


Abstract
Adenosine modulates the immune system and inhibits inflammation via reduction of cytokine biosynthesis and neutrophil functions. Drugs able to prevent adenosine catabolism could represent an innovative strategy to treat inflammatory bowel disorders. In this study, the effects of 4-amino-2-(2-hydroxy-1-decyl)pyrazole[3,4-d]pyrimidine (APP; novel adenosine deaminase inhibitor), erythro-9-(2-hydroxy-3-nonyl)adenine hydrochloride (EHNA; standard adenosine deaminase inhibitor), and dexamethasone were tested in rats with colitis induced by 2,4-dinitrobenzenesulfonic acid (DNBS). DNBS-treated animals received APP (5, 15, or 45 μmol/kg), EHNA (10, 30, or 90 μmol/kg), or dexamethasone (0.25 μmol/kg) i.p. for 7 days starting 1 day before colitis induction. DNBS caused bowel inflammation associated with decrease in food intake and body weight. Animals treated with APP or EHNA, but not dexamethasone, displayed greater food intake and weight gain than inflamed rats. Colitis induced increment in spleen weight, which was counteracted by all test drugs. DNBS administration was followed by macroscopic and microscopic inflammatory colonic alterations, which were ameliorated by APP, EHNA, or dexamethasone. In DNBS-treated rats, colonic myeloperoxidase, malondialdehyde, and tumor necrosis factor (TNF)-α levels as well as plasma TNF-α and interleukin-6 were increased. All test drugs lowered these phlogistic indexes. Inflamed colonic tissues displayed an increment of inducible nitric-oxide synthase mRNA, which was unaffected by APP or EHNA, but reduced by dexamethasone. Cyclooxygenase-2 expression was unaffected by DNBS or test drugs. These findings indicate that 1) inhibition of adenosine deaminase results in a significant attenuation of intestinal inflammation and 2) the novel compound APP is more effective than EHNA in reducing systemic and intestinal inflammatory alterations.
Inflammatory bowel diseases (IBDs) are severe conditions that produce a wide range of gastrointestinal and extradigestive symptoms, with great impact on the quality of life of the patient (Hanauer and Present, 2003). IBDs can be the consequence of abnormal immune reactions against luminal bacteria in genetically susceptible individuals (Sartor, 2004). Several cytokines, such as tumor necrosis factor (TNF)-α, interleukin (IL)-1, and IL-6, contribute to their pathogenesis (Ardizzone and Bianchi Porro, 2005; Pizarro et al., 2006), and the resulting inflammatory responses are mediated predominantly by activated neutrophils and macrophages. TNF-α regulates the expression of adhesion molecules and promotes the infiltration of inflammatory cells (Sasaki et al., 2004). IL-6 is a pleiotropic cytokine with a major role in IBDs (Scheller et al., 2006), suggested to be a relevant serologic marker correlated with inflammatory activity (Reinisch et al., 1999). IBDs are also characterized by extensive inflammatory infiltrates in the lamina propria, consisting of polymorphonuclear neutrophils, eosinophils, and plasma cells, leading to a remarkable production of unstable chemical species (i.e., reactive oxygen and nitrogen species), significantly implicated in tissue injury (Oz et al., 2005; Martin and Wallace, 2006).
At present, pharmacotherapy represents the mainstay of IBD management (Stein and Hanauer, 1999). It is noteworthy that some anti-inflammatory/immunomodulating drugs, including salicylates and methotrexate, are able to decrease intracellular adenosine 5′-triphosphate concentrations and raise extracellular adenosine levels, and it has been proposed that such property can significantly contribute to their therapeutic actions in inflammatory diseases (Cronstein et al., 1999).
Adenosine is a purine nucleoside that, once released from cells or being formed extracellularly, diffuses to the surface of surrounding cells, where it binds specific membrane receptors (Ralevic and Burnstock, 1998). Although adenosine is present at low concentrations in the extracellular space, stressful conditions, such as inflammation, can markedly increase its extracellular levels (Kuno et al., 2006). Recent evidence indicates that adenosine helps to maintain tissue integrity by reducing energy demand, increasing nutrient availability, and modulating the immune system (Siegmund et al., 2001). Accordingly, adenosine can play a beneficial role as immunomodulator in tissues subjected to injurious stimuli, including ischemia and inflammation (Haskó and Cronstein, 2004).
Several reports have demonstrated the ability of adenosine to exert anti-inflammatory actions in a variety of experimental models. In particular, this nucleoside can interfere with the biosynthesis of proinflammatory cytokines (Naganuma et al., 2006), and it down-regulates neutrophil functions, decreasing their adhesion, degranulation, and oxidant activity (Eigler et al., 2000; Haskó and Cronstein, 2004). When considering intestinal inflammation, Siegmund et al. (2001) demonstrated an amelioration of dextran sulfate-induced colitis in mice through an increase in endogenous adenosine levels evoked by inhibition of adenosine kinase, the enzyme responsible for conversion of adenosine to adenosine monophosphate. Odashima et al. (2005) observed that the stimulation of A2a adenosine receptors can reduce the severity of experimental ileitis in rabbits. More recently, Cavalcante et al. (2006) demonstrated that the new A2a receptor agonist ATL 313 reduced tissue injury and inflammation in mice with enteritis induced by Clostridium difficile toxin A. Based on these findings, the adenosine system may represent a promising target for therapies of inflammatory intestinal disorders. The aim of the present study was to evaluate whether the pharmacological modulation of adenosine deaminase, the enzyme responsible for adenosine inactivation by conversion into inosine, can affect bowel inflammation. For this purpose, the effects of 4-amino-2-(2-hydroxy-1-decyl)pyrazole[3,4-d]pyrimidine (APP), a novel adenosine deaminase inhibitor (Da Settimo et al., 2005), and erythro-9-(2-hydroxy-3-nonyl)adenine hydrochloride (EHNA), a reference adenosine deaminase inhibitor (Bessodes et al., 1982), were assayed in a rat model of experimental colitis.
Materials and Methods
Adenosine Deaminase Inhibition Assay. In vitro activity of adenosine deaminase was determined spectrophotometrically by monitoring for 2 min at 262 nm the decrease in absorbance resulting from deamination of adenosine into inosine by the enzyme (Da Settimo et al., 2005). Variations in adenosine concentration per minute were determined by means of a Beckman DU-64 kinetics software program (Solf Pack Module; Beckman Coulter, Fullerton, CA). The enzyme activity was assayed at 30°C in a reaction mixture containing 50 μM adenosine, 50 mM potassium phosphate buffer, pH 7.2, and 0.3 nM adenosine deaminase in a total volume of 500 μl. The inhibitory activity of EHNA and APP was assayed by adding 100 μl of drug solution to the reaction mixture to obtain final concentrations ranging from 0.05 to 50 nM. To correct for absorption by test compounds, reference blanks containing adenosine deaminase, potassium phosphate buffer, and appropriate concentrations of EHNA and APP were also prepared. Indeed, to correct for nonenzymatic changes in adenosine concentration, reference blanks containing all the above assay components, with the exception of substrate, were prepared. Each drug concentration was tested in triplicate, and the determination of IC50 values was performed by linear regression analysis of the log concentration-response curve. Ki values were calculated from IC50 by means of the Cheng-Prusoff equation.
Animals. Albino male Sprague-Dawley rats (250–300 g body weight) were employed throughout the study. The animals were fed standard laboratory chow and tap water ad libitum and were not subjected to experimental procedures for at least 1 week after their delivery to the laboratory. Their care and handling were in accordance with the provisions of the European Union Council Directive 86-609, recognized and adopted by the Italian Government.
Induction of Colitis and Drug Treatments. Colitis was induced in accordance with the method described previously by Barbara et al. (2000) with minor changes. In brief, during a short anesthesia with isoflurane (Abbott, Rome, Italy), 15 mg of 2,4-dinitrobenzenesulfonic acid (DNBS) in 0.25 ml of 50% ethanol was administered intrarectally via a polyethylene PE-60 catheter inserted 8 cm proximal to the anus. Control rats received 0.25 ml of 50% ethanol. Animals underwent subsequent experimental procedures 6 days after DNBS administration to allow a full development of histologically evident colonic inflammation.
Test drugs were administered i.p. for 7 days, starting 1 day before the induction of colitis. Animals were assigned to the following treatment groups: EHNA (90 μmol/kg), APP (15 μmol/kg), or dexamethasone (0.25 μmol/kg). DNBS-untreated animals (control group) and DNBS-treated rats (DNBS group) received drug vehicle to serve as controls. Food intake and body weight were monitored daily starting from the onset of drug treatments.
To select appropriate doses of test drugs, the potencies of EHNA and APP in blocking adenosine deaminase activity were assessed in vitro by an enzyme inhibition assay, performed as reported above. A reference in vivo dose of EHNA (30 μmol/kg) was selected on the basis of a previous study evaluating the anti-inflammatory effects of this drug in a rat model of asthma (Ellis et al., 2003). Effective doses of APP and EHNA were then selected by preliminary experiments designed to assay increasing doses of these compounds (APP, 5, 15, and 45 μmol/kg; EHNA, 10, 30, and 90 μmol/kg) on food intake, body weight, spleen weight, and macroscopic score in the model of DNBS-induced colitis. The dose of dexamethasone was selected on the basis of a previous study performed on a rat model of colitis (Nakase et al., 2001). Macroscopic score was evaluated on the whole colon, whereas microscopic and biochemical analysis were performed on specimens taken from a region of inflamed colon immediately adjacent and distal to the gross necrotic damage.
Assessment of Colitis. At the end of treatments, colonic tissues were excised, rinsed with saline, and scored for macroscopic and histological damage, in accordance with the criteria previously reported by Fornai et al. (2006). The macroscopic damage was scored on a 0- to 6-point scale based on the system reported in Table 1. The presence of adhesions between colonic tissue and other organs (0, none; 1, minor; 2, major adhesions) and the consistency of colonic fecal material (0, formed; 1, loose; 2, liquid stools) were also scored. Microscopic damage and inflammation were assessed by light microscopy on hematoxylin/eosin-stained histological sections obtained from whole gut specimens. Histological criteria included: mucosal architecture loss (0–3), cellular infiltrate (0–3), muscle thickening (0–3), crypt abscess (0, absent; 1, present), and goblet cell depletion (0, absent; 1, present). All parameters of macroscopic and histological damages were recorded and scored for each rat by two observers blinded to the treatment. At the time of experiment, the weight of spleen was also measured.
Criteria for macroscopic scoring of colonic ulceration and inflammation
Score was increased of 1 for each millimeter of bowel wall thickness.
Determination of Tissue Myeloperoxidase. Myeloperoxidase (MPO) levels in colonic tissues were determined as previously reported by Pacheco et al. (2000) and assumed as a quantitative index to estimate the degree of mucosal infiltration by polymorphonuclear cells. In brief, colonic samples (300 mg) were homogenized three times (30 s each) at 4°C with a Polytron homogenizer (Cole Parmer Homogenizer, Vernon Hills, IL) in 1 ml of ice-cold 50 mM phosphate buffer, pH 6.0, containing 0.5% of hexadecyltrimethylammonium bromide to prevent the pseudoperoxidase activity of hemoglobin as well as to solubilize membrane-bound MPO. The homogenate was sonicated for 10 s, frozen-thawed three times, and spun by centrifugation for 20 min at 18,000g. The supernatant was then recovered and used for determination of MPO by means of a kit for enzymelinked immunosorbent assay (Bioxytech; Oxis International Inc., Portland, OR). All samples were assayed within 2 days from collection. The results were expressed as nanograms of MPO per 100 mg of tissue.
Evaluation of Tissue Malondialdehyde. Malondialdehyde (MDA) concentration in colonic specimens was evaluated to obtain quantitative estimation of membrane lipid peroxidation. Colonic tissues were weighed, minced by forceps, homogenized in 2 ml of cold buffer (20 mM Tris-HCl, pH 7.4) by a Polytron homogenizer (Cole Palmer Homogenizer), and spun by centrifugation at 1500g for 10 min at 4°C. Colonic MDA concentrations were determined by means of a kit for colorimetric assay (Calbiochem-Novabiochem Corporation, San Diego, CA), and the results were expressed as micromoles of MDA per milligram of colonic tissue.
Cytokine Assays. At the time of sacrifice, plasma from trunk blood was collected for determination of TNF-α and IL-6 by means of kits for enzyme-linked immunosorbent assay (R&D Systems, Minneapolis, MN). The results were expressed as picograms per milliliter. Tissue TNF-α levels were also measured using a kit for enzymelinked immunosorbent assay (Biosource International, Camarillo, CA). For this purpose, as described by Marquez et al. (2006), tissue samples, previously stored at–80°C, were weighed, thawed, and homogenized in 0.3 ml of phosphate-buffered saline, pH 7.2/100 mg of tissue at 4°C, and centrifuged at 13,400g for 20 min. One hundred-microliter aliquots of the supernatants were then used for assay. Tissue TNF-α levels were expressed as picograms per milligram of tissue.
Reverse Transcription-Polymerase Chain Reaction. Expression of mRNA for inducible nitric-oxide synthase (iNOS) and cyclooxygenase (COX)-2 was assessed by reverse transcription (RT)-polymerase chain reaction (PCR). The analysis was performed on colonic specimens excised as reported above, snap frozen in liquid nitrogen, and stored at–80°C. At the time of extraction, tissue samples were disrupted with cold glass pestles, and total RNA was isolated by TRIzol (Invitrogen Srl, Milan, Italy) and chloroform. Total RNA (4 μg) served as template for cDNA synthesis in a reaction using 2 μlof random hexamers (0.5 μg/μl) with 200 U of Moloney murine leukemia virus-reverse transcriptase in a buffer containing 500 μM deoxynucleotide triphosphate (dNTP) and 10 mM dithiothreitol. cDNA samples were subjected to PCR in the presence of primers based on cloned rat iNOS and COX-2 (Virdis et al., 2005; Fornai et al., 2006). PCR, consisting of 5 μl of RT products, 2.5 U of Taq polymerase, 100 μM dNTP, and 0.5 μM primers, was carried out by a PCR-Express thermocycler (Hybaid Ashford, Middlesex, UK). For iNOS, the cycle condition was 1 min at 94°C, 1 min at 55°C, and 1 min at 72°C for 30 cycles, followed by 7 min at 72°C. For COX-2, the amplification conditions were: 1 min at 94°C, 2 min at 60°C, and 1 min at 72°C for 35 cycles followed by 7 min at 72°C. Aliquots of RNA not subjected to RT were included in PCR reactions to verify the absence of genomic DNA. The efficiency of RNA extraction, RT, and PCR was evaluated by primers for rat β-actin. PCR products were separated by 1.8% agarose gel electrophoresis in a 40 mM Tris buffer containing 2 mM EDTA and 20 mM acetic acid, pH 8, and stained with ethidium bromide. PCR products were then visualized by UV light and subjected to densitometric analysis by Kodak Image Station program (Eastman Kodak Co., Rochester, NY). The relative expression of target mRNA was normalized to that of β-actin.
Drugs and Reagents. 2,4-Dinitrobenzenesulfonic acid, dexamethasone, hexadecyltrimethylammonium bromide, adenosine deaminase type IX from bovine spleen, and adenosine were purchased from Sigma Aldrich (St. Louis, MO). EHNA was purchased from Tocris (Bristol, UK). Random hexamers, Moloney murine leukemia virus-reverse transcriptase, Taq-polymerase, dithiothreitol, and dNTP were from Promega (Madison, WI). The synthesis of APP was performed as previously reported (Da Settimo et al., 2005). In brief, the commercially available 3-amino-4-pyrazolecarbonitrile was alkylated with 1,2-epoxydecane in dimethylformamide at 100°C in the presence of K2CO3. The resulting compound was then cyclized with boiling formamide to the desired inhibitor APP. APP, EHNA, and dexamethasone were dissolved in sterile dimethyl sulfoxide, and further dilutions were made with sterile saline. The solutions were frozen into aliquots of 2 ml and stored at–80°C until use.
Statistical Analysis. The results are given as mean ± S.E.M. The statistical significance of data was evaluated by one-way analysis of variance followed by post hoc analysis by Student-Newman-Keuls test, and p values lower than 0.05 were considered significant. All statistical procedures were performed using Prism 3.0 software (GraphPad, San Diego, CA).
Results
Adenosine Deaminase Inhibition Assay. EHNA and APP were evaluated for their in vitro ability to inhibit adenosine deaminase. The novel compound APP was found to be highly potent, showing a Ki value of 0.28 ± 0.02 nM, and approximately 4-fold more potent than EHNA (Ki, 1.14 ± 0.10 nM).
Food Intake and Body and Spleen Weight. At day 6 after colitis induction with DNBS, food intake was significantly reduced in comparison with control animals (Fig. 1A). In DNBS-treated rats, APP or EHNA dose-dependently increased food consumption, with maximal effects observed at 15, 45, and 90 μmol/kg, respectively. At these doses, food intake was similar to that of control rats. In inflamed animals receiving dexamethasone (0.25 μmol/kg), food intake did not return toward control levels (Fig. 1A). A significant body weight loss was recorded after colitis induction. Six days after DNBS administration, rats had a mean decrease of–15 ± 5 g in their body weight, whereas control rats displayed a weight gain (+37.5 ± 8.7 g) (Fig. 1B). Significant increments of body weight were observed in inflamed rats under treatment with APP (15 and 45 μmol/kg) or EHNA (30 and 90 μmol/kg), whereas APP (5 μmol/kg), ENHA (10 μmol/kg), or dexamethasone (0.25 μmol/kg) were without effects (Fig. 1B). Measurement of spleen weight was assumed as an index of systemic inflammation (Siegmund et al., 2001). Treatment with DNBS resulted in a significant increment of spleen weight (+30.1%) (Fig. 1C). Such an increase was dose-dependently reduced by administration of APP (+2.5% at 15 μmol/kg), EHNA (+5.6% at 90 μmol/kg), or dexamethasone (+4.1%) (Fig. 1C).
Macroscopic Damage. Six days after DNBS administration, the distal colon appeared thickened and ulcerated with evident areas of transmural inflammation. Adhesions were often present, and the bowel was occasionally dilated, with a macroscopic damage accounting for 7.9 ± 1.3. Rats treated with APP or EHNA displayed a dose-dependent reduction in macroscopic damage score (4.1 ± 0.7 at 15 μmol/kg and 4.2 ± 0.8 at 90 μmol/kg, respectively). Dexamethasone-treated animals showed also a significant decrease in macroscopic damage score (Fig. 2A).

Effects of APP (5, 15, and 45 μmol/kg), EHNA (10, 30, and 90 μmol/kg), or dexamethasone (0.25 μmol/kg) on food intake (A), body weight (B), and spleen weight (C) at day 6 after induction of colitis with DNBS. Each column represents the mean ± S.E.M. (n = 8). *, p < 0.05, significant difference versus control group; a, p < 0.05, significant difference versus DNBS group.

Macroscopic (A) and microscopic (B) damage scores estimated for colon in rats under normal conditions or after DNBS treatment, either alone or in the presence of APP, EHNA, or dexamethasone administration. Each column represents the mean ± S.E.M. (n = 8). *, p < 0.05, significant difference versus control group; a, p < 0.05, significant difference versus DNBS group.
Microscopic Damage. Histological examination of colonic specimens from DNBS-treated rats showed that colitis was well evident and associated with large areas of mucosal necrosis, where the glandular architecture was completely destroyed and only amorphous material could be observed. Submucosa was thickened owing to the presence of edema and marked infiltration with inflammatory cells associated with vasodilation. Sometimes, these alterations involved the muscular layer, which appeared also thickened. The mucosa and submucosa surrounding the necrotic area exhibited inflammation associated with marked cellular infiltration (Fig. 3B), compared with tissues obtained from control animals (Fig. 3A). Colonic sections from inflamed rats displayed a severe damage 6 days after DNBS treatment (Figs. 2B and 3B). In colonic tissues obtained from inflamed rats treated with 15 μmol/kg APP, 90 μmol/kg EHNA, or 0.25 μmol/kg dexamethasone, a significant improvement in microscopic scores was observed (Figs. 2B and 3, C–E).
MPO Assay. DNBS-treated rats showed a marked increase in colonic MPO levels (17.1 ± 4.5 ng/100 mg) compared with control animals (3.2 ± 1.8 ng/100 mg). Treatments with 15 μmol/kg APP, 90 μmol/kg EHNA, or 0.25 μmol/kg dexamethasone significantly prevented the increments of colonic MPO levels associated with DNBS administration (Fig. 4A).
MDA Assay. MDA levels in colonic tissues from control rats were 148 ± 23.8 μmol/mg. In DNBS-treated animals, a marked increase in MDA concentrations was observed (548 ± 33.4 μmol/mg tissue). Treatments with 15 μmol/kg APP, 90 μmol/kg EHNA, or 0.25 μmol/kg dexamethasone significantly attenuated the increase in tissue MDA levels elicited by colitis induction with DNBS, although being still significantly higher than control (Fig. 4B).

Hematoxylin and eosin-stained sections of rat colon. Microscopic images refer to control rats (A) or DNBS-treated animals, either alone (B) or in the presence of APP (15 μmol/kg) (C), EHNA (90 μmol/kg) (D), or dexamethasone (0.25 μmol/kg) (E).
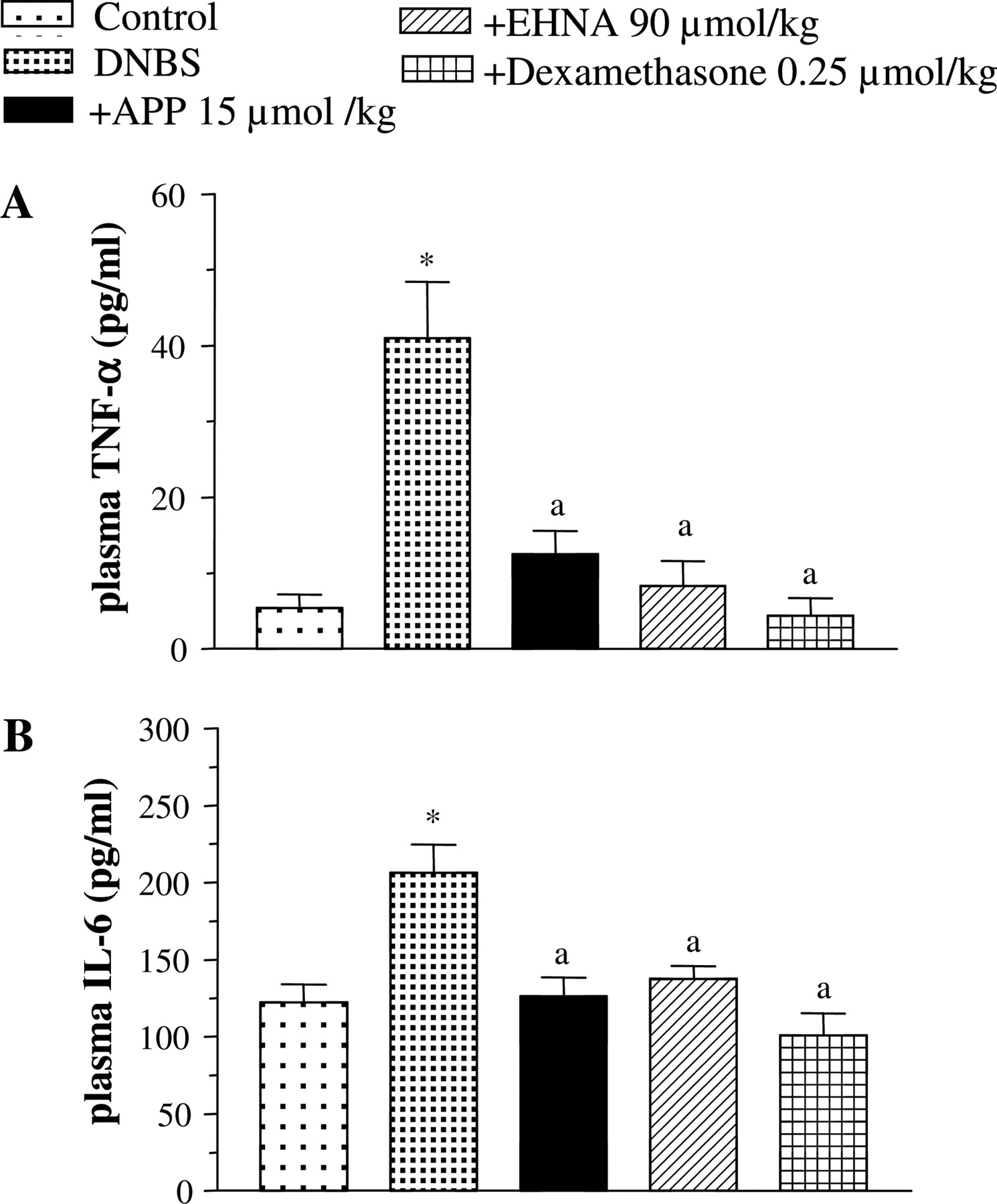
TNF-α and IL-6 Assays. Colonic injury induced by DNBS was associated with a significant increase in tissue TNF-α levels (from 3.7 ± 1.0 to 8.9 ± 1.5 pg/mg). Treatments with all test drugs significantly decreased the concentration of this cytokine in colonic tissues (Fig. 4C). In rats with colitis, plasma TNF-α levels were markedly increased in comparison with control animals (from 5.4 ± 1.8 to 41 ± 7.4 pg/ml) (Fig. 5A). This increment was completely prevented by treatments with 15 μmol/kg APP, 90 μmol/kg EHNA, or 0.25 μmol/kg dexamethasone (Fig. 5A). The induction of colitis was also associated with a significant increment of plasma IL-6 (from 122.7 ± 11.5 to 206.5 ± 18.0 pg/ml). IL-6 plasma levels were normalized upon administration of APP, EHNA, or dexamethasone (Fig. 5B).
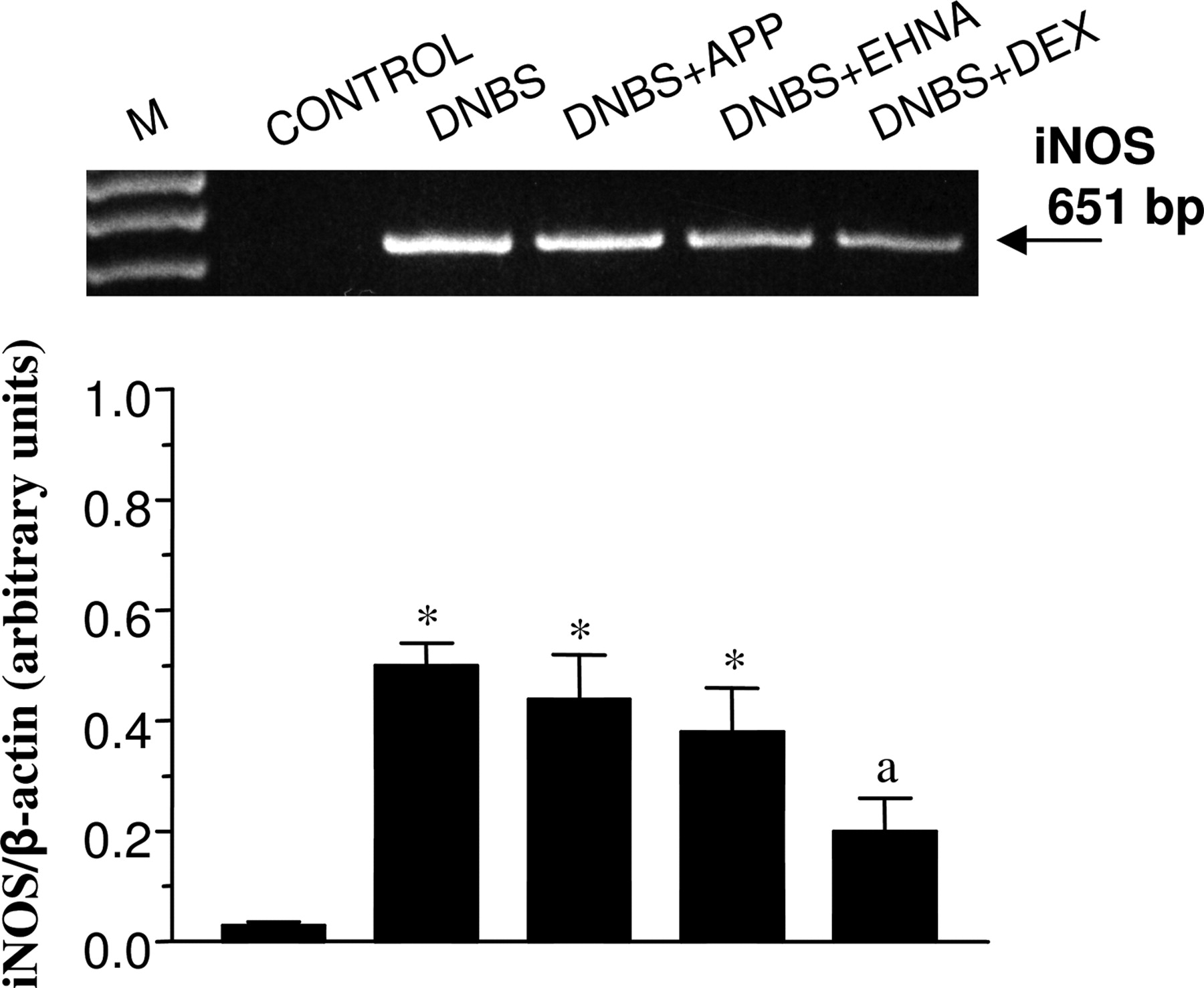
iNOS and COX-2 mRNA Expression. RT-PCR analysis showed that the expression of iNOS mRNA was significantly enhanced in colonic mucosa from DNBS-treated rats in comparison with control animals. In rats with colitis, treatments with 15 μmol/kg APP or 90 μmol/kg EHNA were without effects, whereas 0.25 μmol/kg dexamethasone significantly reduced the induced iNOS expression (Fig. 6). The expression of COX-2 mRNA was detected in the colon of control rats as well as in colonic tissues of animals treated with DNBS alone or in combination with EHNA, APP, or dexamethasone. The densitometric analysis, performed on amplified cDNA bands, indicated no appreciable differences in the expression of COX-2 mRNA among the groups examined (not shown).
Discussion
In the present study, evidence was obtained that the pharmacological modulation of adenosine deaminase, a key enzyme involved in the metabolic fade of endogenous adenosine (Haskó and Cronstein, 2004), attenuates bowel inflammation in an experimental model of colitis and that the novel adenosine deaminase inhibitor APP is more effective than its comparator EHNA in controlling several parameters related to intestinal inflammation.
The DNBS-induced colitis is a model of bowel inflammation characterized by body weight loss, diarrhea, ulceration and bleeding, depletion of goblet cells, and formation of granulomas within the gut wall (Jurjus et al., 2004). In this model, oxidative damage is also known to play a significant role in the development of tissue injury occurring within a few days from DNBS administration (Elson et al., 1995). Animals with DNBS-induced colitis can be employed in pre-clinical investigations to assay the anti-inflammatory properties of novel drugs with potential usefulness against intestinal inflammation in humans (Elson et al., 1995; Wirtz and Neurath, 2000). In our experiments, the suitability of this model was demonstrated by the ability of dexamethasone to attenuate both systemic and tissue phlogistic parameters, and these findings were in keeping with previous reports showing the efficacy of this potent anti-inflammatory drug in rats with colitis (Nakase et al., 2001).
In the present study, both APP and EHNA increased food intake and body weight and decreased spleen weight and plasma TNF-α and IL-6 levels, regarded as systemic indexes of inflammation. Moreover, treatments with both adenosine deaminase inhibitors were associated with improvements in macroscopic and histological features of colitis, thus extending to the digestive tract the concept that preserving endogenous adenosine from metabolic inactivation can result in a significant interference with phlogistic processes (Siegmund et al., 2001; Kuno et al., 2006). In line with this view, drugs able to increase tissue concentrations of adenosine via block-ade of adenosine kinase have been shown to be also effective in experimental bowel inflammation (Siegmund et al., 2001). The anti-inflammatory actions of adenosine could depend on the activation of A2a and A3 receptors because the protective effects exerted by the A2a agonist ATL313 against intestinal inflammation were reverted by ZM241385, an A2a antagonist (Cavalcante et al., 2006), and the stimulation of A3 receptors can afford protection in experimental models of colitis (Mabley et al., 2003a). It is noteworthy that inosine, the metabolite produced by adenosine deaminase, is also effective in reducing the severity of bowel inflammation (Mabley et al., 2003b) and protecting against endotoxin-induced shock (Haskó et al., 2000). Therefore, the blockade of adenosine deaminase could hamper the anti-inflammatory potential resulting from inosine production, but at the same time this deficiency seems to be overcome by the concomitant adenosine accumulation that ensures a significant and more effective anti-inflammatory activity, as shown in the present and previous studies (Siegmund et al., 2001).

MPO (A), MDA (B), and TNF-α levels (C) in colonic tissues from control rats or in animals treated with DNBS, either alone or in combination with APP (15 μmol/kg), EHNA (90 μmol/kg), or dexamethasone (0.25 μmol/kg). Each column represents the mean ± S.E.M. (n = 6–8). *, p < 0.05, significant difference versus control rats; a, p < 0.05, significant difference versus DNBS group.

TNF-α (A) and IL-6 (B) levels in plasma from control or DNBS-treated rats, either alone or in the presence of APP (15 μmol/kg), EHNA (90 μmol/kg), or dexamethasone (0.25 μmol/kg). Each column represents the mean ± S.E.M. (n = 8). *, p < 0.05, significant difference versus control rats; a, p < 0.05, significant difference versus DNBS group.

RT-PCR analysis of iNOS mRNA expression in colonic tissues from control animals or rats treated with DNBS, either alone or in combination with APP (15 μmol/kg), EHNA (90 μmol/kg), or dexamethasone (0.25 μmol/kg). Each column represents the mean ± S.E.M. (n = 5). *, p < 0.05, significant difference versus control rats; a, p < 0.05, significant difference versus DNBS alone. M, size markers; DEX, dexamethasone.
Several inflammatory mediators contribute to the pathogenesis of bowel inflammation (Ardizzone and Bianchi Porro, 2005). In particular, TNF-α plays a pivotal role both in human and experimental colitis, inducing the production of chemoattractants for neutrophils and promoting their activation (Martin and Wallace, 2006). Once activated, neutrophils pass out of circulation, infiltrate the mucosa and sub-mucosa, and contribute to intestinal injury through a variety of mechanisms, including the generation of reactive oxidant species (Oz et al., 2005). Our findings show that TNF-α and MPO levels were reduced in the colonic tissues of animals treated with APP or EHNA. The ability of these compounds to decrease TNF-α levels and neutrophil infiltration by prevention of adenosine degradation is consistent with previous reports showing that high tissue levels of adenosine inhibit the synthesis of inflammatory cytokines induced by phlogistic stimuli and promote self-limiting immune responses through down-regulation of neutrophil recruitment and activation (Haskó and Cronstein, 2004; Kuno et al., 2006). We could also observe a similar profile of cytokine modulation in plasma, where, together with the reduction of TNF-α, APP- and EHNA-treated animals displayed a decrease in IL-6, a cytokine involved in the maintenance of inflammatory activity via activation and differentiation of immune cells (Scheller et al., 2006). It is noteworthy, however, that the present experiments were designed to assay the ability of pretreatment with adenosine deaminase inhibitors to protect bowel tissues from application of inflammatory stimuli, whereas the efficacy of these compounds against established colitis was not evaluated. Nevertheless, it is likely that inhibitors of adenosine deaminase would function also in this setting since previous studies have shown that adenosine receptor agonists are effective against established forms of bowel inflammation (Odashima et al., 2005).
As anticipated above, there is evidence that oxidant-mediated tissue damage plays an important role in the pathophysiology of bowel inflammation. A number of reports have demonstrated an increased generation of reactive oxygen species that exceeds the capacity of intestinal antioxidant defense and contributes to tissue injury in the setting of bowel inflammation (Oz et al., 2005; Martin and Wallace, 2006). In our experiments, a significant increase in lipid peroxidation index was observed in the inflamed colon of DNBS-treated rats, and this parameter was substantially reduced by repeated administrations of APP or EHNA. These findings are in accordance with those of Bor et al. (1999), who demonstrated that the adenosine deaminase inhibitor pentostatin reduced MDA production in the kidney of rats subjected to ischemia-reperfusion damage, and suggest that blunting of oxidant processes contributes to the mechanisms through which adenosine accumulation provides protection against inflammatory damage. Overall, it is conceivable that block-ade of adenosine deaminase can counteract tissue oxidative injury through inhibition of TNF-α release in the inflamed colonic tissue and subsequent reduction of neutrophil activation.
Several studies, based on animal models or conducted in humans, have indicated that iNOS is involved in intestinal inflammation and exerts a pathogenetic role in IBD, supporting the phlogistic process by production of nitric oxide that can be converted into reactive nitrogen species via interaction with reactive oxygen species (Kolios et al., 2004). Consistently with this notion, we observed iNOS induction in the colon of inflamed rats. Furthermore, the increase in iNOS expression was counteracted by dexamethasone, as previously reported by Asfaha et al. (1999). However, the enhanced iNOS expression was unaffected by treatments with APP or EHNA, suggesting that the anti-inflammatory effects of these adenosine deaminase inhibitors are not likely to depend on modulation of iNOS expression. The significance of cyclooxygenase pathways in maintaining mucosal homeostasis and/or promoting gut inflammation is highly debated (Wallace and Devchand, 2005; Martin and Wallace, 2006). Under the present experimental conditions, colitis was not associated with significant variations of COX-2 expression, and this pattern was not modified by APP, EHNA, or dexamethasone. These findings are not fully consistent with other reports showing increments of COX-2 expression in inflamed bowel tissues (Khan et al., 2002). However, this apparent discrepancy can be explained considering that COX-2 expression varies throughout the time course of enteric inflammation. For instance, Kankuri et al. (2001) demonstrated that COX-2 colonic expression in rats with colitis reaches its maximal level after 12 to 24 h from application of inflammatory stimulus and returns to basal values after 72 h. Moreover, in line with our results, Kankuri et al. (2001) observed that dexamethasone did not affect COX-2 expression in rats with colitis. Therefore, it is conceivable that the amelioration of bowel inflammation in rats treated with adenosine deaminase inhibitors occurs independently of changes in COX-2 expression.
One major purpose of this study was to evaluate the pharmacodynamic properties of APP as a novel adenosine deaminase inhibitor. To pursue this goal, we first assayed APP in vitro to estimate its potency in inhibiting adenosine deaminase activity and then in vivo to test its effects in the model of DNBS-induced colitis. Our data show that APP counteracted intestinal inflammation with greater potency than EHNA and that the difference in the anti-inflammatory effects of these compounds reflects their respective potencies in inhibiting the activity of adenosine deaminase in vitro. Moreover, APP displays a favorable profile of selectivity for adenosine deaminase because it binds adenosine receptors with extremely low potency (Da Settimo et al., 2005); therefore, it may represent a lead compound for the identification of novel adenosine deaminase inhibitors, characterized by high degrees of potency and selectivity.
The limitations in both efficacy and safety, encountered with drugs available for treatment of bowel inflammation, continue to foster the search for better therapeutic agents. Recent insights into the pathogenesis of IBD have led to identification of novel treatment options (Hanauer and Present, 2003). Although traditional treatments have focused on nonspecific suppression of inflammation, advances in the knowledge of the immunopathogenesis of intestinal inflammation have paved the way to targeted therapies, allowing a selective blockade of the inflammatory cascade and modulation of key cytokines. In this framework, the adenosine system represents an attractive target for novel therapies against inflammatory diseases. According to the present results, the blockade of adenosine deaminase allows an effective control of experimental intestinal inflammation, and the novel adenosine deaminase inhibitor APP can provide a basis for development of anti-inflammatory drugs suitable for treatment of inflammatory intestinal disorders.
Acknowledgments
We acknowledge Piero Duranti (Department of Internal Medicine, University of Pisa) for valuable technical support.
Footnotes
-
Article, publication date, and citation information can be found at http://jpet.aspetjournals.org.
-
doi:10.1124/jpet.107.122762.
-
ABBREVIATIONS: IBD, inflammatory bowel disease; TNF, tumor necrosis factor; IL, interleukin; APP, 4-amino-2-(2-hydroxy-1-decyl)pyrazole[3,4-d]pyrimidine; EHNA, erythro-9-(2-hydroxy-3-nonyl)adenine hydrochloride; DNBS, 2,4-dinitrobenzenesulfonic acid; MPO, myeloperoxidase; MDA, malondialdehyde; iNOS, inducible nitric-oxide synthase; COX, cyclooxygenase; RT, reverse transcription; PCR, polymerase chain reaction; dNTP, deoxynucleotide triphosphate.
- Received March 15, 2007.
- Accepted May 7, 2007.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}