


Abstract
Paraquat (N,N-dimethyl-4–4′-bipiridinium; PQ), a widely used herbicide, when ingested accidentally or intentionally can cause major organ toxicities in lung, liver, and kidney. Because PQ is primarily eliminated in the kidney, renal elimination, including tubular transport, plays a critical role in controlling systemic exposure to the herbicide. The goal of this study was to determine the molecular identities of the transporters involved in the renal elimination of PQ. Using stably transfected human embryonic kidney (HEK)-293 cells, we examined the role of human organic cation transporters (hOCTs, SLC22A1–3) and human multidrug and toxic compound extrusion (hMATE)1 in the cellular accumulation and cytotoxicity of PQ. We found that overexpression of hOCT2 but not hOCT1 and hOCT3 in HEK-293 cells significantly enhanced the accumulation and cytotoxicity of PQ (-fold increase for uptake was 12 ± 0.5, p < 0.01; -fold increase of cytotoxicity was 18 ± 1.5, p < 0.001). The kinetics of PQ transport was altered in cells expressing a genetic polymorphism of hOCT2 (A270S) in comparison with those expressing the reference hOCT2. In addition, the cellular accumulation and cytotoxicity of PQ were also enhanced in cells expressing hMATE1 (-fold increase for uptake was 18 ± 3.7, p < 0.0001; -fold increase of cytotoxicity was 5.7 ± 0.5, p < 0.0001). These results suggest that hOCT2 and hMATE1 mediate PQ transport. These transporters may play an important role in the accumulation and renal excretion of PQ, and they may serve as molecular targets for the prevention and treatment of PQ-induced nephrotoxicity.
Paraquat (N,N-dimethyl-4–4′-bipiridinium; PQ) is a potent herbicide widely used in more than 130 countries. Thousands of deaths each year are due to accidental or intentional ingestion of PQ. In addition, epidemiological studies have suggested that exposure to PQ may increase the risk for Parkinson's disease (Dinis-Oliveira et al., 2006). Upon oral administration, PQ accumulates to high concentrations in kidney, liver, and lung, resulting in damage to these organs. PQ is predominantly excreted by the kidney; therefore, renal excretion is a major determinant of the systemic exposure to PQ (Chan et al., 1998). Although numerous studies have suggested that organic cation transport systems are involved in active secretion of PQ, the molecular identities of these transporters have not been established. The goal of this study was to identify the specific transporters responsible for PQ renal secretion.
Renal secretion of organic cations is a process that involves an entry step at the basolateral membrane and an exit step at the apical membrane of renal tubular cells. Organic cation transporters (OCTs) in the SLC22A family are mainly expressed at the basolateral membrane and responsible for the entry of organic cations into the tubular cells (Wright, 2005). There are three distinct OCT transporters, including OCT1, OCT2, and OCT3. The OCTs mediate the cellular uptake of cationic compounds of various structures with molecular mass generally less than 400 g/mol (type I organic cations) (Jonker and Schinkel, 2004; Wright, 2005). The OCTs have overlapping substrates and inhibitors specificities. Common substrates of OCTs include the neurotoxin 1-methyl-4-phenylpyridinium (MPP+) and endogenous compounds 5-hydroxytryptamine, noradrenaline, histamine, and agmatine (Koepsell, 2004). In addition, OCTs interact with many drugs, including the anticancer drug oxaliplatin (Yonezawa et al., 2006; Zhang et al., 2006) and the antidiabetic drug metformin (Kimura et al., 2005). In humans, the predominant OCT in the kidney is OCT2 (Koepsell, 2004).
For many years, the transporters responsible for the flux of organic cations from the tubule cell to the tubule lumen were not known. Recently, using database searches, Otsuka et al. (2005) identified two human genes designated as hMATE1 and hMATE2, which encode orthologs of a newly classified multidrug resistance-conferring protein family in bacteria called multidrug and toxic compound extrusion (MATE). hMATE1 was found to be an organic cation/H+ exchanger mainly expressed in the apical membrane of renal tubule cells and in the canalicular membrane of hepatocytes. The transporter interacts with structurally diverse small-molecular-weight cations such as tetraethylammonium (TEA), MPP+, and metformin, and it is therefore thought to play a potential role in the renal and hepatic elimination of drugs, endogenous compounds, and environmental toxins. Recently, mouse MATE1 and rat MATE1 were cloned and functionally characterized (Hiasa et al., 2006; Terada et al., 2006). A kidney-specific hMATE2 (hMATE2-K) was also cloned and shown to transport various organic cations such as TEA, MPP+, cimetidine, metformin, and procainamide. These studies suggest that both hMATE2-K and hMATE1 play a role in the tubular transport of cationic drugs across the brush-border membrane (Masuda et al., 2006).
PQ is a divalent cation and has a molecular weight of 186.25. Because PQ is predominantly excreted in the kidney, we hypothesized that OCTs and MATEs may play an important role for PQ renal excretion. In the present study, we examined the interaction of PQ with the basolateral OCTs and the apical MATEs. We also characterized the interaction of PQ with nonsynonymous polymorphisms of hOCT2.
Materials and Methods
cDNAs. cDNAs of hMATE1 (GenBank accession no. NM_018242) and rMATE1 (GenBank accession no. BC088413.1) were amplified by reverse transcription-polymerase chain reaction from human or rat kidney RNA and cloned into pcDNA5-FRT (Invitrogen, Carlsbad, CA). The sequences of hMATE1 and rMATE1 were confirmed to be free of errors by comparing them with the human or rat genome sequence.
Mutagenesis. Variants of hOCT2 were constructed by site-directed mutagenesis using high-fidelity DNA polymerase PfuTurbo (Stratagene, La Jolla, CA) according to manufacturer's protocol. The variants were fully sequenced to verify that they had only the desired mutation.
Cell Culture and Transfection. Human embryonic kidney (HEK)-293-Flp-In cells (Invitrogen) were transfected with pcDNA5/FRT empty vector or with pcDNA5/FRT vector containing the full-length hMATE1, rMATE1, hOCT2, and hOCT2 variants using Lipofectamine 2000 (Invitrogen). Forty eight hours after transfection, 75 μg/ml hygromycin B was added to the medium, and stable clones were selected. HEK-293 cells stably transfected with hOCT1 and hOCT3 were established previously in our laboratory (Zhang et al., 2006). The culture medium for stably transfected HEK-293 cells (except hOCT3) was Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum, 100 unit/ml penicillin, 100 μg/ml streptomycin and 60 μg/ml hygromycin B. All cell lines were grown at 37°C in a humidified atmosphere with 5% CO2. The culture medium for hOCT3 stably transfected cells was Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum, 100 U/ml penicillin, 100 μg/ml streptomycin, and 600 μg/ml neomycin.
Cytotoxicity Assay. The cytotoxicity of PQ was measured by the MTT assay. In brief, cells were seeded in biocoated 96-well plates. The next day, cells were exposed to different concentrations of PQ for 6 h. Drug-containing medium was replaced with fresh medium, and cells were incubated for another 66 h. MTT reagent was added and incubated for 4 h. The purple formazan was dissolved with 0.1 N HCl in isopropanol, and the samples were read using a microplate reader at a wavelength of 570 nm. The IC50 values were obtained by fitting F, the percentage of the maximal cell growth at different drug concentrations, to the equation F = 100 × [1 – Cγ/(IC50γ + Cγ)] using WinNonlin (Pharsight, Mountain View, CA). The maximal cell growth was the cell growth in the medium without PQ; C is the concentration of PQ, and γ is the slope factor.
Cellular Uptake of PQ. HEK-293 stably transfected cells were grown in monolayer in 24-well plate. When cells reach more than 90% confluence (generally within 24 h), they were incubated in buffer containing 125 mM NaCl, 4.8 mM KCl, 5.6 mM d-glucose, 1.2 mM CaCl2, 1.2 mM KH2PO4, 1.2 mM MgSO4, and 25 mM Tricine, pH 8.0. Transport assays were initiated by adding 4.5 μM[14C]PQ. The uptake was stopped after the indicated time by washing cells three times with ice-cold choline buffer containing 128 mM choline, 4.73 mM KCl, 1.25 mM CaCl2, 1.25 mM MgSO4, and 5 mM HEPES/Tris, pH 7.4. Cells were lysed and incubated with 0.1 N NaOH/0.1% SDS for 1 h. Radioactivity in aliquot of cell lysates was determined by scintillation counting. Protein was measured by BCA protein assay (Pierce Chemical, Rockford, IL). For kinetic studies, varying amounts of unlabeled PQ were added to the uptake solutions to give increasing total (radiolabeled plus unlabeled) substrate concentrations, typically ranging from 5 to 1000 μM. Nonspecific cell-associated radioactivity was determined by measuring substrate uptake in mock cells at each substrate concentration, and these values were then subtracted from the results in hOCT2- or MATE1-transfected cells to give the final kinetic data. The Km and Vmax values were obtained by fitting the Michaelis-Menten equation V = Vmax × [S]/(Km + [S]) using WinNonlin, where V refers to the rate of substrate transport, Vmax refers to the maximum rate of substrate transport, [S] refers to the concentration of substrate, and Km is defined as the concentration of substrate at the half-maximal transport rate. For inhibition studies, different concentrations of unlabeled n-tetra-alkylammoniums (nTAAs) were added to uptake solution containing 4.5 μM[14C]PQ. The IC50 value was estimated by fitting F, the percentage of the maximal uptake at different drug concentrations, to the equation F = 100 × [1 – Cγ/(IC50γ + Cγ)] using WinNonlin. The maximal uptake was the cell uptake of PQ without any nTAAs. C is the concentration of different nTAAs, and γ is the slope factor.
Statistical Analysis. Data are expressed as mean ± S.D. Unpaired Student's t test was used to analyze differences between two groups. Multiple comparisons were analyzed using one-way analysis of variance followed by Dunnett's two-tailed test. The data were analyzed with GraphPad Prism 4.0 (GraphPad Software Inc., San Diego, CA). A p value less than 0.05 is considered statistically significant.
Results
Transport of PQ in HEK-293 Cells Stably Transfected with hOCT1, hOCT2, and hOCT3. We first examined whether PQ was a substrate of the human organic cation transporters hOCT1, hOCT2, and hOCT3. As shown in Fig. 1A, the uptake of PQ in HEK-hOCT1 and HEK-hOCT2 cells was 1.9 ± 0.1- (p < 0.01) and 12.3 ± 0.6 (p < 0.01)-fold greater than that in HEK-Mock cells, respectively. However, the uptake of PQ in HEK-hOCT3 was comparable with that in HEK-Mock cells (0.62 ± 0.04; p > 0.05). These stably transfected hOCT1, hOCT3, and hOCT3 cells were shown to transport the model substrate MPP+ (data not shown). These data suggest that PQ is an excellent substrate of hOCT2 and a weak substrate of hOCT1, but it is not a substrate of hOCT3. The finding that PQ is a poor substrate of hOCT1, a liver-specific OCT, suggests that PQ accumulation in the liver involves other transporters. In addition, the hOCT2-expressing cells also exhibited a time-dependent uptake of PQ, whereas the mock control cells did not (Fig. 1B). The kinetics of interaction of hOCT2 with PQ was saturable with a Km value of 114 ± 23 μM and Vmax value of 174 ± 37 pmol/mg/min (mean ± S.D.) (Fig. 1C).

Transport of PQ in HEK-293 cells stably transfected with hOCT1, hOCT2, and hOCT3. A, stably transfected HEK-Mock, HEK-hOCT1, -hOCT2, or -hOCT3 cells were incubated with 4.5 μM[14C]PQ for 10 min, and uptake of PQ in these cells was determined. B, time course of 4.5 μM PQ uptake by HEK-293 cells expressing hOCT2 or a mock control. C, kinetics of PQ uptake. HEK-hOCT2 or HEK-Mock were incubated with 4.5 μM[14C]PQ as well as different concentrations of unlabeled PQ for 10 min. Nonspecific cell-associated radioactivity was determined by measuring substrate uptake in mock cells at each substrate concentration, and these values were then subtracted from the values in hOCT2-transfected cells to give the final kinetic curves. Studies were performed in triplicate in each individual experiment. Data are from three separate experiments (mean ± S.D.).
Transport of PQ by hOCT2 and Its Genetic Variants. Four common nonsynonymous coding region variants of hOCT2 (M165I, A270S, R400C, and K432Q) were identified previously in our laboratory (Leabman et al., 2002). To examine the interaction of genetic polymorphisms of hOCT2 with PQ, we constructed stably transfected HEK-293 cells expressing the empty vector, the reference hOCT2, and the four common variants. The interaction of the variants of hOCT2 with PQ was saturable as shown in Fig. 2. The Vmax and Km values of PQ are shown in Table 1 for the reference OCT2 and all four variants. We observed that the average values of Vmax and Km for M165I, R400C, and K432Q were not significantly different from those of the reference OCT2 (p > 0.05). However, the average Vmax and Km value for PQ in cells expressing A270S variant was 3-fold greater than in cells expressing OCT2 reference (p < 0.01 for Vmax and p < 0.05 for Km, respectively).
Kinetic values of PQ uptake in HEK-293 cells expressing hOCT2 reference or variants
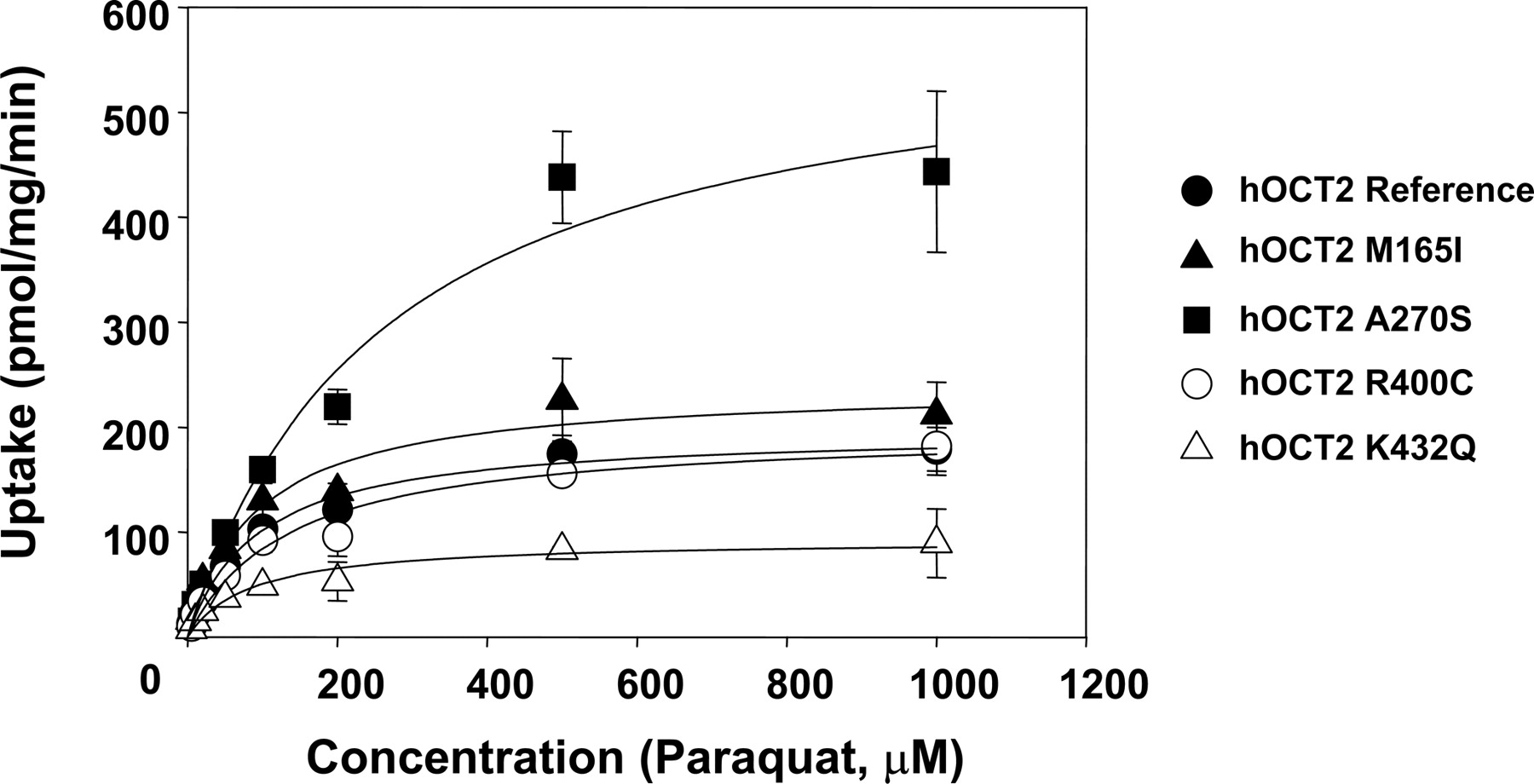
Transport of PQ by hOCT2 and its genetic variants. HEK-293 cells stably transfected with empty vector, hOCT2 reference, M165I, A270S, R400C, and K432Q were incubated with 4.5 μM [14C]PQ as well as different concentrations of unlabeled PQ for 10 min. Final uptake values were obtained by subtracting the uptake in mock cells from that in the cells expressing hOCT2 reference or variants at each corresponding substrate concentration. Studies were performed in triplicate in each individual experiment. Data are from three separate experiments (mean ± S.D.). Kinetic values listed in the Table 1.
Transport of PQ by hMATE1 and rMATE1. The interaction of the apical membrane transporter hMATE1 with PQ was examined in stably transfected HEK-293 cells. As shown in Fig. 3A, PQ accumulated in the hMATE1-expressing cells with time, whereas no apparent accumulation was observed in the empty vector-transfected cells, suggesting that PQ is a substrate of hMATE1. The uptake of PQ by hMATE1 was saturable with a Km value of 212 ± 19 μM and Vmax value of 289 ± 46 pmol/min/mg protein (Fig. 3B). As shown in Fig. 3C, the rat ortholog rMATE1 also exhibited a time-dependent uptake of PQ, whereas the mock control cells did not. The uptake of PQ by rMATE1 was saturable with a Km value of 170 ± 22 μM and a Vmax value of 723 ± 76 pmol/min/mg protein (Fig. 3D). The Vmax value of PQ in rMATE1-expressing cells was 2.5-fold greater than that in cells expressing hMATE1 (p < 0.01). The Km value of PQ in rMATE1-expressing cells was not significantly different from that in cells expressing hMATE1 (p > 0.05).

Transport of PQ by hMATE1 and rMATE1. A and C, time course of 4.5 μM PQ uptake by HEK-293 cells expressing hMATE1, rMATE1, or a mock control. Stably transfected HEK-Mock, HEK-hMATE1, or HEK-rMATE1 cells were incubated with 4.5 μM[14C]PQ for different times, and uptake of PQ in these cells was determined. B and D, concentration-dependent PQ uptake in hMATE1- or rMATE1-expressing cells. HEK-Mock, HEK-hMATE1, or HEK-rMATE1 were incubated with 4.5 μM [14C]PQ as well as different concentrations of unlabeled PQ for 10 min. Final uptake values were obtained by subtracting the uptake in mock cells from that in hMATE1 or rMATE1 cells at each corresponding substrate concentration. Studies were performed in triplicate in each individual experiment. Data are from three separate experiments (mean ± S.D.).
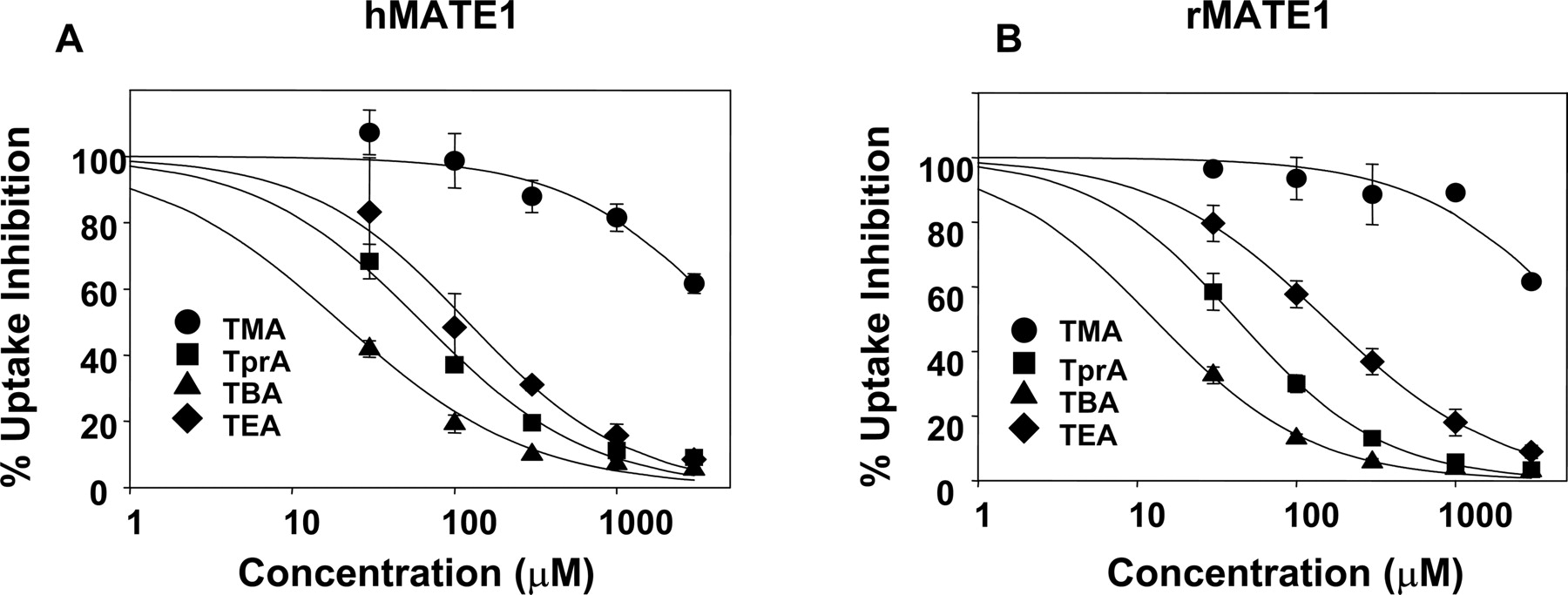
Inhibition of MATE1-Mediated Uptake of PQ by nTAA Compounds. Previously, we observed pronounced differences in the specificities and kinetics of interaction of nTAA for the human paralogs hOCT1 and hOCT2 (Dresser et al., 2002). Here, we determined the interaction of nTAAs with the rat and human orthologs of MATE1. As shown in Fig. 4A and Table 2, hMATE1-mediated PQ uptake was decreased by increasing concentrations of nTAAs. Tetrabutylammonium showed the highest potency of inhibition, followed by tetrapropylammonium, TEA, and tetramethylammonium. PQ uptake by the rat ortholog rMATE1 was inhibited by nTAAs in a similar pattern (Fig. 4B; Table 2). There was no significant difference in terms of affinity for hMATE1 or rMATE1 for each individual compound, suggesting little species differences in the inhibition kinetics of alkylammonium compounds with rodent and human MATE1.
IC50 value (micromolar) for each nTAA to inhibit PQ uptake in HEK-293 cells expressing either hMATE1 or rMATE1
Cytotoxicity of PQ in HEK-hOCT2 and HEK-hMATE1. To determine whether the transporters could potentiate PQ toxicity, we examined the cytotoxicity of PQ in cells expressing hOCT2 and hMATE1. The IC50 value of PQ in HEK-Mock cells and HEK-hOCT2 cells were 417 ± 60 and 23 ± 2 μM, respectively (18-fold; p < 0.001), indicating that expression of hOCT2 greatly enhances PQ-induced cytotoxicity (Fig. 5A). The IC50 values of PQ in HEK-Mock cells and HEK-hMATE1 cells were 717 ± 51 and 125 ± 5 μM, respectively (5.7-fold; p < 0.0001), indicating that hMATE1 also enhances, although to a lesser extent, PQ-induced cytotoxicity (Fig. 5B).

Inhibition of MATE1-mediated uptake of PQ by nTAAs. HEK-hMATE1 cells (A) or HEK-rMATE1 cells (B) were incubated with 4.5 μM [14C]PQ as well as varying amounts of nTAAs for 10 min. Studies were performed in triplicate in each individual experiment. Data are from two separate experiments (mean ± S.D.). IC50 values for each nTAA is listed in Table 2.
Discussion
PQ is one of the most toxic herbicides. PQ causes death after large oral doses (>40 mg/kg) by producing multiorgan failure. The major organs affected by the agent are the kidney, liver, and lung, which accumulate high concentrations of PQ. PQ is a small divalent cation predominantly eliminated by the kidney in individuals with normal renal function (Chan et al., 1998). Therefore, it is crucial to maintain renal function in treating patients with PQ poisoning to facilitate its elimination. However, the precise mechanism(s) by which the kidney eliminates PQ remains unclear.
The renal clearance of PQ was reported to exceed that of glomerular filtration rate in different species, suggesting that PQ is actively secreted in the kidney (Chan et al., 1998). Cellular studies indicate that PQ is transported in the renal epithelial cell line derived from proximal tubule, LLC-PK1, but to a much lesser extent in the distal tubule derived cell line Madin-Darby canine kidney (Chan et al., 1996b, 1997a), suggesting that the secretion of PQ occurs in the proximal tubule.
Early studies by Wright and coworkers suggested that the transport of PQ across the luminal membrane of the proximal tubule involved an organic cation/H+ exchange mechanism (Wright and Wunz, 1995). Using rat renal proximal tubular cells in primary culture, Chan et al. (1996a) showed that PQ was predominantly taken up from the basolateral side by a saturable, energy-dependent mechanism that could be inhibited by quinine and TEA. Chan et al. (1997b) further demonstrated that the renal clearance of PQ was greater than glomerular filtration rate and that quinine and N-methylnicotinamide reduced the fractional excretion of PQ in anesthetized rats. These studies are consistent with the notion that PQ is actively secreted by the rat kidney via a cation transport system.
In the present study, we tested basolateral and apical membrane transporters, OCTs and MATEs, respectively, to determine the molecular mechanisms responsible for PQ elimination in the kidney. hOCT2 is the major OCT in human kidney, and it is expressed on the basolateral membrane (Jonker and Schinkel, 2004; Koepsell, 2004). Our data indicated that PQ is a good substrate of hOCT2 (Fig. 1), suggesting that hOCT2 may mediate PQ uptake from the circulation into the human kidney. The transporter has previously been found to interact mainly with monovalent organic cations (Jonker and Schinkel, 2004). Our finding that PQ, a divalent organic cation, is also a substrate of hOCT2 suggests that the transporter has a broader specificity than previously thought. Although speculative, the data suggest that hOCT2 may accept two organic cations simultaneously, and they indicate the need for more detailed studies of specificity and mechanism of the transporter. Furthermore, the finding that unlike hOCT2, hOCT1 and hOCT3 do not interact well with PQ is consistent with differences in the specificity of OCT orthologs (Zhang et al., 2006) and the idea that OCTs, which have distinct tissue distributions, serve different physiological roles in the body. Irrespective of mechanism, the data suggest that hOCT2 contributes to and may be the predominant mechanism for the uptake of PQ by the renal proximal tubule.

Effect of hOCT2 or hMATE1 on PQ cytotoxicity. HEK-hOCT2 (A), HEK-hMATE1 (B), or mock cells were incubated with different concentrations of PQ for 6 h. Medium was removed and replaced with drug-free fresh medium for another 66 h. MTT assays were performed. Studies were performed in quadruplicate in each individual experiment. Data are from three separate experiments (mean ± S.D.).
With the cloning of MATEs, it is now possible to determine the molecular identity of apical membrane organic cation transporters that are responsible for the second step in renal organic cation secretion. As shown in Fig. 3, PQ was transported by both hMATE1 and rMATE1 in a time- and dose-dependent manner, suggesting that MATE1 contributes to the transport of PQ across the apical membrane. It is noteworthy that the data suggest that in addition to transporting monovalent organic cations, MATE1 can transport divalent cations. In addition, hMATE2 and kidney-specific hMATE2 (hMATE2-K) were shown to be expressed in the kidney (Otsuka et al., 2005; Masuda et al., 2006). Our data indicated that PQ was not a substrate of hMATE2 (data not shown). Whether PQ is a substrate of hMATE2-K remains to be determined. The finding that there was a relationship between molecular size of the nTAAs and potency of inhibition of PQ uptake by human and rat orthologs of MATE1 is consistent with the idea that increasing hydrophobicity is associated with greater potency of inhibition of hMATE1. Interestingly, we observed a similar relationship between inhibition potency and hydrophobicity for OCT1, but not for OCT2 (Dresser et al., 2002). However, other studies have shown a positive correlation between hydrophobicity and increasing interaction with OCT2 (Arndt et al., 2001; Suhre et al., 2005). Collectively, these studies suggest that a hydrophobic organic cation may act at either or both the basolateral and the luminal membrane to inhibit tubular transport of organic cations.
PQ is a nephrotoxic agent. Exposure to PQ can cause morphological change in proximal renal tubules, which subsequently lead to renal functional change (Chan et al., 1998). In this study, we observed that overexpression of hOCT2 not only increased the uptake of PQ (Fig. 1) but also enhanced the cytotoxicity of the agent (Fig. 5), suggesting that the function of hOCT2 may play an important role in the modification or prevention of PQ-induced nephrotoxicity. It is well recognized that genetic variations in drug transporters may alter drug disposition and subsequent clinical drug response (Leabman et al., 2003). Leabman et al. (2003) identified 28 variable sites by screening all 11 exons as well as flanking intronic sequences of hOCT2 from a collection of 247 ethnically diverse DNA samples. They also determined the activity of four polymorphic nonsynonymous variants, M165I, A270S, R400C, and K432Q, and they found that all four variants altered the transporter function assayed in Xenopus laevis oocytes heterologously expressing the variant transporters (Leabman et al., 2002). Recently, Lazar et al. (2006) reported that patients with 270S allele of hOCT2 were less frequently affected by the clinical diagnosis of hypertension than the homozygous A270, suggesting a significant association of the A270S polymorphism with essential hypertension in their patients samples. The mechanism for this association is not known. Zhang et al. developed a structural model using the GlpT crystal structure (1PW4) as a template (Zhang et al., 2005; Pelis et al., 2006). Here, we generated a model with the same template using Modeler (Fiser and Sali, 2003). We found that the A270S variant may have two effects. First, the substitution is near a neighboring tyrosine, so the addition of an OH group may therefore alter the tertiary structure. Second, the change substitutes a nonpolar residue with a polar residue. A270 partially resides on the model's protein surface. Nonpolar amino acids tend to reside in the center of a protein where they can interact with similar nonpolar neighbors, whereas polar side chains (e.g., Serine) tend to be present on the surface of a protein where they can interact with the aqueous environment. Interestingly, in this study we observed that A270S had an increased maximal transport rate (Vmax) of PQ in stably transfected HEK cells, suggesting that this common variant found at allele frequencies of 10 to 20% in ethnic populations may be associated with altered renal transport of PQ in humans. One study by Lee et al. (1999) showed that the plasma PQ levels of victims who died within 1 day of ingestion ranged from 12.3 to 3417 μM (average 685 μM). Our data suggest that individuals who carry OCT2-A270S may exhibit altered toxicity to PQ. At high concentrations of PQ, hOCT2 would be functioning at maximal rates of transport. Those individuals with A270S may clear the herbicide more quickly than those with the reference hOCT2. The identification of genetic variants of hMATE1 is ongoing, and the role of genetic variations of hMATE1 in PQ transport will be determined.
Based on the findings of this study, we propose that hOCT2 contributes to the first step in the renal tubule secretion of PQ and that hMATE1 contributes to the second step. The contribution of these transporters in vivo to the renal clearance and nephrotoxicity of PQ needs to be examined in studies using animal models. Although other transporters may also play a role in the elimination of PQ in the kidney, hOCT2 and hMATE1 may serve as potential molecular targets for the prevention and treatment of PQ-induced nephrotoxicity.
Footnotes
-
This work was supported by National Institutes of Health Grants GM36780, GM61390, GM074929, and ES12077. Y.C. was supported by National Research Service Award T32 GM07546 from the National Institutes of Health.
-
Article, publication date, and citation information can be found at http://jpet.aspetjournals.org.
-
doi:10.1124/jpet.107.123554.
-
ABBREVIATIONS: PQ, paraquat; OCT, organic cation transporter; MPP+, 1-methyl-4-phenylpyridinium; MATE, multidrug and toxic compound extrusion; h, human; TEA, tetraethylammonium; HEK, human embryonic kidney; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium; nTAA, n-tetraalkylammonium; r, rat.
- Received March 28, 2007.
- Accepted May 9, 2007.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}