


Abstract
Pruritus (itch sensation) is a significant clinical problem. The aim of this study was to elucidate the roles of opioid receptor types and the site of action in opioid-induced itch in monkeys. Observers who were blinded to the conditions counted scratching after administration of various drugs. Intravenous (i.v.) administration of μ opioid receptor (MOR) agonists (fentanyl, alfentanil, remifentanil, and morphine) evoked scratching in a dose- and time-dependent manner. However, the κ opioid agonist U-50488H [trans-(±)-3,4-dichloro-N-methyl-N-(2-[1-pyrrolidinyl]-cyclohexyl)-benzeneacetamide] and δ opioid agonist SNC80 [(+)-4-[(αR)-α-[2S,5R)-4-allyl-2,5-dimethyl-1-piperazinyl]-3-methoxybenzyl]-N,N-diethylbenzamide] did not increase scratching. Intrathecal (i.t.) administration of peptidic MOR agonist [d-Ala2,N-Me-Phe4,Gly5-ol]-enkephalin (DAMGO, 0.00032–0.01 mg) evoked scratching, but i.v. DAMGO (0.01–1 mg/kg) did not increase scratching. A similar difference between i.t. and i.v. effectiveness was seen with morphine. Antagonist studies revealed that i.v. administration of an opioid receptor antagonist (naltrexone, 0.0032–0.1 mg/kg) dose dependently attenuated scratching induced by i.v. fentanyl (0.018 mg/kg) or morphine (1 mg/kg). However, a peripherally selective opioid antagonist (quaternary naltrexone, 0.0032–0.32 mg/kg) did not block i.v. fentanyl- or morphine-induced scratching. Moreover, a histamine antagonist (diphenhydramine, 0.1–10 mg/kg), failed to attenuate scratching induced by i.t. morphine (0.032 mg) or i.v. morphine (1 mg/kg). Pretreatment with a selective MOR antagonist (clocinnamox, 0.1 mg/kg), but not κ or δ opioid antagonists (nor-binaltorphimine or naltrindole), blocked i.t. morphine-induced scratching. Together, these data suggest that MOR, not other opioid receptor types or histamine, mediates scratching evoked by opioid analgesics. More important, this study provides in vivo pharmacological evidence that activation of central MOR plays an important role in opioid-induced itch in primates.
Pruritus (itch sensation) is a significant clinical problem (Twycross et al., 2003), but little effort has been made to understand mechanisms by which itch sensation is provoked. Clinical studies have reported pruritus after spinal or systemic administration of various opioid analgesics (Ellis et al., 1990; Palmer et al., 1999; Szarvas et al., 2003). Spinal opioid-induced itch is sometimes severe, and it may lessen the value of spinal opioids for pain relief (Szarvas et al., 2003). In addition, opioid receptor antagonists have been proposed to treat pruritus caused by cholestasis, indicating the involvement of endogenous opioids in itch (Wolfhagen et al., 1997; Jones et al., 2002). Nevertheless, few basic mechanism studies have thoroughly elucidated the role of opioid receptor types and sites of actions in pruritus.
Compared with the literature on pain research, there is a large deficiency in the basic research on pruritus. This could be in part due to lack of reliable animal models. For example, histamine has been widely used in humans to evoke itch sensation (Simone et al., 1987; Schmelz et al., 1997). However, histamine does not evoke itch (i.e., scratching behavior) in rodents (Kuraishi et al., 1995; Jinks and Carstens, 2002). In addition, spinal administration of morphine produces intense long-lasting itch sensation/responses in both humans and monkeys (Bailey et al., 1993; Palmer et al., 1999; Ko and Naughton, 2000). However, spinal administration of morphine in rodents does not evoke long-lasting scratching responses (Kuraishi et al., 2000; Lee et al., 2003). Therefore, it should be noted that species differences may contribute to different results or interpretations in the in vivo pharmacology of itch.
Previously, we have established an experimental model of itch in monkeys (Ko and Naughton, 2000). Intrathecal (i.t.) administration of morphine dose dependently produces both scratching responses and antinociception in monkeys and this observation parallels closely with the behavioral effects of morphine in human studies (Bailey et al., 1993; Palmer et al., 1999). This experimental model of itch provides a valuable tool to elucidate several issues regarding opioid-induced itch in primates. For instance, systemic administration of κ opioid receptor (KOR) or δ opioid receptor (DOR) agonists may represent potential analgesics (Butelman et al., 1993; Negus et al., 1998), but the roles of KOR and DOR in itch have not been extensively verified. The medullary dorsal horn has been identified as one of central sites of action in morphine-evoked facial scratching in monkeys (Thomas et al., 1993a). However, it is not clear whether itch associated with systemic administration of various opioid analgesics is mediated exclusively by central μ opioid receptors (MORs). It is important to investigate whether the sites of pruritic action of MOR agonists are mainly located in the central nervous system. Such studies would clarify the role of central MOR in itch and facilitate the development of antipruritics targeting central MOR.
Although morphine can release histamine from mast cells, clinical studies have suggested that antihistamines may not be effective in relieving morphine-induced itch (Dunteman et al., 1996; Krajnik and Zylicz, 2001). The roles of MOR and histamine in opioid-induced itch can be further clarified by comparing the effectiveness between opioid receptor antagonists and antihistamines in reducing morphine-evoked scratching in monkeys. In addition, other MOR agonists, such as fentanyl, do not induce histamine release (Rosow et al., 1982; Hermens et al., 1985). It will be important to study whether the durations of scratching evoked by fentanyl-like analogs are correlated with their durations of MOR-mediated antinociception and respiratory depression (Ko et al., 2002). Furthermore, an opioid's site of action can be investigated by using compounds with different lipophilicities after different administration routes (Ko et al., 1998b). If the site of pruritic action is mainly central, it is expected that hydrophilic opioids would have a large potency difference between i.t. and systemic administration. The aim of this study was therefore to investigate opioid-induced itch in primates by using pharmacological approaches. In particular, the present study examined the roles of central versus peripheral site and opioid receptor types in itch.
Materials and Methods
Subjects
Twenty adult intact male and female rhesus monkeys, Macaca mulatta, with body weights ranging between 6.7 and 14.0 kg were used. They were housed individually with free access to water and were fed approximately 25 to 30 biscuits (Purina Monkey Chow; Ralston Purina, St. Louis, MO) and fresh fruit daily. No monkey had exposure to opioids for 1 month before the present study. The monkeys were housed in facilities accredited by the American Association for the Accreditation of Laboratory Animal Care. The studies were conducted in accordance with the University Committee on the Use and Care of Animals in the University of Michigan and the Guide for the Care and Use of Laboratory Animals (National Academy Press, Washington DC, revised 1996).
Procedures
Scratching Responses. Scratching responses, inferred as an itch sensation (Ko and Naughton, 2000), were recorded on videotapes when monkeys were in their home cages. Each recording session was conducted for 15 min/test session. A scratch was defined as one short-duration (<1 s) episode of scraping contact of the forepaw or hind paw on the skin surface of other body parts. Scratches usually occurred repetitively at the same location. Scratching responses were scored by trained individuals who were blinded to experimental conditions.
Intrathecal Injection. Monkeys were positioned in primate restraint chairs and anesthetized by i.v. propofol (2.5–4.0 mg/kg for bolus infusion and 0.3–0.4 mg/kg/min for continuous infusion; AstraZeneca Pharmaceuticals LP, Wilmington, DE). The lower back of the trunk was shaved and prepared sterilely with Betadine. A spinal needle (22-gauge × 1.5; BD Biosciences, Franklin Lakes, NJ) was inserted into the subarachnoid space between L4/L5 lumbar vertebra. Needle position was confirmed by a free flow of clear cerebrospinal fluid. A 1-ml solution of compound was infused slowly through the spinal needle within 30 s. Monkeys recovered from anesthesia within 10 min after termination of propofol infusion and were returned to their home cages.
Intravenous Injection. Monkeys who were involved in the first part of the study had been adapted to wearing a Teflon mesh jacket attached to a flexible steel tether (Lomir Biomedical, Malone, NY). Aseptic surgical techniques were used to implant a silicone catheter into one of eight major veins (external or internal jugular, femoral, or brachial viens). The proximal end of the catheter was inserted close to the heart, and the distal end was run subcutaneously from the incision site toward the monkey's back, existing at the midscapular region. The catheter traveled through the tether to the back of cage where it connected to a swivel. This apparatus allows immediate videorecording of scratching after i.v. administration of test compounds without moving monkeys back and forth to their cages. However, this apparatus does not allow performing i.t. injection in monkeys in primate restraint chairs. Monkeys who were involved in the second and third parts of the study (i.e., i.t. administration) received i.v. administration of drugs through a saphenous vein and then they were returned to their home cages.
Experimental Designs
Effects of i.v. Opioid Receptor Agonists. The first part of the study was to investigate effects of selective MOR, KOR, and DOR agonists in evoking scratching responses after i.v. administration. Four male and four female monkeys were used in all experiments, which were conducted once per week. Selective MOR agonists (fentanyl, 0.0032–0.018 mg/kg; alfentanil, 0.0032–0.018 mg/kg; remifentanil, 0.001–0.0056 mg/kg), a KOR agonist (U-50488H, 0.01–0.32 mg/kg), and a DOR agonist (SNC80, 0.01–0.32 mg/kg) were administered intravenously using a single-dosing procedure. Doses of these opioid receptor agonists were chosen based on previous studies in monkeys showing behavioral effects after systemic administration (Negus et al., 1998; Ko et al., 1998a, 2002). Higher doses of i.v. U-50488H and SNC80 (i.e., ≥1 mg/kg) were not tested to avoid sedation and convulsions. Recording of scratching activity was performed in 15-min bins for 1 h immediately after injection. As noted, i.v. fentanyl 0.018 mg/kg produced maximum scratching responses and a higher dose of fentanyl could not be tested due to respiratory arrest. Additional investigations with antagonists were conducted to compare the effectiveness of two opioid receptor antagonists in attenuating i.v. fentanyl-induced scratching. A single dose of naltrexone (NTX, 0.0032–0.1 mg/kg) or quaternary-naltrexone (Q-NTX, 0.0032–0.1 mg/kg) was administered intravenously 15 min before i.v. injection of fentanyl 0.018 mg/kg.
Comparison of MOR Agonist-Induced Scratching. The second part of the study was to determine the potency of morphine and DAMGO, a peptidic MOR-selective agonist, after either i.t. or i.v. administration. Three male and three female monkeys were used in all experiments, which were conducted with a 10-day interinjection interval. The effects of morphine (i.t., 0.0032–0.032 mg; i.v., 0.1–3.2 mg/kg) and DAMGO (i.t., 0.00032–0.01 mg; i.v., 0.01–1 mg/kg) were studied by giving different doses randomly in a single dosing procedure. Doses of both morphine and DAMGO were chosen based on previous studies in monkeys that have shown behavioral effects after injection (Ko et al., 1998b, 2003a). Recording of scratching was performed during the 24 to 39th min of each hour (i.e., 15 min/session) for 6 h after either i.t. or i.v. injection.
Effects of Opioid and Histamine Antagonists on Morphine-Induced Scratching. The third part of the study was to investigate the effects of opioid receptor antagonists and a histamine antagonist (diphenhydramine) on morphine-induced scratching. Three male and three female monkeys were used in all experiments. The first experiment was conducted with a 10-day interinjection interval. NTX (0.0032–0.032 mg/kg) or diphenhydramine (0.1–10 mg/kg) was administered intramuscularly at 45 min after i.t. morphine to determine whether either compound could reverse scratching. Morphine 0.032 mg was chosen because i.t. morphine 0.032 mg produced maximum scratching responses (Ko and Naughton, 2000; Ko et al., 2003a). In addition, doses of diphenhydramine were chosen based on monkey studies showing that they are systemically active doses (Rowlett et al., 1999). Recording of scratching was conducted during 24 to 39th min and 54 to 09th min of each hour (i.e., 15 min/session) for 3 h after i.t. injection of morphine. Moreover, pretreatment doses of NTX (0.032 mg/kg), Q-NTX (0.032–0.32 mg/kg), or diphenhydramine (0.32–3.2 mg/kg) were administered to examine whether any compound could prevent i.v. morphine-induced scratching. Morphine 1 mg/kg was chosen because this dose produced maximum scratching responses after systemic administration in rhesus monkeys (Ko et al., 2003a; the present study). Because the durations of action of these pretreatment compounds vary, and they are shorter than the duration of morphine-induced scratching, only one session of recording was conducted. Recording of scratching was performed during 24 to 39th min after i.v. morphine injection. NTX, Q-NTX, or diphenhydramine was administered intramuscularly at 15 min before i.v. morphine.
The second experiment was conducted to examine the effects of MOR-, KOR-, and DOR-selective antagonists on i.t. morphine-induced scratching. A MOR antagonist (clocinnamox, 0.1 mg/kg) or a KOR antagonist (nor-binaltorphimine, 3.2 mg/kg) was given intramuscularly 24 h before i.t. morphine (0.032 mg). A DOR antagonist (naltrindole, 1 mg/kg) was given intramuscularly 30 min before i.t. morphine. The pretreatment time and dose of each antagonist were chosen based on previous studies showing that selective receptor antagonism occurs after systemic administration in monkeys (Butelman et al., 1993; Zernig et al., 1994; Negus et al., 1998). The interinjection interval between antagonists was 3 weeks and norbinaltorphimine was the last antagonist to be tested due to its long-acting KOR-antagonism (Butelman et al., 1993). Recording of scratching was conducted during 24 to 39th min and 54 to 09th min of each hour (i.e., 15 min/session) for 3 h after i.t. morphine.
Data Analysis
Mean values (mean ± S.E.M.) were calculated from individual values for all behavioral endpoints. Comparisons were made for the same monkeys across all test sessions in the same experiment. As noted, we did not find a significant difference in the effects of agonists or antagonists between male and female monkeys, so mean values for all monkeys in the same condition were used for data analysis. All data were analyzed by either one-way or two-way analysis of variance followed by the Newman-Keuls test for multiple (post hoc) comparisons. The criterion for significance was set at p < 0.05.
Drugs
Fentanyl HCl, alfentanil HCl, DAMGO trifluoroacetate, NTX HCl, naltrindole HCl (National Institute on Drug Abuse, Bethesda, MD), remifentanil HCl (GlaxoSmithKline, Research Triangle Park, NC), U-50488H methanesulfonate, diphenhydramine HCl, nor-binaltorphimine 2HCl (Sigma-Aldrich, St. Louis, MO), Q-NTX HCl (Dr. H. I. Mosberg, University of Michigan, Ann Arbor, MI), and morphine sulfate (Mallinckrodt, St. Louis, MO) were dissolved in sterile water. SNC80 free base (Dr. K. C. Rice, National Institute of Diabetes and Digestive and Kidney Diseases, Bethesda, MD) and clocinnamox mesylate (Dr. J. W. Lewis, University of Bath, Bath, UK) were dissolved in 3% lactic acid in sterile water. For i.m. administration, all compounds were administered intramuscularly into the thigh muscle at a volume of 0.1 ml/kg. Doses are presented in the compound forms listed above.
Results
Effects of i.v. Opioid Receptor Agonists.Fig. 1 illustrates the magnitude and duration of i.v. MOR agonist-induced scratching responses. Fentanyl increased scratching in both dose-[F(3,21) = 89.2; p < 0.05] and time-dependent [F(3,21) = 3.6; p < 0.05] manners. Scratching evoked by i.v. fentanyl 0.018 mg/kg peaked at the first observation period and continued throughout the 1-h observation period (see Table 1 for other details). Alfentanil also increased scratching in both dose-[F(3,21) = 31.4; p < 0.05] and time-dependent [F(3,21) = 30.9; p < 0.05] manners. Alfentanil was equipotent with fentanyl, also producing the maximum response at a dose of 0.018 mg/kg. Alfentanil-induced scratching had returned to control levels by approximately 30 to 45 min after i.v. administration of 0.018 mg/kg. Remifentanil was also effective in provoking scratching dose [F(3,21) = 12.0; p < 0.05] and time dependently [F(3,21) = 171.8; p < 0.05]. Scratching evoked by i.v. remifentanil 0.0056 mg/kg peaked at the first observation period and declined by approximately 15 to 30 min after administration. The durations of remifentanil-induced scratching were shorter than those of fentanyl and alfentanil (Table 1). In contrast, both U-50488H (0.01–0.32 mg/kg) and SNC80 (0.01–0.32 mg/kg) did not significantly evoke scratching responses compared with the vehicle condition during the entire observation period. Table 1 summarizes the duration and significance of scratching evoked by above-mentioned opioid receptor agonists based on post hoc comparisons.

Time course of MOR agonist-induced scratching responses. Abscissae, time in minutes after i.v. administration of each agonist (milligrams per kilogram). Ordinates, scratches per 15 min. Each value represents mean ± S.E.M. (n = 8). Symbols represent different experimental conditions for the same monkeys. See Table 1 and Materials and Methods for other details.
Comparison of scratching responses evoked by selective MOR, KOR, and DOR agonists in monkeys All compounds (milligrams per kilogram) were administered intravenously. See Fig. 1 for other details.
Figure 2 compares the antagonist effects of both NTX and Q-NTX on i.v. fentanyl-induced scratching. Pretreatment with i.v. NTX dose dependently attenuated i.v. fentanyl (0.018 mg/kg)-induced scratching [F(4,35] = 63.9; p < 0.05]. Post hoc comparisons indicated that NTX at the dose range of 0.01 to 0.1 mg/kg significantly attenuated fentanyl-induced scratching throughout the entire test sessions. In contrast, pretreatment with i.v. Q-NTX at the same dose range did not significantly block fentanyl-induced scratching. These doses of NTX or Q-NTX alone did not produce any behavioral effect (e.g., sedation) in monkeys.

Effects of NTX or Q-NTX on fentanyl-induced scratching. Abscissae, dose of antagonist (milligrams per kilogram). Ordinates, total scratching responses in 60 min after i.v. administration of fentanyl (0.018 mg/kg). Each value represents mean ± S.E.M. (n = 8). Either NTX or Q-NTX was given intravenously 15 min before i.v. fentanyl. The open circle represents the effect of i.v. fentanyl alone without pretreatment with NTX or Q-NTX (i.e., the vehicle condition). The asterisk represents a significant difference from the vehicle condition (*, p < 0.05). See Fig. 1 and Materials and Methods for other details.
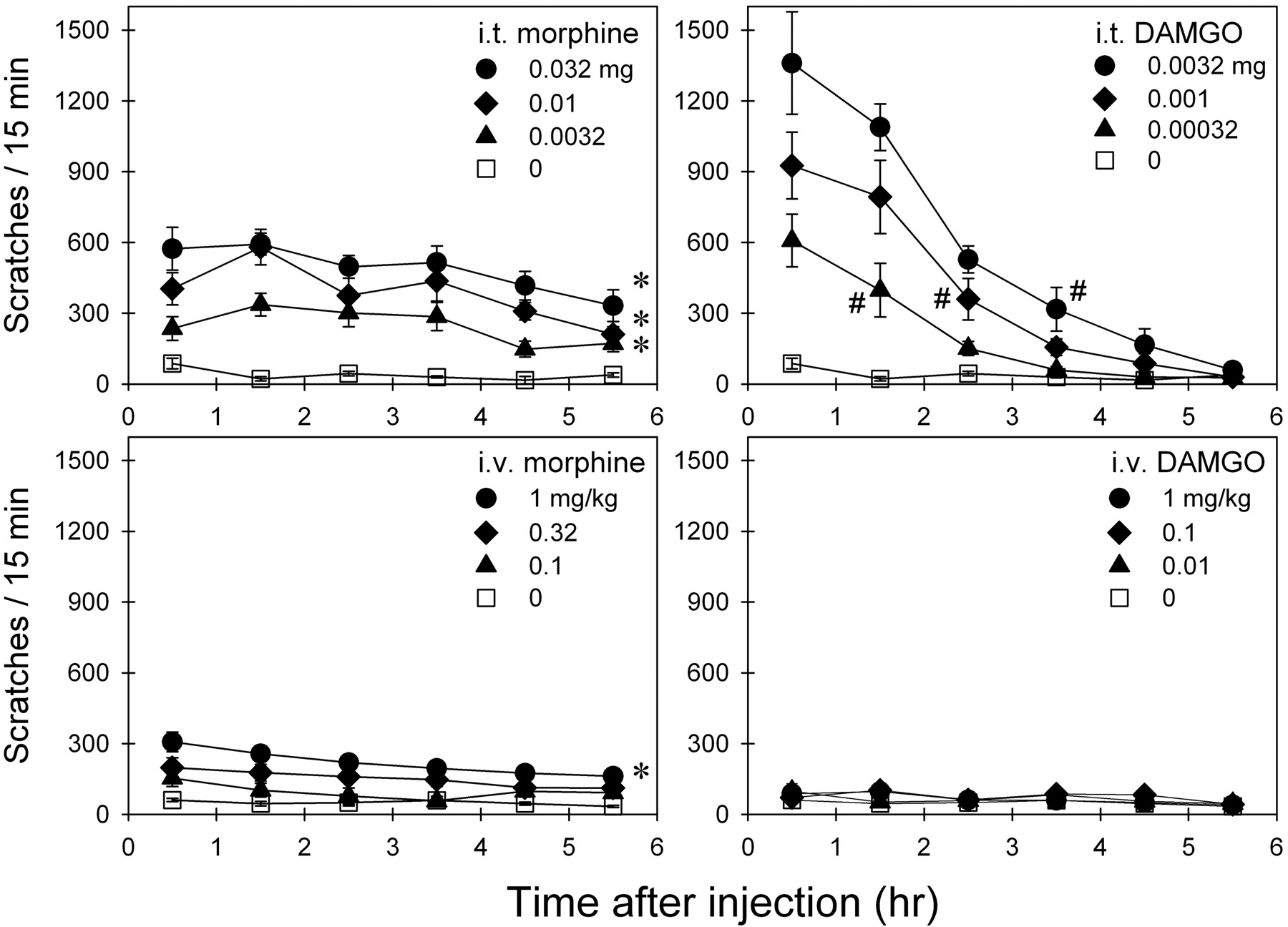
Comparison of MOR Agonist-Induced Scratching.Fig. 3 illustrates the magnitude and duration of scratching responses evoked by i.t. and i.v. administration of morphine or DAMGO. Intrathecal administration of morphine increased scratching in both dose-[F(3,15) = 56.2; p < 0.05] and time-dependent [F(5,25) = 17.5; p < 0.05] manners. Post hoc comparisons indicated that scratching evoked by all three doses of morphine (0.0032–0.032 mg) significantly lasted for 6 h compared with the vehicle condition (p < 0.05). In addition, i.v. administration of morphine increased scratching dose [F(5,25) = 18.6; p < 0.05] and time dependently [F(5,25) = 7.0; p < 0.05]. Post hoc comparisons indicated that scratching evoked by two doses of morphine (i.e., 1 and 1.8 mg/kg) significantly lasted for 6 h (p < 0.05).

Time course of scratching responses evoked by i.t. and i.v. administration of morphine or DAMGO. Abscissae, time in hours after administration of each agonist. Ordinates, scratches per 15 min. Each value represents mean ± S.E.M. (n = 6). Symbols represent different experimental conditions for the same monkeys. The asterisk represents a significant difference from the vehicle condition between time points 0.5 and 5.5 h (*, p < 0.05). The # represents a significant difference from the vehicle condition from time points 0.5 h to the corresponding time point for each dose (p < 0.05). Not all of the data are shown for the sake of clarity. See Fig. 4 for the complete dose-response of i.v. morphine-induced scratching.
In the same group of monkeys, i.t. administration of DAMGO also increased scratching in both dose-[F(4,20) = 38.2; p < 0.05] and time-dependent [F(5,25) = 83.4; p < 0.05] manners. Post hoc comparisons indicated that both doses of DAMGO (i.e., 0.0032–0.01 mg) produced similar scratching responses that peaked in the first hour and declined within 3 to 4 h after i.t. injection. In particular, the maximum scratching responses evoked by DAMGO (0.0032–0.01 mg) were larger than those evoked by i.t. morphine 0.032 mg in the first 2 h after administration (p < 0.05). It is worth noting that 15 min pretreatment with i.m. NTX 0.032 mg/kg significantly blocked i.t. DAMGO (0.0032 mg)-induced scratching (data not shown). In contrast, i.v. administration of DAMGO (i.e., 0.01–1 mg/kg) did not significantly evoke scratching responses compared with the vehicle condition during the entire observation period.
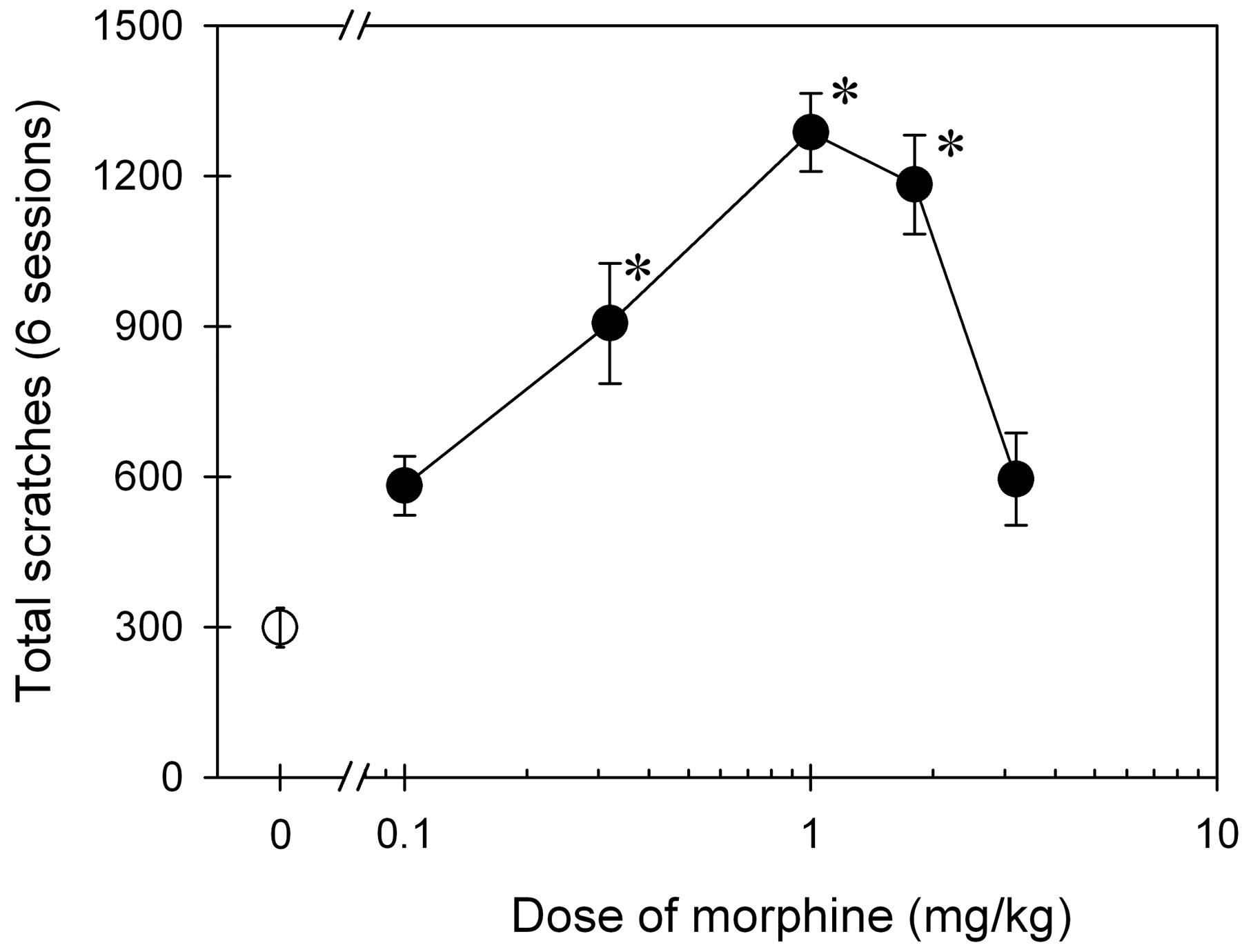
Figure 4 illustrates a complete dose-response curve of i.v. morphine for scratching responses sampled by a 15 min bin per hour for the entire 6-h observation period (Note: part of the data were directly converted from Fig. 3). There are significant differences in total scratching evoked by i.v. morphine and vehicle [F(8,45) = 22.7; p < 0.05]. Post hoc comparisons indicated that i.v. morphine at the dose range of 0.32 to 1.8 mg/kg significantly increased scratching compared with the vehicle condition (p < 0.05).

Dose-response curve of i.v. morphine for scratching in monkeys. Abscissae, dose of morphine given intravenously (milligrams per kilogram). Ordinates, total scratching responses (i.e., accumulated scratches in a 15-min bin/h for 6 h). Each value represents mean ± S.E.M. (n = 6). Symbols represent different doses of morphine. The asterisk represents a significant difference (*, p < 0.05) from the vehicle condition (open circle). See Fig. 3 and Materials and Methods for other details.
Effects of Opioid and Histamine Antagonists on Morphine-Induced Scratching.Fig. 5 illustrates the effects of NTX and diphenhydramine on i.t. morphine-induced scratching. Intramuscular NTX dose dependently attenuated i.t. morphine (0.032 mg)-induced scratching [F(3,15) = 16.5; p < 0.05] (Fig. 5, top). Post hoc comparisons indicated that NTX (0.01–0.032 mg/kg) significantly attenuated scratching between time points 1 and 2.5 h after injection of morphine (p < 0.05). In contrast, i.m. diphenhydramine at any dose (0.1–10 mg/kg) did not significantly attenuate i.t. morphine-induced scratching in the same group of monkeys (Fig. 5, bottom). Furthermore, a single dose (0.032 mg/kg) of NTX pretreatment significantly blocked i.v. morphine (1 mg/kg)-induced scratching (i.e., from mean 326 scratches/15 min to mean 19 scratches/15 min). Neither Q-NTX (0.032–0.32 mg/kg) nor diphenhydramine (0.32–3.2 mg/kg) pretreatment significantly changed i.v. morphine-induced scratching (data not shown).
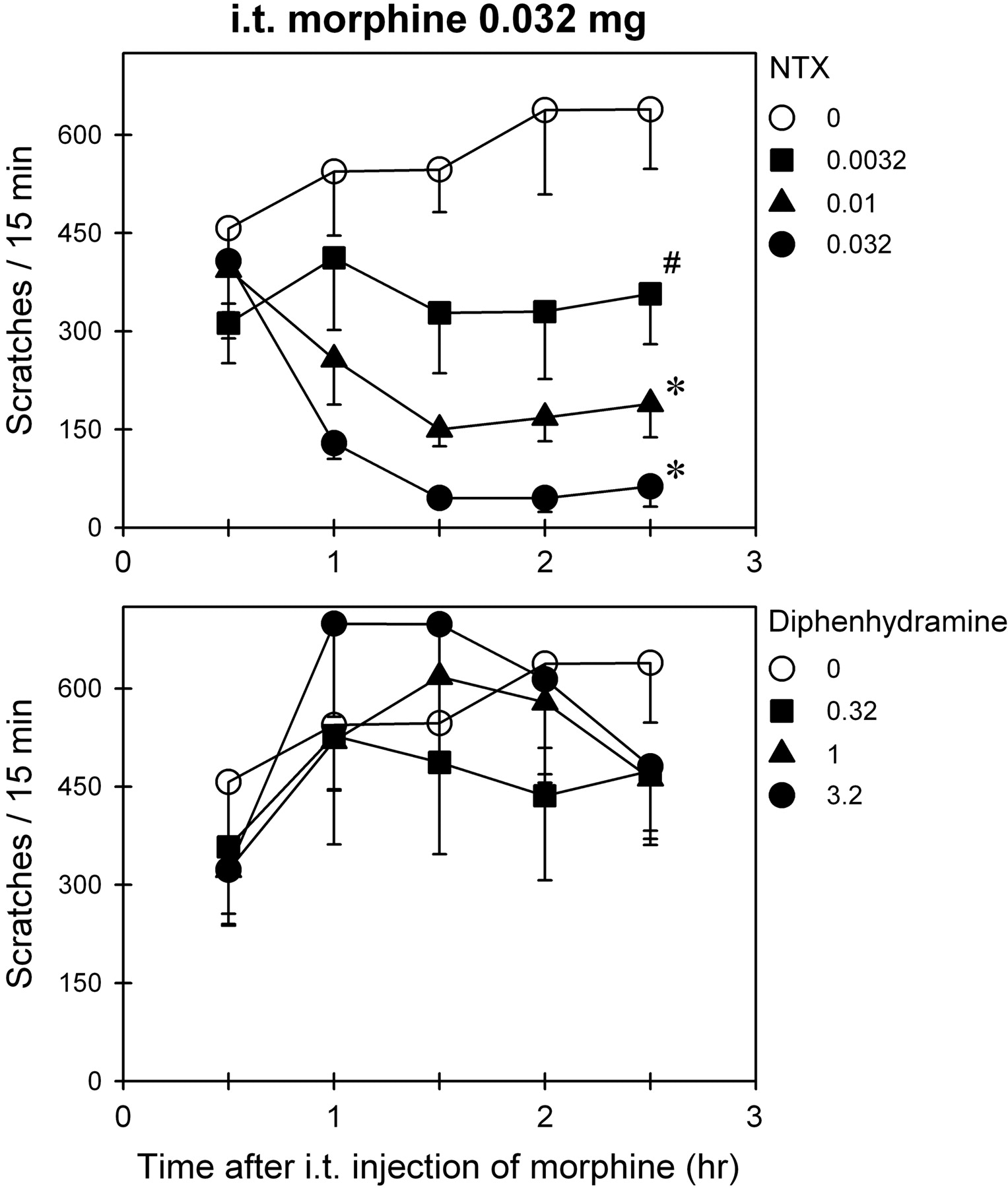
Effects of NTX or diphenhydramine on i.t. morphine-induced scratching responses. Abscissae, time in hours after i.t. administration of morphine (0.032 mg). Ordinates, scratches per 15 min. Each value represents mean ± S.E.M. (n = 6). Symbols represent different experimental conditions for the same monkeys. Either NTX or diphenhydramine (milligrams per kilogram) was given intramuscularly at 45 min after i.t. morphine. The asterisk represents a significant difference from the vehicle condition between time points 1 and 2.5 h (*, p < 0.05). The # represents a significant difference from the vehicle condition from time points 1.5 to 2.5 h (p < 0.05). The data of diphenhydramine 0.1 and 10 mg/kg are not shown for the sake of clarity.
Figure 6 illustrates the effects of MOR-, KOR-, and DOR-selective antagonists on i.t. morphine-induced scratching. There are significant differences in the inhibition of scratching by various antagonists [F(3,15) = 17.4; p < 0.05]. Post hoc comparisons indicated that administration of a MOR antagonist (clocinnamox, 0.1 mg/kg) significantly blocked i.t. morphine-induced scratching (p < 0.05). However, both KOR and DOR antagonists (nor-binaltorphimine, 3.2 mg/kg; and naltrindole, 1 mg/kg) failed to attenuate scratching induced by i.t. morphine.
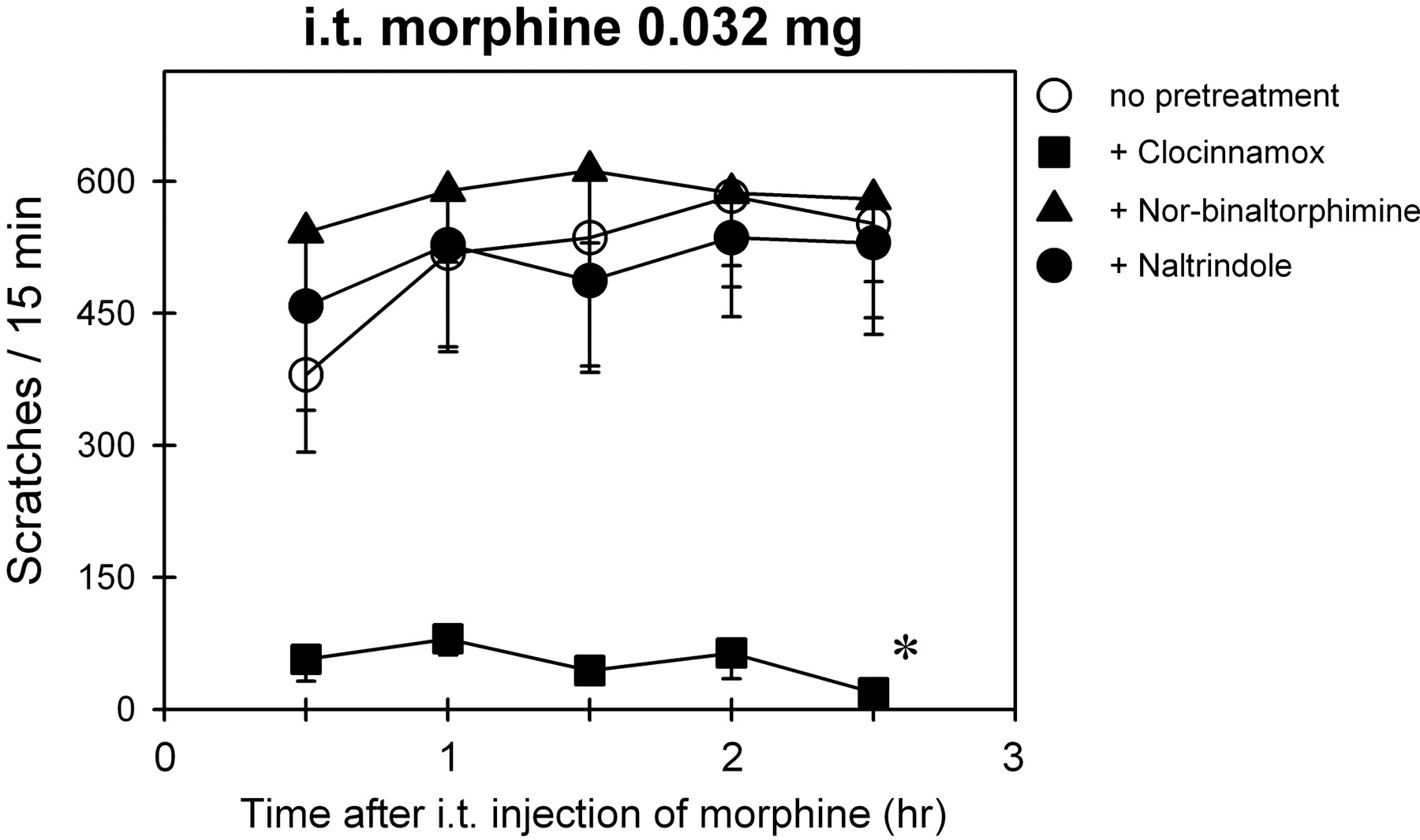
Effects of MOR-, KOR-, and DOR-selective antagonists on i.t. morphine-induced scratching responses. Abscissae, time in hours after i.t. administration of morphine (0.032 mg). Ordinates, scratches per 15 min. Each value represents mean ± S.E.M. (n = 6). Symbols represent different experimental conditions for the same monkeys. A MOR antagonist (clocinnamox, 0.1 mg/kg) or KOR antagonist (nor-binaltorphimine, 3.2 mg/kg) was given intramuscularly 24 h before i.t. morphine. A DOR antagonist (naltrindole, 1 mg/kg) was given intramuscularly 30 min before i.t. morphine. The asterisk represents a significant difference from the baseline condition (no pretreatment) between time points 0.5 and 2.5 h (*, p < 0.05). See Materials and Methods for other details.
Discussion
The present study demonstrated that systemic administration of MOR agonists produced scratching, but KOR and DOR agonists did not evoke scratching (Table 1). Doses of U-50488H and SNC80 used in this study are active in behavioral assays, such as drug discrimination and antinociception, in monkeys (Butelman et al., 1993; Negus et al., 1998). This finding may suggest that MOR selectively plays an important role in opioid-induced pruritus in primates. Such a notion can be supported by other reports. First, microinjection of U-50488H and DPDPE (a DOR agonist) into the medullary dorsal horn did not produce facial scratching in monkeys (Thomas et al., 1992). Second, i.t. administration of U-50488H and SNC80 did not evoke scratching in monkeys (Ko et al., 2003b). Third, the antagonist potency of nalmefene, an opioid receptor antagonist, confirms that MOR mediates i.t. morphine-induced scratching (Ko and Naughton, 2000). Last, a MOR antagonist, but not KOR or DOR antagonists, blocked i.t. morphine-induced scratching (Fig. 6). These findings strongly indicate that activation of MOR, not other opioid receptor types, mainly mediates itch evoked by opioid analgesics in primates.
The site of pruritic action of MOR can be studied by using pharmacological approaches. In the present study, only doses of i.v. morphine up to 0.32 to 1.8 mg/kg produced moderate scratching. This potency difference (>300-fold) between i.v. (1 mg/kg) and i.t. morphine (0.032 mg)-induced scratching could be due to the limited distribution of morphine in the spinal cord after i.v. administration. Opioid peptide analogs generally have difficulty crossing the blood-brain barrier to any appreciable extent (Omoniyi et al., 2000). If the pruritic action of MOR agonists is mainly spinally mediated, it is expected that peptidic compounds would have a larger potency difference between i.t. and i.v. administration. Our data illustrate that a peptidic MOR agonist, DAMGO, was very potent in evoking intense scratching when it was given intrathecally (i.e., 0.0032 mg). However, i.v. administration of DAMGO up to 1 mg/kg (>3000-fold) did not increase scratching (Fig. 3). These findings strongly suggest the involvement of spinal MOR in pruritus. In addition, the greater magnitude of scratching responses evoked by i.t. DAMGO than morphine could be due to that DAMGO has higher efficacy than morphine at MOR (Alt et al., 1998). It is worth noting that the magnitudes of scratching evoked by i.t. MOR agonists are much larger than those evoked by systemic administration of MOR agonists in monkeys (Ko et al., 2003a,b; Fig. 3 in the present study). This observation is consistent with clinical reports that the incidence of pruritus is low, approximately 1%, after systemic administration; and it increases to 20 to 90% in patients who receive spinal administration of opioid analgesics (Krajnik and Zylicz, 2001; Twycross et al., 2003).
Another approach to verify the role of central MOR in itch is to examine whether peripherally acting opioid receptor antagonists can modulate MOR agonist-induced scratching. Q-NTX is an N-methylated derivative of naltrexone. Although it is an effective antagonist in vitro, systemic Q-NTX is ineffective in precipitating withdrawal in morphine-dependent monkeys at doses up to one thousand times larger than the effective dose of NTX (Valentino et al., 1983). Such large differential potency could be explained by the fact that most quaternary compounds have poor access to the central nervous system after systemic administration (Brown and Goldberg, 1985). In the present study, NTX potently attenuated i.v. fentanyl-induced scratching. However, in the same dose range, Q-NTX was not effective (Fig. 2). This differential effectiveness between NTX and Q-NTX can also be found in i.v. morphine-induced scratching. The doses of Q-NTX used in this study are active in peripheral opioid receptor-mediated actions (Ko et al., 1998b; Butelman et al., 2004). In our pilot study, i.t. pretreatment with a single dose of Q-NTX (0.032 mg/kg) could significantly attenuate i.v. morphine (1 mg/kg)-induced scratching in monkeys (our unpublished observations). However, systemic administration of Q-NTX 0.032 mg/kg was not effective in attenuating MOR agonist-induced scratching. These findings further suggest that central MOR mediates scratching evoked by systemic administration of MOR agonists in monkeys.
The dose-response curve of i.v. morphine for scratching responses seems to be bell-shaped (Fig. 4), which is similar to the scratching profile evoked by s.c. morphine (Ko et al., 2003a). An early study also reports a similar observation that a single dose of i.m. morphine partially reduced ongoing facial scratching in another species, Macaca fascicularis, of monkey (Thomas et al., 1993b). However, the dose-response curve for i.t. morphine-induced scratching in monkeys seems to reach a plateau, which is similar to clinical observations (Palmer et al., 1999; Ko and Naughton, 2000). It is possible that high doses of systemic morphine produce general suppression of behaviors. Nevertheless, the mechanisms underlying the antipruritic action of high doses of systemic morphine need to be further studied. In contrast, i.v. administration of other MOR agonists, such as fentanyl, can only establish the ascending part of dose-response curve for scratching due to potential respiratory arrest (Fig. 1). The potencies of fentanyl-like analogs in producing scratching versus antinociception are similar under these experimental conditions (Ko et al., 2002). Although the antinociceptive potency of MOR agonists may depend on different forms and intensities of pain, our findings parallel closely with clinical studies that analgesic doses of MOR agonists produce side effects such as pruritus in humans (Ellis et al., 1990; Mackersie et al., 1991).
Systemic administration of NTX, but not diphenhydramine, attenuated i.t. and i.v. morphine-induced scratching. More important, the durations of scratching evoked by i.v. administration of fentanyl-like analogs are similar to their durations of antinociception and respiratory depression in monkeys (Ko et al., 2002). Both findings indicate that histamine does not seem to be a mechanism of opioid-induced itch and further support clinical studies that antihistamines are not effective in treating opioid-induced pruritus (Dunteman et al., 1996; Twycross et al., 2003). Tachyphylaxis quickly develops to histamine-induced itch and antihistamines are of limited use for treating pruritus derived from systemic disease and most dermatoses (Krajnik and Zylicz, 2001; Szarvas et al., 2003; Twycross et al., 2003). The role of histamine may be minimal in chronic itch. Interestingly, itch-specific primary afferents and spinal neurons have been discovered based on histamine-evoked itch (Schmelz et al., 1997; Andrew and Craig, 2001). It would be important to investigate whether these neurons respond to other pruritogenic agents such as MOR agonists.
Systemic administration of NTX is very potent and effective in preventing or reversing scratching evoked by MOR agonists (Figs. 2 and 5). NTX displays higher affinity for MOR and low doses of NTX are sufficient to antagonize MOR-mediated actions (Ko et al., 1998a). Therefore, the effectiveness of opioid receptor antagonists is expected when itch is evoked by MOR opioid analgesics or mediated by endogenous opioid peptides. Our finding that pretreatment with a MOR-selective antagonist blocked i.t. morphine-induced scratching further supports this notion. It is worth noting that opioid receptor antagonists produce parallel rightward shifts in dose-response curves of morphine-induced scratching (Ko and Naughton, 2000; Lee et al., 2003). These observations indicate that NTX or nalmefene antagonism is competitive and reversible at MOR and could be used to distinguish selective opioid actions. In contrast, KOR agonists have been reported to have antipruritic actions (Cowan and Gmerek, 1986; Togashi et al., 2002; Ko et al., 2003a). KOR agonists such as U-50488H produce downward shifts in dose-response curves of morphine-induced scratching, and the antipruritic actions of U-50488H can be reversed by a KOR-selective antagonist (Ko et al., 2003a). These observations indicate that KOR agonists do not produce MOR antagonism, but inhibit MOR-mediated itch through KOR activation. To date, pharmacological studies in monkeys have demonstrated that both opioid receptor antagonists and KOR agonists are effective in attenuating MOR agonist-induced itch (Ko and Naughton, 2000; Ko et al., 2003a; the present study). It is important to further investigate neurobiological mechanisms between MOR and KOR in itch-specific neurons.
In summary, this study demonstrates that MOR, not KOR, DOR, or histamine, mediates scratching evoked by opioid analgesics in monkeys. In particular, the scratching behavior represents an in vivo indicator of central MOR activation, which provides a pharmacological basis for characterizing newly developed opioid analgesics. Using pharmacological approaches, scratching in monkeys can be further used to identify specific receptors involved in pruritus and facilitate development of antipruritics targeting these receptors.
Acknowledgments
We thank John Busenbark, Wayne Yang, and Chenzhi Lu for excellent technical assistance.
Footnotes
-
This study was supported by U.S. Public Health Service Grant R01-DA13685 (to M.C.H.K.).
-
DOI: 10.1124/jpet.103.061101.
-
ABBREVIATIONS: KOR, κ opioid receptor; DOR, δ opioid receptor; MOR, μ opioid receptor; U-50488H, trans-(±)-3,4-dichloro-N-methyl-N-(2-[1-pyrrolidinyl]-cyclohexyl)-benzeneacetamide; SNC80, (+)-4-[(αR)-α-[2S,5R)-4-allyl-2,5-dimethyl-1-piperazinyl]-3-methoxybenzyl]-N,N-diethylbenzamide; NTX, naltrexone; Q-NTX, quaternary naltrexone; DAMGO, [d-Ala2,N-Me-Phe4,Gly5-ol]-enkephalin.
- Received October 5, 2003.
- Accepted March 25, 2004.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}