 0.012), but neither dose of PQCA mitigated that effect. A two-way repeated-measures ANOVA revealed a significant main effect of testing phase (F(1, 7) = 12.976, P = 0.009) and treatment (F(3, 21) = 17.100, P < 0.001) and a phase x treatment interaction (F(3, 21) = 6.446, P = 0.003) for the number of missed targets. The number of missed targets significantly increased during the second half of testing (P = 0.009). A one-way ANOVA on the first phase of testing revealed a main effect of treatment on missed targets (F(3, 21) = 7.577, P = 0.001). Scopolamine significantly increased missed targets during the first phase of testing (P = 0.002), and 0.3 mg/kg PQCA significantly reduced that effect (P = 0.009). In addition, 1.0 mg/kg PQCA showed a trend of attenuation (P = 0.074). A one-way ANOVA on the second phase of testing revealed a main effect of treatment on missed targets (F(3, 21) = 19.866, P < 0.001). Scopolamine significantly increased missed targets during the first phase of testing (P < 0.001), and the high dose of PQCA significantly reduced the scopolamine effect (P = 0.05).
Effect of PQCA on the scopolamine-impaired CPT in rhesus monkeys
Data are mean ± S.E.M, n = 8.

PQCA improves accuracy on the scopolamine impaired continuous performance task in rhesus monkeys. Scopolamine significantly impaired the hit rate, and PQCA (0.3–1.0 mg/kg) attenuated this impairment. *P < 0.05 and ***P < 0.001 compared with scopolamine treatment (n = 8).
In the color-discrimination control task, total accuracy (F(3, 18) = 0.680, P = 0.576) was not affected by scopolamine or PQCA; however, the two-way repeated measures ANOVA for omissions was significant for treatment (F(3, 18) = 5.233, P = 0.009); scopolamine alone (P = 0.038), scopolamine + 0.3 (P = 0.047), and scopolamine + 1.0 mg/kg PQCA (P = 0.023) produced more omissions than vehicle. Neither distractor type (F(1, 6) = 0.051, P = 0.829) nor the distractor type x treatment interaction (F(3, 18) = 0.205, P = 0.892) significantly affected omissions. Furthermore, a two-way repeated-measures ANOVA for accuracy revealed a main effect of distractor type (F(1, 6) = 19.364, P = 0.005), with difficult distractors producing more errors, but no main effect of treatment (F(3, 18) = 0.302, P = 0.823) or treatment x distractor-type interaction (F(3, 18) = 1.633, P = 0.217).
Discussion
The present studies demonstrate the procognitive effects of donepezil and the novel M1 PAM PCQA in two different, highly translatable cognition assays in rhesus monkeys. In addition, the combination of subeffective doses of PQCA and donepezil showed additivity/synergy in attenuating the scopolamine deficit in PAL. To our knowledge, this is the first report of an improvement in scopolamine-impaired NHP PAL and CPT by either an AChEI or an M1 PAM. These results further support the therapeutic potential of highly selective M1 PAMs for AD and other diseases characterized by cognitive dysfunction.
Paired-Associates Learning.
Deficits in PAL, a test of visuospatial memory dependent on the hippocampus, are among the earliest impairments observed in AD (Fowler et al., 2002; O’Connell et al., 2004; de Jager and Budge, 2005; Egerhazi et al., 2007; Junkkila et al., 2012). In healthy subjects, pharmacologic impairments and improvements in cholinergic function can produce deficits and rescue PAL performance, respectively (Harel et al., 2013).
In the present set of studies, scopolamine treatment significantly and reliably impaired PAL performance across all trial types. These results are consistent with Taffe et al. (2002), who showed an impairment of all trial types at the highest scopolamine doses tested. Although the doses of scopolamine tested here were individually titrated to produce a reliable impairment, it is worth noting that the mean dose tested (∼17 µg/kg) is the same as the high dose used by Taffe et al. (2002). Donepezil treatment significantly improved PAL performance for the moderately difficult 3-3 trials but not for the easiest (2-2) or the most difficult (4-4) trials. This pattern is similar to the effects of donepezil seen in the NHP delayed match to sample task (Buccafusco et al., 2008) in which donepezil only improved scopolamine deficits at the small to moderate delays but not at the longer, most demanding delays. We believe that the lack of significant effect on the demanding (4-4) trials might simply be due to the smaller dynamic range between the scopolamine- and vehicle-treated animals for those trials. Indeed, both donepezil and donepezil + PQCA trended toward significantly reversing the effects in the more demanding trials. Of course, the lack of donepezil’s effect on easy and difficult trial types might also be due to subject variability or a small sample size. Importantly, scopolamine-impaired PAL performance in humans is also attenuated by acute donepezil treatment, showing that the effects in NHPs appear to translate well to humans (Harel et al., 2013).
PQCA improved PAL performance for both easy and moderate trial types. It is unclear whether the difference in effect compared with donepezil on easy trials is due to a true difference in the mechanism of action or to between-study variability in the scopolamine deficit. Regardless, it is clear that PCQA produces procognitive effects at least equal in magnitude to donepezil in this task. The effective doses here are consistent with our group’s previous findings in the object-retrieval detour (ORD) tasks and self-ordered spatial search (Uslaner et al., 2013). These data further suggest that compounds which selectively increase M1 activity will improve cognitive function.
The combination of subeffective doses of PQCA and donepezil enhanced PAL performance. Interestingly, these effects were achieved with the same doses, which given in combination, improved performance in ORD (Vardigan et al., 2014). Considering the lack of side effects when combining low doses of PQCA and donepezil (Vardigan et al., 2014), a combination therapy may allow for improvement in cognition and a reduction in gastrointestinal side effects compared with donepezil alone.
Continuous-Performance Tasks.
Attentional impairments arise shortly after the memory deficits in AD (Parasuraman and Haxby, 1993; Parasuraman and Martin, 1994; Parasuraman et al., 1995; Perry and Hodges, 1999; Rizzo et al., 2000; Berardi et al., 2005; Bohnen et al., 2005). Similar to memory, activation of the cholinergic system improves attention in humans (Furey et al., 2006, 2008; Ricciardi et al., 2013) and NHPs (Callahan, 1999; O’Neill et al., 1999, 2003; Schwarz et al., 1999; Terry et al., 2002; Katner et al., 2004), whereas cholinergic blockade produces attentional impairments in both species (Callahan et al., 1993; Callahan, 1999; Taffe et al., 1999; Thienel et al., 2009a,b).
In this novel version of CPT, scopolamine treatment significantly and reliably impaired accuracy to the target stimulus while maintaining baseline false alarm rates for both easy and difficult distractors. Scopolamine treatment also increased reaction time in both studies, which theoretically could result from a reduction in attention or a decrease in motivation/motor function. The stability of false alarm rates during scopolamine treatment, however, suggests a sparing of motivation/motor function. False alarms to the difficult distractor did increase with scopolamine treatment, although this effect did not reach statistical significance. This increase in false alarms may signify an increase in impulsivity (Zeamer et al., 2011). Finally, scopolamine significantly impacted the number of misses during both the early and late phases of testing, findings that are consistent with the scopolamine-impaired mouse model of CPT described by Young et al. (2013). Importantly, in the current study, the magnitude of this effect increased in the late phase. This late stage increase in misses suggests a greater impact of scopolamine on sustained attention, which has been shown to be impaired in AD patients (Sahakian et al., 1989; Mendez et al., 1997; Levin et al., 1998; White and Levin, 1999).
Donepezil treatment attenuated the scopolamine impairments in accuracy, reaction time, and misses over time. Similarly, PQCA treatment attenuated the scopolamine impairments in accuracy and misses over time but not reaction time. The reason for the lack of effect of PQCA on reaction time compared with donepezil is unclear. Some evidence suggests that reaction time is influenced more by nicotinic receptors than by muscarinic receptors (Mirza and Stolerman, 2000). Thus, although donepezil’s widespread effect on muscarinic and nicotinic receptors might be responsible for many of its adverse effects, donepezil’s activation of nicotinic receptors might be beneficial for reaction time improvements. Additional research is needed to test this hypothesis.
It is important to acknowledge that the scopolamine model is limited and is not meant to model the neuronal loss, plaques, and tangles observed in AD. Furthermore, the behavioral disturbances of AD do not manifest with this model. We do believe, however, that the scopolamine model is useful for preclinical translation for a number of reasons. First, many of the symptoms observed in AD are thought to be due to a loss of cholinergic function, which is meant to be modeled by the muscarinic receptor antagonist scopolamine. Indeed, many of the same cognitive domains affected by scopolamine are impacted in AD (Ebert and Kirch, 1998), and donepezil, the standard of care for AD, produces its effects by increasing cholinergic neurotransmission. Second, AD patients perform an average of two to four standard deviations below normal age-matched controls (Pillon et al., 1991). In both the tasks described herein, the scopolamine deficit represents a two to three standard deviation reduction from baseline performance, which suggests a similar level of impairment to AD patients. Third, donepezil is able to attenuate a scopolamine deficit in healthy volunteers in the PAL task, and AD patients show a modest but significant improvement with donepezil treatment. Thus, there is some pharmacologic validation of the scopolamine model in humans, and the present findings demonstrate that similar responses are seen preclinically.
Here we have demonstrated the ability of the highly specific M1 PAM, PQCA to attenuate a scopolamine deficit in two very different and highly translatable NHP cognition assays. The expression and magnitude of PQCA’s effect were similar to the clinical standard, donepezil, for both visuospatial memory (PAL) and attention (CPT), and the effects of donepezil observed here are similar to those observed in the human scopolamine model. Thus, the use of NHP models like these should improve the probability of success in the clinic by accurately predicting which cognitive tests and domains will be impacted by novel medicines, such as M1 PAMs.
Acknowledgments
The authors thank the Merck Chemistry group for preparing the compounds for these studies, the Merck SALAR group for dosing assistance and exceptional animal care, Spencer Tye for helpful discussions regarding assay design, and Joshua Vardigan for comments on this manuscript.
Authorship Contributions
Participated in research design: Lange, Cannon, Drott, Uslaner.
Conducted experiments: Lange, Cannon, Drott.
Contributed new reagents or analytic tools: Kuduk.
Performed data analysis: Lange, Cannon, Drott.
Wrote or contributed to the writing of the manuscript: Lange, Cannon, Drott, Uslaner.
Footnotes
- Received June 11, 2015.
- Accepted October 6, 2015.
Abbreviations
- AchEI
- acetylcholinesterase inhibitors
- AD
- Alzheimer disease
- ANOVA
- analysis of variance
- CD
- color discrimination
- CPT
- continuous-performance task
- NHP
- nonhuman primate
- ORD
- object-retrieval detour
- PAL
- paired-associates learning
- PAM
- positive allosteric modulator
- PQCA
- (1-((4-cyano-4-(pyridine-2-yl)piperidin-1-yl)methyl-4-oxo-4 H-quinolizine-3-carboxylic acid)
- Copyright © 2015 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}