


Abstract
Heart failure–mediated skeletal myopathy, which is characterized by muscle atrophy and muscle metabolism dysfunction, often manifests as dyspnea and limb muscle fatigue. We have previously demonstrated that increasing Ca2+ sensitivity of the sarcomere by a small-molecule fast skeletal troponin activator improves skeletal muscle force and exercise performance in healthy rats and models of neuromuscular disease. The objective of this study was to investigate the effect of a novel fast skeletal troponin activator, CK-2127107 (2-aminoalkyl-5-N-heteroarylpyrimidine), on skeletal muscle function and exercise performance in rats exhibiting heart failure–mediated skeletal myopathy. Rats underwent a left anterior descending coronary artery ligation, resulting in myocardial infarction and a progressive decline in cardiac function [left anterior descending coronary artery heart failure (LAD-HF)]. Compared with sham-operated control rats, LAD-HF rat hindlimb and diaphragm muscles exhibited significant muscle atrophy. Fatigability was increased during repeated in situ isokinetic plantar flexor muscle contractions. CK-2127107 produced a leftward shift in the force-Ca2+ relationship of skinned, single diaphragm, and extensor digitorum longus fibers. Exercise performance, which was assessed by rotarod running, was lower in vehicle-treated LAD-HF rats than in sham controls (116 ± 22 versus 193 ± 31 seconds, respectively; mean ± S.E.M.; P = 0.04). In the LAD-HF rats, a single oral dose of CK-2127107 (10 mg/kg p.o.) increased running time compared with vehicle treatment (283 ± 47 versus 116 ± 22 seconds; P = 0.0004). In summary, CK-2127107 substantially increases exercise performance in this heart failure model, suggesting that modulation of skeletal muscle function by a fast skeletal troponin activator may be a useful therapeutic in heart failure–associated exercise intolerance.
Introduction
Despite advances in device and pharmacological therapy, heart failure (HF) continues to be a leading cause of morbidity and mortality. Although cardiac dysfunction is the central phenotypic abnormality, the compensatory hemodynamic, autonomic, and neurohormonal responses to this dysfunction ultimately lead to a multisystem syndrome that contributes to overall morbidity (Piepoli et al., 2010a,b). Skeletal muscle is among the organ systems affected by HF, as changes in muscle mass, myocyte calcium handling, capillary density, muscle fiber type, mitochondrial structure, and oxidative capacity have been observed in patients and animal models of HF (Drexler et al., 1992; Perreault et al., 1993; Schieffer et al., 1995; Anker et al., 1997; Delp et al., 1997; Duscha et al., 1999; Ward et al., 2003). These alterations contribute to decreased muscle strength and endurance (Pina et al., 2003; Toth et al., 2010) and, collectively, this skeletal myopathy is believed to be a substantial contributor to exercise intolerance and dyspnea, resulting in disability and diminished quality of life in patients with HF (Pina et al., 2003).
The sarcomere is the basic contractile unit of skeletal muscle. Within the sarcomere, the calcium ion (Ca2+) is a key regulatory signaling molecule, as the binding of Ca2+ to troponin C in the troponin complex permits tropomyosin movement, consequent actin-myosin interaction, and muscle contraction (Brooks, 2004). The magnitude of Ca2+ release increases proportionally with the level of neuromuscular transmission and, in turn, at submaximal levels of neuromuscular activation, the force of contraction increases proportionally with calcium release (Chin, 2010).
Modulation of the sarcomere is an emerging therapeutic target of interest to treat heart and skeletal muscle–related diseases (Ochala, 2010; Malik et al., 2011; Kho et al., 2012a). We have demonstrated that the small molecule tirasemtiv (formerly CK-2017357) specifically binds to fast skeletal troponin and slows the off rate of Ca2+ from troponin, thus sensitizing the complex to Ca2+ ions (Russell et al., 2012). This mechanism of action leads to a significantly higher force output at submaximal levels of motor nerve stimulation in animals and humans (Russell et al., 2012; Hansen et al., 2014). Tirasemtiv and its structural analogs have demonstrated efficacy in improving muscle strength in preclinical disease models of neuromuscular origin, including a mouse model of nemaline myopathy (Lee et al., 2013; Ottenheijm et al., 2013), rat model of myasthenia gravis (Russell et al., 2012), and mouse model of amytrophic lateral sclerosis (Hwee et al., 2014). Furthermore, in clinical trials, tirasemtiv improved respiratory function and other measures of skeletal muscle strength and endurance in patients with amytrophic lateral sclerosis (Shefner et al., 2012), myasthenia gravis, and exercise-limiting calf claudication (Bauer et al., 2014).
Increasing sarcomeric calcium sensitivity by a fast skeletal troponin activator might mitigate the skeletal muscle dysfunction observed in HF. In this study, we examined the effect of CK-2127107, a novel fast skeletal troponin activator that is structurally distinct from tirasemtiv in rats with induced systolic heart failure that exhibited characteristics of skeletal myopathy and substantially reduced exercise performance.
Materials and Methods
Small Molecule CK-2127107.
CK-2127107 (2-aminoalkyl-5-N-heteroarylpyrimidine) (Yang et al., 2011), was identified and optimized as a small-molecule fast skeletal troponin activator candidate. The CK-2127107 used in these experiments was synthesized at Cytokinetics (South San Francisco, CA).
Myofibril Isolation and Protein Purification.
Slow skeletal, fast skeletal, and cardiac myofibrils were prepared from slow bovine masseter, fast rabbit psoas, and bovine heart muscle, respectively, as previously described (Solaro et al., 1971; Young and Davey, 1981; Herrmann et al., 1993). Skeletal isoforms of myosin, actin, troponin, and tropomyosin were purified from rabbit psoas muscle, and cardiac isoforms were purified from the bovine heart, as previously described (Margossian and Lowey, 1982; Pardee and Spudich, 1982; Potter, 1982; Smillie, 1982). Troponin-tropomyosin–regulated actin myosin was reconstituted according to published methods (Ebashi et al., 1967; Spudich and Watt, 1971).
ATPase Assays.
Myosin ATPase activity was measured as previously described (De La Cruz and Ostap, 2009; Russell et al., 2012). Fast and slow skeletal myofibril steady-state activity was measured in a buffer containing 12 mmol/l 1,4-piperazinediethanesulfonic acid (Pipes), 2 mmol/l MgCl2, 60 mmol/l KCl, and 1 mmol/l dithiothreitol at a pH of 6.8. Activity was measured at 25°C in a pyruvate kinase and lactate dehydrogenase–coupled enzyme system. This assay regenerates myosin-produced ADP into ATP by oxidizing NADH, producing an absorbance change at 340 nm. A SpectraMax plate reader (Molecular Devices, Sunnyvale, CA) monitored the change in absorbance as a function of time in the presence of varying concentrations of CK-2127107.
Measurement of Calcium Release from Troponin.
Calcium release from the troponin complex was measured using an adapted protocol from Rosenfeld and Taylor (1985). Quin-purified rabbit skeletal troponin was exchanged into 1× PM12 buffer (12 mmol/l K-Pipes, 2 mmol/l MgCl2, pH 6.8) by passage over a P-6DG column (Bio-Rad, Hercules, CA) that was pre-equilibrated in that buffer. Troponin concentrations were determined by UV absorbance (278 nm) in 6 mol/l guanidine-HCl using an extinction coefficient of 0.45 for a 1 mg/ml solution. Quin-2 (Invitrogen, Carlsbad, CA) was dissolved in dimethylformamide and diluted in 1× PM12 just prior to use. Transient fluorescence measurements were made using an SF-61DX stopped-flow fluorimeter (Hi-Tech Scientific, Bradford-on-Avon, UK), with excitation provided by a monochromator (337 nm, 10-mm slit width) and emission measured through a glass filter (495-nm long pass). Fluorescence intensities were translated into calcium concentrations using calcium standard curves prepared in the presence of equivalent concentrations of dimethyl sulfoxide (DMSO) and CK-2127107, as were used in the transient reactions. Released calcium concentrations were expressed as the final concentration after mixing.
Isothermal Titration Calorimetry.
Isothermal titration calorimetry (ITC) was performed in a Micro-Cal VP-ITC microcalorimeter (GE Healthcare, Inc., Piscataway, NJ) at 25°C. Titrations were conducted by filling the sample chamber with 25 μmol/l rabbit fast skeletal troponin complex in 12 mmol/l Pipes, 100 mmol/l KCl, 250 μmol/l CaCl2, 3% DMSO, and 5 mmol/l β-mercaptoethanol. The ligand solution that was injected into the sample chamber contained 300 μmol/l CK-2127107 in a buffer consisting of 12 mmol/l Pipes, 100 mmol/l KCl, 250 μmol/l CaCl2, 3% DMSO, and 5 mmol/l β-mercaptoethanol. Ligand injections were made every 300 seconds at a volume of 10 µl. To correct for the heats of dilutions of CK-2127107, the stable heat signal from the injections near the end of the experiment was subtracted from the remaining values. All data collection and analysis was conducted using the modified Origin software (OriginLab, Northampton, MA) included with the instrument using a single binding site model. Troponin concentrations were determined by UV absorbance (278 nm) in 6 mol/l guanidine-HCl using an extinction coefficient of 0.45 for a 1-mg/ml solution.
Sham and Heart Failure Rats.
Rats used in this study were maintained in accordance with the Guide for the Care and Use of Laboratory Animals of the Institute (Seventh Edition, National Research Council) and under the supervision of the Cytokinetics Institutional Animal Care and Use Committee. Ligation of the left anterior descending coronary artery was performed in female Sprague-Dawley rats at Charles River Laboratories (Raleigh, NC). Sham surgeries were also performed with the same surgical preparation, except that ligation with the suture material was not performed. Sham (n = 25) and left anterior descending coronary artery (LAD) heart failure (LAD-HF) (n = 21) rats were shipped to the investigators’ animal facility. After 3 days of acclimation, cardiac function was assessed by echocardiography in all rats. Cardiac function was measured longitudinally at 2, 4, 7, and 10 weeks after arrival.
Histology.
Sham and LAD-HF hearts were excised, weighed, fixed in 10% formalin for 24 hours, and embedded in paraffin. Heart sections were stained with Masson’s Trichome for visualization of collagen deposition. The diaphragm, soleus, and extensor digitorum longus (EDL) were excised, cleaned, pinned to a corkboard, and frozen in melting isopentane. Serial frozen cross sections were cut at 10 µm and stained for myosin ATPase after preincubation at a pH of 4.35. Digital images were obtained under 200× total magnification (Olympus BX41; Olympus America, Melville, NY) and analyzed by Axiovision software (Zeiss, Jena, Germany). Stained fibers were classified as type I, IIa, or II b/x and measured for the individual myofiber cross-sectional area (µm2).
Western Blot Analysis.
Soleus muscle was homogenized in a cell lysis buffer (Life Technologies, Grand Island, NY). Fifty micrograms of protein was prepared in a Laemmli sample loading buffer, separated by SDS-PAGE, and transferred to a nitrocellulose membrane. The membranes were incubated overnight at 4°C in a blocking buffer with sarcoendoplasmic reticulum Ca2+ ATPase 2a (SERCA2a; 1:1000; Thermo Scientific, Waltham, MA) and glyceraldehyde 3-phosphate dehydrogenase (1:12,500; Cell Signaling Technology, Danvers, MA). Following three rinses in 1× Tris-buffered saline/Tween 20, membranes were incubated with corresponding fluorescent secondary antibodies (1:10,000; LI-COR Biosciences, Lincoln, NE). After three rinses, membranes were visualized with a LI-COR infrared imaging system. Image quantification was performed using Odyssey 2.1 software (LI-COR).
Skinned Muscle Fibers Force–pCa Relationship.
Muscle tissue for in vitro skinned fiber studies was prepared using an adapted protocol based on Lynch and Faulkner (1998). Briefly, rat diaphragm, soleus, and EDL muscle from sham and LAD-HF rats were rapidly dissected, rinsed in physiologic saline, and then incubated in a skinning solution (125 mmol/l K-propionate, 20 mmol/l imidazole, 5 mmol/l EGTA, 2 mmol/l MgCl2, 2 mmol/l ATP, pH 7.0) supplemented with 0.5% Triton X-100 (Sigma-Aldrich, St. Louis, MO) for 30 minutes at 4°C. The buffer was then changed to a storage solution (125 mmol/l K-propionate, 20 mmol/l imidazole, 5 mmol/l EGTA, 2 mmol/l MgCl2, 2 mmol/l ATP, 50% glycerol, pH 7.0) and stored at −20°C for later use.
For skinned fiber analysis, single muscle fibers were dissected from larger segments of tissue in a rigor buffer at 4°C (20 μmol/l 4-morpholinepropanesulfonic acid, 5 μmol/l MgCl2, 120 μmol/l potassium acetate, 1 μmol/l EGTA, pH 7.0). The fibers were then suspended between a 400A force transducer (Aurora Scientific, Aurora, ON, Canada) and a fixed post and secured with 2–4 μl of a 5% solution of methylcellulose in acetone. Fibers were then incubated at 10°C in a relaxing buffer (20 μmol/l 4-morpholinepropanesulfonic acid, 5.5 μmol/l MgCl2, 132 μmol/l potassium acetate, 4.4 μmol/l ATP, 22 μmol/l creatine phosphate, 1 mg/ml creatine kinase, 1 mM dithiothreitol, 44 ppm antifoam 204, pH 7.0) and the baseline tension was adjusted. Tension was generated in each fiber by changing the fiber buffer over to a relax buffer supplemented with 1 mmol/l EGTA and a 15 mmol/l solution of calcium chloride and calculated using a web resource (http://www.stanford.edu/~cpatton/webmaxc/webmaxcS.htm). Relax buffer pCA concentrations ranged from 8 (low Ca2+ activation and fiber tension) to 4 (saturated Ca2+ activation and maximum tension). CK-2127107 was added to these buffers from a DMSO solution (final DMSO concentration = 1%).
In Situ Muscle Contractile Characteristics.
Isometric and isokinetic hindlimb force was measured in sham and LAD-HF rats in the presence and absence of CK-2127107 (10 mg/kg i.v. in 50% polyethylene glycol:40% Cavitron [Ligand Pharmaceuticals Inc., La Jolla, CA]:10% dimethylacetamide). Rats were placed under anesthesia with isoflurane (1–5%). One incision was made on the midthigh region of the right leg to expose the sciatic nerve. To prevent cocontraction of the ankle dorsiflexors, an additional incision was made lateral to the patella to isolate and sever the peroneal nerve. Rats were then placed on a temperature-maintained in situ muscle analysis rig (Model 806C; Aurora Scientific). The knee was immobilized in a clamp between two sharpened screws, and the foot was taped to a footplate attached to a force transducer (Aurora Scientific). Stainless steel needle electrodes (0.10 mm) were hooked around the exposed sciatic nerve. Sham and LAD-HF rats were treated with vehicle or CK-2127107 (10 mg/kg i.v.). Muscle contractile properties were assessed by applying an electrical current (under supramaximal voltage conditions) to the nerve and recording the force generated by the muscle via a servomotor. An isometric force–frequency relationship (10–150 Hz, 1-millisecond pulse width, 350-millisecond train duration) was assessed with the ankle joint at 90° of flexion. An isokinetic force-velocity relationship in response to a 30-Hz stimulation was assessed over a range of 0–20.1 radians/s. The fatigue properties of the ankle plantar flexor muscles were assessed by repeated isokinetic contractions (30-Hz stimulation, 3.1 rad/s, once per second) over a 10-minute period.
Diaphragm Force–Frequency Relationship.
Diaphragm contractile force was measured by electrical field stimulation in an organ bath system (Radnoti, Monrovia, CA). The diaphragm and last floating rib from sham and LAD-HF rats were excised, rinsed in physiologic saline, and placed in a temperature controlled water-jacketed chamber (26–27°C) containing Krebs-Henseleit buffer (118 mmol/l NaCl, 10 mmol/l glucose, 4.6 mmol/l KCl, 1.2 mmol/l KH2PO4, 1.2 mmol/l MgSO4·7H2O, 24.8 mmol/l NaHCO3, 2.5 mmol/l CaCl2, 50 mg/l tubocurarine, 50 U/l insulin, pH 7.4) that was continuously aerated with 95% O2/5% CO2. After 10 minutes of equilibration, vertical strips spanning the floating rib to the central tendon were cut from diaphragms. Braided silk sutures were tied at the central tendon and floating rib and attached to a force transducer between two platinum electrodes. Diaphragm strips were set to a length that produced maximum twitch tension (Lo). The force-frequency profile of the muscle was obtained by stimulating the muscle at frequencies between 10 and 150 Hz (Grass Stimulator [Grass Technologies, Warwick, RI], 800-millisecond train duration, 0.6-millisecond pulse width). CK-2127107 was suspended in DMSO and directly added into the bath.
Rotarod Exercise Assay.
Exercise performance was assessed in sham and LAD-HF rats utilizing a rotarod protocol developed at Cytokinetics. At least 3 days after echocardiography, rotarod performance was assessed at weeks 4, 7, and 10 to observe the decline of exercise performance. The rotarod assay consisted of a training day that was immediately followed by an assessment day. The training day consisted of two sessions: a constant speed of 10 rpm for 5 minutes in the morning and an accelerating speed of 14–16 rpm over 5 minutes in the afternoon. On the assessment day, rats were run again at an increasing speed from 14 to 16 rpm over 5 minutes. Rats were then immediately run at a constantly accelerating rate from 12 to 25 rpm over the course of 10 minutes. The time to fall was recorded, with the test terminated at 600 seconds. Once exercise-intolerant LAD-HF rats with a fractional shortening of less than 25% were identified, sham and LAD-HF rats were advanced to assessment of rotarod running under therapeutic intervention.
A blinded cross-over study design was employed to investigate the effect of CK-2127107 (10 mg/kg p.o.) and its vehicle [19.3% polyethylene glycol:80% (15%) Captisol, pH 3, 0.2% Tween 80, 0.5% hydroxypropyl methylcellulose] dosed 30 minutes prior to evaluating rotarod performance. At each assessment time point, all rats were dosed with either CK-2127107 or vehicle on the first day, and on the next day, each rat received the converse treatment. Each day consisted of a 5-minute primer session, whereby rats were run at an increasing speed from 14 to 16 rpm. Rats were then run at a constantly accelerating rate from 12 to 25 rpm over the course of 10 minutes. Time to fall was recorded, with the test being terminated at 600 seconds. Only LAD-HF rats that ran less than 300 seconds on their vehicle day at week 10 and thus exhibited signs of exercise intolerance were included in the study.
Statistical Analyses.
Statistical analyses between mean averages employing one-way analysis of variance and unpaired t tests were performed as appropriate using GraphPad Prism software version 6.03 (La Jolla, CA). Repeated measures analysis of covariance (ANCOVA) was performed with SAS software version 9.2 (Cary, NC). Unless noted otherwise, results are expressed as mean ± S.E.M. Significance was set at P values < 0.05.
Results
CK-2127107 Selectively Binds to and Sensitizes the Fast Skeletal Troponin Complex to Calcium.
CK-2127107 (Yang et al., 2011) was identified and optimized as a small-molecule fast skeletal troponin activator candidate utilizing high-throughput screens of type II, fast skeletal myofibrils. Using purified cardiac, slow skeletal, and fast skeletal myofibrils, the specificity, potency, and mechanism of action of CK-2127107 was characterized through a series of biochemical assays.
Calcium activates the myosin ATPase activity of skinned (detergent permeabilized) myofibrils. The midpoint of the ATPase-calcium curve relationship is defined as its half maximal activation (pCa50) and is a measure of the sensitivity of the sarcomere to calcium. The addition of either 2.5, 10, or 40 μmol/l CK-2127107 to fast skeletal rabbit psoas myofibrils resulted in a concentration-dependent leftward shift of the myosin ATPase-calcium relationship, with the ATPase activity pCa50 shifting from a control value of 5.67 to 5.99 (2.5 µmol/l), 6.23 (10 μmol/l), and 6.55 (40 μmol/l) (Fig. 1A). The muscle-type selectivity of CK-2127107 was examined using skinned myofibrils prepared from different muscles, i.e., rabbit psoas (fast skeletal), bovine masseter (slow skeletal), and bovine heart (cardiac), as previously described (Russell et al., 2012). CK-2127107 selectively activated fast skeletal myofibrils (EC50 = 3.4 μmol/l; maximal activation = 3.6-fold). CK-2127107 had no effect on slow skeletal or cardiac myofibrils, demonstrating its selectivity for fast skeletal muscle (Fig. 1B).

Characterization of the fast skeletal troponin activator CK-2127107. (A) The ATPase activity of skinned rabbit fast skeletal myofibrils was measured using a pyruvate kinase/lactate dehydrogenase–coupled assay. In the presence of CK-2127107, the control pCa50 of 5.67 (DMSO; circles) shifts to 5.99 (2.5 μmol/l; squares), 6.23 (10 μmol/l; triangles), and 6.55 (40 μmol/l; inverted triangles). Plotted are the means ± S.E.M. for two experiments. (B) Concentration-response ATPase activity analysis was performed at a tissue-specific pCa25, with fast skeletal (rabbit), slow skeletal (bovine), and cardiac (bovine) detergent-skinned myofibrils (n = 4 per group). Data were fitted using a four-parameter concentration-response equation, resulting in a calculated fast skeletal maximal fold activation of 3.6 and an EC50 of 3.4 μmol/l. Slow skeletal and cardiac myofibril activation was insufficient to produce reliable EC50 estimates. (C) Activation of the thin filament (as indicated by an increase in cardiac myosin ATPase observed at a fixed calcium concentration) is found in the presence of mixed thin filaments containing the fast skeletal isoform of troponin but not the cardiac isoform of troponin. The tropomyosin isoform did not substantially affect the myosin ATPase rate, indicating that the fast skeletal troponin complex is required for compound activity. (D) CK-2127107 (green line) slows release of Ca2+ from skeletal troponin. Calcium release was monitored using the fluorescence enhancement of Quin-2 upon binding to calcium.
Having established the specificity of CK-2127107 for fast skeletal myofibrils, we leveraged that property to identify the specific target of CK-2127107 in the fast skeletal sarcomere. Individual components of the sarcomere, including the thin filament proteins actin, troponin, tropomyosin, and the thick filament protein myosin can be purified from different sources (fast, slow, or cardiac) and reconstituted in different combinations to generate similar ATPase activity and calcium dependence as intact skinned myofibrils. Using cardiac myosin as a probe for thin filament activation, CK-2127107 increased its cardiac myosin ATPase activity in only those heterologous reconstituted versions of the thin filament that contained fast skeletal troponin, demonstrating that CK-2127107 interacts specifically with the fast skeletal troponin complex (Fig. 1C).
The observed selectivity and activity in heterologous reconstituted assays suggested that CK-2127107 interacts directly with fast skeletal troponin. This interaction was verified using ITC to directly characterize its binding interaction. Titration of CK-2127107 into a solution of purified rabbit fast skeletal troponin complex (TnT/TnI/TnC) resulted in an exothermic reaction that fit a single site binding model (Supplemental Fig. 1). Pooled data from several experiments indicated that CK-2127107 bounded to fast skeletal troponin with high affinity (Kd = 3.0 ± 0.06 µmol/l; n = 5), and with a stoichiometry consistent with a single binding site per troponin complex (n = 1.24 ± 0.016). One potential mechanism for sensitizing the sarcomere to calcium is through stabilization of the calcium-troponin complex. Stabilization slows the rate of calcium dissociation from the troponin complex, thus prolonging the time troponin spends in the active (open) conformation. The rate of calcium dissociation from troponin was monitored by rapidly mixing the calcium-saturated, fast skeletal troponin complex with Quin-2, a fluorescent calcium chelator whose fluorescence intensity increases when it binds calcium. When CK-2127107 was tested in this assay, the calcium release rate slowed (from 24.1 to 9.8 s−1), consistent with a stabilization of the Ca2+-bound, “active” conformation of troponin (Fig. 1D). Thus, CK-2127107 increases calcium sensitivity, and by slowing the calcium release rate from troponin, exerts a leftward shift on the force-calcium relationship of muscle.
Skeletal Muscle Limb Atrophy Occurs in Rats with Systolic Dysfunction Following Myocardial Infarction.
As previously reported (Pfeifer et al., 2001; Koh et al., 2003), skeletal muscle atrophy and exercise intolerance occurs in the rat LAD-HF model produced by ligation of LAD. Ligation of the LAD to produce myocardial infarction was performed on 8-week-old female Sprague-Dawley rats, and cardiac function was serially assessed by echocardiography every 2–3 weeks following surgery. Compared with rats undergoing a sham surgery, significant decreases in fractional shortening (49.6 ± 2.2% versus 28.3 ± 3.0%; P < 0.0001) were observed in LAD-HF rats 2 weeks after ligation, and cardiac function continued to gradually decline during the next 8 weeks (Fig. 2A). Sham and LAD-HF hearts were collected postmortem for mass and histologic assessment. Heart mass, normalized to body mass, was significantly higher in LAD-HF rats than in the sham controls (4.3 ± 0.1 versus 3.5 ± 0.01 mg heart/g body mass; P < 0.0001; Fig. 2B), which is consistent with the pathologic cardiac hypertrophy that typically occurs in myocardial infarction–induced heart failure (Rubin et al., 1983; Purdham et al., 2008). Masson’s trichrome stains of heart sections from sham controls and LAD-HF rats revealed extensive collagen disposition distal to the ligation site in the left ventricle of LAD-HF rats (Supplemental Fig. 2).

Coronary LAD artery ligation produces cardiac dysfunction and skeletal muscle atrophy. (A) Echocardiography revealed significant decreases in left ventricular fractional shortening over time in LAD-HF rats (n = 13–21 group; ****P < 0.0001 versus sham). (B) Heart mass (normalized to body mass) is significantly higher in LAD-HF rats, suggestive of pathologic hypertrophy (21–25 rats/group; ****P < 0.0001 versus sham). (C) LAD-HF plantarflexor muscles (soleus and plantaris) had a significantly smaller myofiber area and exhibited a leftward shift in fiber size distribution (n = 5–7 per group; *P < 0.05).
We evaluated the skeletal muscle mass and myofiber size characteristics of sham and LAD-HF rats. The hindlimb muscle mass, normalized to body mass, was not significantly different between sham and LAD-HF groups (soleus: 0.49 ± 0.01 versus 0.48 ± 0.01 mg muscle/g body mass; plantaris: 1.2 ± 0.04 versus 1.1 ± 0.04 mg muscle/g body mass). By mass, there was a trend for smaller muscles in LAD-HF rats compared with sham rats (sham versus LAD-HF soleus: 160 ± 4 versus 148 ± 4 mg; P = 0.059; plantaris: 372 ± 11 versus 347 ± 8 mg; P = 0.09). We also evaluated the myofiber cross-sectional area in histologic sections from hindlimb EDL, soleus, and plantaris muscles. The soleus and plantaris myofiber area was significantly smaller in LAD-HF rats than in sham control rats (sham versus LAD-HF soleus: 3039 ± 122.9 µm2 versus 2338 ± 245 µm2; plantaris: 3025 ± 230 versus 2349 ± 176 µm2; n = 5–8 per group; P < 0.05; Fig. 2C), demonstrating that the condition of heart failure led to changes in skeletal muscle myofiber morphology.
Soleus Muscle Sarcoendoplasmic Reticulum Ca2+ ATPase Protein Is Reduced in LAD-HF Rats.
Soleus muscle SERCA is responsible for the reuptake of calcium ions from the cytoplasm into the sarcoplasmic reticulum. Previous studies have suggested that decreases in SERCA protein due to HF may contribute to calcium dysregulation and overall HF skeletal myopathy (Simonini et al., 1999; Bueno et al., 2010; Middlekauff et al., 2012). In this study, SERCA2a protein levels in LAD-HF soleus muscle was significantly lower than in sham soleus muscle (Supplemental Fig. 4).
CK-2127107 Increases Ca2+ Sensitivity in Fast Skeletal Muscle Fibers and Increases Muscle Force in Response to Submaximal Stimulation Frequencies in LAD-HF Rats.
Fast muscle fibers from EDL and slow muscle fibers from soleus muscle from sham and LAD-HF rats were harvested and chemically skinned to assess the single fiber force–pCa relationship. CK-2127107 (3 µmol/l) produced a significant leftward shift of the force-pCa relationship in both sham and LAD-HF EDL muscle fibers (pCa5 sham: DMSO versus CK-2127107 is 5.49 ± 0.06 versus 5.85 ± 0.05; LAD-HF: DMSO versus CK-2127107 is 5.5 ± 0.04 versus 5.86 ± 0.03; n = 7–9 per group; Fig. 3A), consistent with the calcium sensitizing effect of CK-2127107. As expected, given its selectivity for fast skeletal muscle troponin, CK-2127107 (10 µmol/l) did not have an effect on the force-pCa relationship of slow fibers from soleus muscle (Fig. 3A).

CK-2127107 increases Ca2+ sensitivity, submaximal force, and fatigue resistance in situ. (A) CK-2127107 (3 µmol/l) caused a leftward shift in the force-Ca2+ relationship in skinned sham and LAD-HF fast fiber EDL muscle. No changes were observed in slow fiber soleus muscle with CK-2127107 treatment. (B) LAD-HF plantarflexor muscles produced a significantly lower peak isometric force (*P < 0.05 versus sham). CK-2127107 significantly increased submaximal isometric tension up to 60-Hz stimulation in LAD-HF plantarflexor muscles (†P < 0.01 versus vehicle). (C) CK-2127107 significantly increased the force-isokinetic velocity relationship in both sham and LAD-HF plantarflexor muscles at 30-Hz stimulation (†P < 0.001 versus respective vehicle groups). (D) LAD-HF plantarflexor muscles produced significantly less isokinetic force during a fatigue-inducing repeated stimulations protocol (*P < 0.01 versus sham). (E) CK-2127107 significantly increased work in both sham and LAD-HF plantarflexor muscles in response to 300 repeated stimulations (bar graph: mean ± S.E.M.; *P < 0.05; **P < 0.01; ****P < 0.0001).
Significant decreases in isometric and isokinetic leg muscle force and increased muscle fatigability have been previously observed in heart failure patients (Clark et al., 1997; Harrington et al., 1997; Gielen et al., 2012). The in situ isometric and isokinetic force characteristics of the rat ankle plantarflexor muscles in sham and LAD-HF rats were measured following sciatic nerve stimulation. Isometric muscle force was significantly lower in LAD-HF rat plantarflexor muscles at stimulation frequencies from 60 Hz up to tetanus (maximum activation) at 150 Hz (Fig. 3B) Administration of CK-2127107 (10 mg/kg i.v.) significantly increased plantar flexor isometric force in response to subtetanic nerve frequency stimulations to the same extent in both sham and LAD-HF rats (Fig. 3B). Compared with vehicle treatment, CK-2127107 increased the isometric force to the greatest magnitude in response to the 30-Hz stimulation frequency. Thus, to provide the most sensitive assessment of CK-2127107, 30 Hz was selected for subsequent in situ muscle function evaluation. The isokinetic plantarflexor force output at 30-Hz stimulation was similar between vehicle-treated sham and LAD-HF rats. CK-2127107 significantly increased the isokinetic force to the same extent in both sham and LAD-HF rats (Fig. 3C). Thus, increasing sarcomere Ca2+ sensitivity by CK-2127107 treatment increases both the isometric and isokinetic force at an equal, submaximal, stimulation frequency in the presence or absence of heart failure.
As a measure of muscle fatigability in situ, the sham and LAD-HF ankle plantar flexor muscle groups were subjected to 300 repetitive isokinetic contractions (30-Hz stimulation, 3.1 rad/s) over a period of 10 minutes. Administration of CK2127107 (10 mg/kg i.v.) significantly increased initial force at 30 Hz in both LAD-HF and sham animals (Fig. 3D). After 300 contractions, LAD-HF muscle produced less total work than sham muscles (6.74 ± 0.25 versus 7.62 ± 0.24 J/g; P < 0.05; Fig. 3E), suggesting increased fatigability in LAD-HF skeletal muscles. Sham and LAD-HF rats treated with CK-2127107 (10 mg/kg i.v.) significantly increased total work and attenuated fatigue throughout the entire protocol (sham: 7.62 ± 0.24 versus 9.82 ± 0.53 J/g; P < 0.01; LAD-HF: 6.74 ± 0.25 versus 10.58 ± 0.49 J/g; P < 0.0001; Fig. 3, D and E).
CK-2127107 Significantly Increases Ca2+ Sensitivity and Submaximal Diaphragm Force.
Respiratory muscle weakness has been highlighted as a specific contributor to dyspnea and exercise intolerance in heart failure patients (Meyer et al., 2001). The diaphragm is a primary muscle involved in respiration, and dysfunction of this muscle is specifically a noted complication of heart failure (Meyer et al., 2001; van Hees et al., 2008). In this study, the LAD-HF diaphragm myofiber area was significantly smaller than that of sham diaphragms (1534 ± 114.7 versus 1141 ± 77.9 µm2; P < 0.05; Fig. 4A; Supplemental Fig. 3).
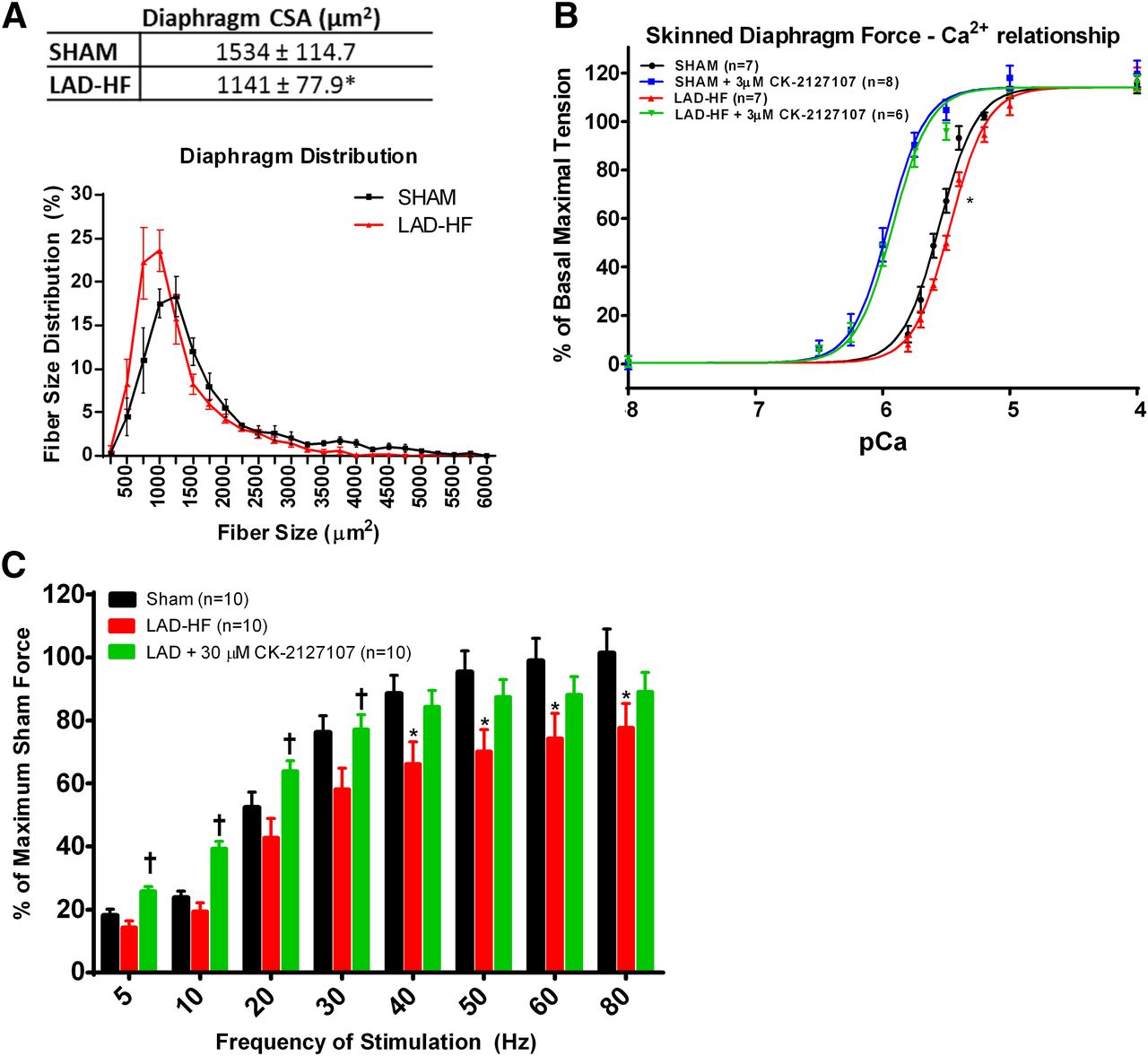
CK-2127107 increases Ca2+ sensitivity and submaximal force in LAD-HF diaphragms with characteristics of skeletal myopathy. (A) LAD-HF myofiber area was significantly smaller than sham myofibers. Fiber size distribution reveals a leftward shift in LAD-HF diaphragm fiber size (n = 6–7 per group; *P < 0.05). (B) Skinned LAD-HF diaphragm fibers exhibit reduced Ca2+ sensitivity compared with sham diaphragm fibers, with a significant rightward shift in the LAD-HF force-calcium relationship (*P < 0.05 based on pEC50 values), demonstrating reduced calcium sensitivity in LAD-HF fibers. CK-2127107 (3 µM) significantly increased the force-pCa relationship in both sham and LAD-HF diaphragm fibers. (C) LAD-HF diaphragm muscles demonstrate a lower isometric peak force (*P < 0.05 versus sham). CK-2127107 significantly increased force in LAD-HF diaphragm muscle at submaximal frequencies of stimulation (†P < 0.05 versus untreated LAD-HF diaphragm, *P < 0.05).
We chemically skinned and isolated single diaphragm muscle fibers to assess the difference in the force-calcium relationship between sham and LAD-HF diaphragms. Compared with sham diaphragm fibers, there was a significant rightward shift in the LAD-HF force-calcium relationship (sham DMSO versus LAD-HF DMSO: 5.57 ± 0.02 versus 5.5 ± 0.02; P < 0.05 by F test; Fig. 4B), indicating reduced calcium sensitivity in LAD-HF fibers. CK-2127107 (3 µmol/l) caused a significant leftward shift in the force-calcium relationship, restoring the calcium sensitivity lost to heart failure (Fig. 4B).
Given the atrophy and reduced calcium sensitivity in LAD-HF diaphragms, the isometric force output was measured in sham and LAD-HF diaphragms ex vivo by electrical field stimulation. The LAD-HF diaphragm muscle produced a lower force compared with sham diaphragms at stimulation frequencies ranging from 40 to 80 Hz (P < 0.05 by t test within each stimulation frequency; Fig. 4C). CK-2127107 (30 µmol/l) significantly increased force in both sham and LAD-HF diaphragms, with the effect being more prominent at stimulation frequencies less than or equal to 30 Hz (P < 0.05 by analysis of variance within each stimulation frequency; Fig. 4C). The leftward shift in the force-frequency relationship is consistent with the leftward shift observed in the force-calcium relationship of skinned muscle fibers (Fig. 4C).
CK-2127107 Significantly Improves Rotarod Performance in Exercise-Intolerant LAD-HF Rats.
Previous investigators demonstrated reduced exercise performance in an LAD-infarct model of heart failure (Pfeifer et al., 2001; Koh et al., 2003; Allen et al., 2008). We used an accelerating rotarod to assess exercise performance in sham and LAD-HF rats 16 weeks after coronary artery ligation. LAD-HF rats had a significantly shorter running time on an accelerating rotarod than sham rats (116 ± 22 versus 193 ± 31 seconds, P < 0.05, by repeated measures ANCOVA; Fig. 5), indicating a decrease in exercise performance. A single dose of CK-2127107 (10 mg/kg p.o.) increased performance time by approximately 150% in LAD-HF rats (116 ± 22 versus 283 ± 47 seconds, P = 0.0004, by repeated measures ANCOVA). There was no difference between sham and LAD-HF groups treated with CK-2127107 (sham versus LAD-HF was 274.8 ± 31 versus 283 ± 47 seconds).

CK-2127107 significantly increases rotarod performance in LAD-HF rats with exercise intolerance. LAD ligation resulted in a reduced rotarod running time compared with sham-operated control rats (193 ± 31 versus 116 ± 32 seconds; P = 0.042; mean ± S.E.). CK-2127107–treated sham rats ran longer than vehicle-treated sham rats (275 ± 31 versus 193 ± 31 seconds; P = 0.022). LAD-HF rats treated with CK-2127107 increased their rotarod running time approximately 2.5-fold compared with vehicle treatment (283 ± 47 seconds, P = 0.0004, by ANCOVA; *P < 0.05, ***P < 0.001), completely normalizing exercise capacity in LAD-HF rats.
Discussion
Chronic HF patients are often limited by breathlessness and muscular fatigue, and HF is typically associated with skeletal muscle dysfunction and exercise intolerance (Wilson et al., 1995). The extent of impaired ventricular systolic function in HF patients does not correlate well with the severity of dyspnea and exercise intolerance (Gibbs et al., 1990; Harrington and Coats, 1997). Therefore, improvements solely in cardiac function as a consequence of therapy may not necessarily reverse the existing skeletal muscle abnormalities and functional deficits in HF patients. Thus, the development of skeletal muscle–specific therapies may be useful in improving heart failure–mediated exercise intolerance.
Left anterior descending coronary artery ligation was used to produce myocardial infarction and consequent HF in rats, as this model has been shown to have significant reductions in exercise capacity (Yamaguchi et al., 1999; Pfeifer et al., 2001; Koh et al., 2003). Alterations in skeletal muscle Ca2+ sensitivity and handling have also been noted with heart failure (Ward et al., 2003; Kho et al., 2012b), and pharmacological calcium sensitizers have previously been shown to increase calcium sensitivity in diaphragms from heart failure rats (van Hees et al., 2011). In this study, LAD-HF rats exhibited significant and progressive heart dysfunction and subsequently demonstrated characteristics of skeletal myopathy, including myofiber atrophy and changes in myofibril calcium sensitivity and handling in diaphragm and hindlimb muscles at rest. Compared with sham rats, these skeletal muscle changes contributed to significant decreases in isometric diaphragm force output and increased hindlimb muscle fatigability in situ and exercise intolerance in vivo by rotarod assessment.
CK-2127107 is a novel small molecule fast skeletal troponin activator currently in human clinical trials for conditions of muscle weakness and fatigue. CK-2127107 increased sarcomeric Ca2+ sensitivity in skinned LAD-HF skeletal fibers in vitro, increased force output at submaximal stimulation frequencies in the diaphragm ex vivo and hindlimb muscle in situ, and significantly improved rotarod performance in LAD-HF rats in vivo, thus displaying efficacy on skeletal muscle function at multiple levels of investigation. These results demonstrated that increasing sarcomere Ca2+ sensitivity is a potentially unique therapeutic approach to ameliorate heart failure–mediated deficits in skeletal muscle strength and function.
One mechanism for fatigue in skeletal muscle is a reduction in sarcoplasmic reticular Ca2+ release and uptake that then results in decreased force production (Allen et al., 2008). Fatiguing exercise can impair SERCA function, which can affect both uptake and storage of calcium. In the absence of altered troponin calcium sensitivity, decreased Ca2+ release from the sarcoplasmic reticulum would lead to diminished troponin activation and subsequently diminished force production (Blanchard and Solaro, 1984; Nosek et al., 1987; Westerblad and Allen, 1996; Chin and Allen, 1998; Allen et al., 2008, 2011). The fatigue experienced by exercising heart failure patients is associated with an increased inorganic phosphate to phosphocreatine ratio and a lower intracellular pH relative to control subjects (Massie et al., 1987; Mancini et al., 1989), which conceivably affects sarcoplasmic Ca2+ release, sensitivity, and force output. In the current study, we observed an increase in muscle fatigue in situ and significantly reduced exercise performance time in LAD-HF rats. CK-2127107 reversed these deficits, suggesting that increasing troponin Ca2+ sensitivity can compensate for the presumed reduction in SERCA function in HF.
Inspiratory muscle weakness is noted in 30–50% of heart failure patients (Meyer et al., 2001; Ribeiro et al., 2012), and indices of diaphragm dysfunction and structural abnormalities have been noted in both patients and animal models of heart failure (Meyer et al., 2001; van Hees et al., 2008; Wong et al., 2011). LAD-HF rats in this study exhibited significant diaphragm atrophy and decreased sarcomeric Ca2+ sensitivity. CK-2127107 increased Ca2+ sensitivity and caused a leftward shift in the isometric force frequency relationship in LAD-HF diaphragm muscle. Patients with heart failure and inspiratory muscle weakness have an improved cardiorespiratory response to exercise following inspiratory muscle training (Winkelmann et al., 2009; Lin et al., 2012). Thus, much like the specific training of the respiratory muscles, it is possible that increased diaphragm muscle Ca2+ sensitivity by CK-2127107 may improve respiratory function during exercise and might thereby improve overall exercise tolerance.
The present study demonstrates that a single dose of CK-2127107 can substantially improve exercise performance in a model of mild to moderate heart failure. Increasing the calcium sensitivity of fast skeletal muscle with the attendant increases in responsiveness to neuromuscular input and fatigue resistance appear to underlie this improvement in exercise performance affected by CK-2127107. These findings suggest that skeletal muscle dysfunction rather than cardiac muscle dysfunction may play a greater role in exercise intolerance early in the course of heart failure. Overall, these findings have important implications for understanding how muscle fatigue and weakness contribute to reduced exercise performance in heart failure. They further suggest a potential therapeutic role for small molecule fast skeletal troponin activators as a means to improve skeletal muscle function and exercise performance in heart failure.
Authorship Contributions
Participated in research design: Hwee, Kennedy, Hartman, Ryans, Durham, Malik, Jasper.
Conducted experiments: Hwee, Kennedy, Hartman, Ryans, Durham.
Performed data analysis: Hwee, Kennedy, Hartman, Ryans, Durham, Malik, Jasper.
Wrote or contributed to the writing of the manuscript: Hwee, Kennedy, Hartman, Durham, Malik, Jasper.
Footnotes
- Received December 17, 2014.
- Accepted February 11, 2015.
D.T.H. and A.R.K. contributed equally to this work.
All research studies were financially supported by Cytokinetics.
↵
 This article has supplemental material available at jpet.aspetjournals.org.
This article has supplemental material available at jpet.aspetjournals.org.

Abbreviations
- ANCOVA
- analysis of covariance
- CK-2127107
- 2-aminoalkyl-5-N-heteroarylpyrimidine
- DMSO
- dimethyl sulfoxide
- EDL
- extensor digitorum longus
- HF
- heart failure
- ITC
- isothermal titration calorimetry
- LAD
- left anterior descending coronary artery
- SERCA
- sarcoendoplasmic reticulum Ca2+ ATPase
- Copyright © 2015 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}