


Abstract
The dopamine transporter (DAT) is a critical recognition site for cocaine and contributes to its significant abuse liability. Accordingly, the development of compounds that target the DAT represents a logical approach in the pharmacological treatment of cocaine abuse. The present study characterized the effects of DAT inhibitors as pretreatments in rhesus monkeys trained to self-administer cocaine under a second-order schedule of i.v. drug delivery. The drugs also were substituted for cocaine to characterize their effectiveness in maintaining drug self-administration. Positron emission tomography neuroimaging with [18F]8-(2-[18F]fluoroethyl)-2β-carbomethoxy-3β-(4-chlorophenyl) nortropane established the DAT occupancy associated with behaviorally relevant doses of each drug. The drugs studied included a selective DAT inhibitor, [1-(2[bis(4-fluorophenyl-) methoxy]ethyl)-4-(3-phenylpropyl)piperazine] bimesylate hydrate (GBR 12909); an inhibitor with equal potency at dopamine and norepinephrine transporters, [3β-(4-chlorophenyl)tropane-2β-(3-phenylisoxazol-5-yl)] HCl (RTI-177); and a nonselective inhibitor of dopamine, norepinephrinem and serotonin transporters, [(-)-3β-(3′-methyl-4-chlorophenyl)tropane-2β-carboxylic acid methyl ester] tartrate (RTI-112). All drugs produced dose-related reductions in cocaine self-administration. Doses of GBR 12909 and RTI-177 that reduced responding by 50% (ED50) resulted in DAT occupancies of 67 ± 5 and 73 ± 5%, respectively. In contrast, DAT occupancy was below the limit of detection for the ED50 dose of RTI-112. Both GBR 12909 and RTI-177 reliably maintained drug self-administration, and DAT occupancies at doses that maintained peak rates of responding were 57 ± 1 and 92 ± 7%, respectively. In contrast, RTI-112 failed to maintain robust drug self-administration in any subject. The results indicate that selective DAT inhibitors may require high DAT occupancy to reduce cocaine self-administration and maintain drug self-administration. Moreover, the behavioral profile of DAT inhibitors may be influenced by actions at other monoamine transporters.
Despite extensive efforts directed toward the development of medications to treat cocaine abuse, no effective pharmacotherapy is currently in clinical use. The therapeutic approach of substitute agonist or replacement medication has been successful in the context of methadone maintenance for heroin dependence and nicotine replacement for tobacco use. These positive outcomes, combined with recent advances in the understanding of the neuropharmacology of cocaine, support efforts to develop a similar type of medication for cocaine abuse. Of the various types of medications being pursued, dopamine transporter (DAT) inhibitors represent a promising approach in drug development (Mello and Negus, 1996; Carroll et al., 1999; Howell and Wilcox, 2001). The DAT is an important recognition site for cocaine and likely mediates its reinforcing effects that contribute to significant abuse liability (Ritz et al., 1987; Kuhar et al., 1991; Woolverton and Johnson, 1992). The affinities of several cocaine-like drugs for the DAT correlate well with their potencies for supporting self-administration behavior (Ritz et al., 1987; Bergman et al., 1989; Wilcox et al., 1999). Importantly, a variety of preclinical studies in nonhuman primates provide evidence that DAT inhibitors can effectively attenuate cocaine self-administration (Glowa et al., 1995; Nader et al., 1997; Howell et al., 2000; Wilcox et al., 2002). Hence, the development of compounds that target the DAT represents a logical approach for the pharmacological treatment of cocaine abuse.
DAT occupancy seems to play a significant role in the subjective and reinforcing effects of cocaine. Doses of cocaine within the range used by humans resulted in DAT occupancy between 67 and 69% in baboons (Volkow et al., 1996). Doses of cocaine that maintained peak response rates in drug self-administration studies resulted in DAT occupancy between 65 and 76% in rhesus monkeys (Wilcox et al., 2002). Similarly, doses of 0.1 and 1.0 mg/kg cocaine occupied 53 and 87% of the transporters, respectively, in rhesus monkeys (Votaw et al., 2002), confirming that high levels of DAT occupancy are associated with behaviorally active doses of cocaine. In human cocaine users, a significant correlation was observed between DAT occupancy and the subjective effects of cocaine (Volkow et al., 1997). In addition, DAT occupancy has been determined for DAT inhibitors shown to be effective in reducing cocaine self-administration. Doses of GBR 12909 that decreased cocaine self-administration in rhesus monkeys resulted in DAT occupancy greater than 50% in baboons (Villemagne et al., 1999). Similarly, doses of RTI-113 that decreased cocaine self-administration in rhesus monkeys resulted in DAT occupancies between 72 and 84% (Wilcox et al., 2002). Collectively, these results indicate that DAT occupancy is an important determinant of the reinforcing effects of cocaine and of the effectiveness of DAT inhibitors to reduce cocaine self-administration.
The present study extended these findings by characterizing the effects of several DAT inhibitors with varying degrees of selectivity for the DAT. Previous in vitro uptake assays have characterized their potency and selectivity at monoamine transporters (Anderson, 1989; Kuhar et al., 1999; Table 1). The DAT inhibitors were administered as pretreatments in rhesus monkeys trained to self-administer cocaine. In addition, the DAT inhibitors were substituted for cocaine to characterize their effectiveness in maintaining drug self-administration. PET neuroimaging established the level of DAT occupancy associated with behaviorally relevant doses of each DAT inhibitor. Based on the unique behavioral profile exhibited by a nonselective monoamine transporter inhibitor, the present study also established the level of serotonin transporter (SERT) occupancy associated with a behaviorally relevant dose of the drug. Direct, within-subject comparisons between drug effects on behavior and in vivo transporter occupancy will have important implications for understanding the neuropharmacology of cocaine. Moreover, the results obtained should lead to a better understanding of the mechanisms underlying cocaine self-administration and direct efforts to develop pharmacotherapies to treat cocaine abuse.
Uptake inhibition values (IC50, nM) in rat tissue at DAT, SERT, and NET
Materials and Methods
General Methods
Subjects. Six female and five male adult rhesus monkeys (Macaca mulatta) weighing 7.5 to 13.0 kg were used as subjects. Each subject was housed individually and fed Purina monkey chow (Ralston Purina, St. Louis, MO), fruits, and vegetables. Water was continuously available. Animal care procedures strictly followed the National Institutes of Health Guide for the Care and Use of Laboratory Animals and were approved by the Institutional Animal Care and Use Committee of Emory University.
Surgery. Each subject was prepared surgically with a chronic indwelling venous catheter under sterile surgical conditions using a technique described previously (Wilcox et al., 2002). Preoperative antibiotics (25 mg/kg Rocephin or 25 mg/kg Cefazolin) were given on the day of surgery to help prevent infection. A silicone catheter (0.65 mm i.d., 1.75 mm o.d.; Access Technologies, Skokie, IL) was implanted under a combination of Telazol (4.0 mg/kg) and isoflurane anesthesia using aseptic techniques. The proximal end of the catheter terminated in the vena cava above the right atrium, and the distal end was routed under the skin and attached to a subcutaneous vascular access port (Access Technologies) located in the center of the lower back. After surgery, the subject was returned to its home cage and received Banamine (1.0 mg/kg i.m.) every 6 h for 24 h postoperatively to reduce pain and discomfort associated with surgery. Catheters were flushed daily with 1.0 ml of heparinized (100 units/ml) saline to maintain patency.
Drugs. Cocaine HCl (National Institute on Drug Abuse, Rockville, MD), [3β-(4-chlorophenyl)tropane-2β-(3-phenylisoxazol-5-yl)] HCl (RTI-177), and [(-)-3β-(3′-methyl-4-chlorophenyl)tropane-2β-carboxylic acid methyl ester] tartrate (RTI-112) were synthesized as reported previously (Carroll et al., 1992; Kotian et al., 1995) and were dissolved in 0.9% saline. [1-(2[Bis(4-fluorophenyl-) methoxy-]ethyl}-4-(3-phenylpropyl)piperazine] bimesylate hydrate (GBR 12909) (Kenner Rice, Laboratory of Medicinal Chemistry, National Institute of Diabetes and Digestive and Kidney Diseases, Rockville, MD) was dissolved in sterile water. All drug doses were determined as salts. [18F][8-(2-[18F]fluoroethyl)-2β-carbomethoxy-3β-(4-chlorophenyl)nortropane] (FECNT) was used as a PET ligand to label the DAT (Goodman et al., 2000). [11C][2β-carbo[11C]methoxy-3β-(4′-((Z)-2-iodoethenyl)phenyl)nortropane] (ZIENT) was used as a PET ligand to label the SERT (Goodman et al., 2003).
Behavioral Methods
Apparatus. During behavioral testing, each monkey was seated in a commercially available primate chair (Primate Products, Redwood City, CA). A response panel with one lever was mounted on the front of chair. Located above the lever in the center of the response panel were red and white stimulus lights. Once the monkey was seated in the chair, a Huber needle (Access Technologies) was inserted into the venous access port. The polyvinyl chloride tubing attached to the Huber needle was connected to a motor-driven syringe (Coulbourn Instruments, Allentown, PA) located outside of the chamber containing the drug solution. A volume of 2.0 ml/infusion was delivered over 7 s. Testing during daily 1-h sessions occurred in a ventilated, sound-attenuating chamber. IBM-compatible computers controlled experimental events and recorded data.
Procedure. Subjects responded for i.v. infusions of cocaine under a second-order schedule of reinforcement, as described previously (Wilcox et al., 2002). The training dose of cocaine was 0.1 mg/kg/infusion. When the daily session began, the red light on the response panel was illuminated and responding resulted in the delivery of a drug infusion and brief 2-s illumination of the white light. Initially, the fixed ratio was 1 and gradually increased to fixed ratio 20. Ultimately, a second-order schedule of reinforcement was in effect, with the first fixed ratio 20 completed after 10 min (fixed interval 10 min), resulting in a drug infusion. Fixed ratio 20 components completed within the 10-min fixed interval resulted in illumination of the white light for 2 s. There was a 30-s limited hold for completion of the first fixed ratio 20 after the fixed interval 10 min had elapsed, and a drug infusion was not delivered if the limited hold expired. Drug infusions were signaled by a change in the lights from red to white for 15 s. After each drug infusion, there was a 1-min time-out during which responding on the lever had no programmed consequences. A total of five infusions could be delivered during a daily session comprising five consecutive fixed interval 10-min components.
The training sequence remained in effect until responding for cocaine was stable (<20% variance in daily response rate over five consecutive days), after which saline was substituted for cocaine until responding decreased to below 30% of responding for the training dose of cocaine. After saline extinction, the maintenance dose (0.1 mg/kg/infusion) of cocaine was reinstated and responding was allowed to stabilize. For pretreatment studies, a given dose of drug was administered i.v. 15 min presession on three consecutive days, typically Tuesday, Wednesday, and Thursday. Subjects remained in the test chamber during the 15-min pretreatment period. No trend was present across treatment days for any drug dose, so daily response rate was averaged across the 3 days. Vehicle was administered on all days that subjects did not receive a drug pretreatment, and these data contributed to ongoing calculations of baseline stability. Typically, each pretreatment dose was administered on two separate occasions in quasi-random order. If responding was reduced to less than 30% of baseline, a second determination was not made. All doses of a particular drug were studied in combination with 0.1 mg/kg/infusion cocaine first. Subsequently, the maintenance dose of cocaine was changed to 0.3 mg/kg/infusion, and drug pretreatments were repeated as described above. The order of testing was RTI-177, GBR 12909, and RTI-112. After pretreatment studies, the maintenance dose of cocaine was reinstated and responding was allowed to stabilize. For drug substitution studies, each subject was allowed to self-administer several doses of each drug in quasi-random order. Substitution for each drug dose continued for at least five consecutive sessions and until responding stabilized (<20% variance in daily response rate).
Data Analysis. Response rates and cocaine intake for individual subjects were analyzed as a function of dose for each drug. Average rates of cocaine-maintained responding and average number of cocaine infusions received after pretreatment drugs were compared with values obtained after saline pretreatments using one-way analysis of variance. Post hoc comparisons were made using Dunnett's method. Drug doses predicted to reduce rates of responding by 10% (ED10) and 50% (ED50) of baseline were derived from nonlinear regression analysis of data from individual subjects.
PET Neuroimaging Methods
Apparatus. PET neuroimaging was performed at the Emory University Pet Center on a Siemens 951 scanner, which can image an entire rhesus monkey brain with a 10-cm axial field of view. A set of Ge-68 ring sources was used for attenuation correction before injection of [18F]FECNT. All images were reconstructed with measured attenuation correction, zoom factor 8, and Shepp-Logan reconstruction filter cut off at 1 cycle/cm. This produced images with an inplane pixel size of 1.17- and 8-mm resolution. The axial slice thickness was 3.375 mm. All images were decay-corrected to the time of injection. Regions of interest were manually drawn on the late images over the putamen and cerebellum. The regions of interest were then overlaid on all images to obtain time-activity curves.
Procedure. Measures of DAT occupancy for a given drug were obtained after the completion of behavioral studies with the drug in individual subjects. In drug pretreatment studies, the ED10 and ED50 doses of GBR 12909 and RTI-177 were administered as a single i.v. bolus. The ED10 dose of RTI-112 was not tested in PET studies because the ED50 dose resulted in DAT occupancy below the threshold of detection. Instead, the highest dose of RTI-112 was administered. In drug substitution studies, the dose of GBR 12909 or RTI-177 that maintained peak response rates was administered as an i.v. bolus of the total dose the animal received during its self-administration sessions (dose in milligrams per kilogram per infusion × average number of infusions per session). On days of the PET studies, subjects were immobilized in their home cage with Telazol (4.0 mg/kg) and transported to the Emory University PET Center. Subsequently, they were intubated, and anesthesia was maintained with 1.0 to 2.0% isoflurane. Subjects were positioned in the tomograph, and a 15-min transmission scan was obtained for attenuation correction; 1.5 h after isoflurane anesthesia began, a slow bolus of approximately 5.0 mCi of [18F]FECNT (specific activity 1.5 Ci/μmol) was injected over 5 to 6 min at a rate of 1.0 ml/min. Scanning began coincident with the start of injection. The initial acquisition was a 28-frame dynamic sequence starting with 30-s scans and ending with 20-min scans for a total duration of 1.5 h. At 1.5 h, a single bolus dose of a DAT inhibitor was injected, and a second dynamic sequence was acquired starting with 2-min frames and ending with 5-min frames for a total duration of 1.5 h. A separate group of three subjects participated in PET imaging experiments in both anesthetized and awake states as described previously (Howell et al., 2001, 2002) to determine the effects of isoflurane anesthesia on measures of DAT occupancy. Last, measures of SERT occupancy for the ED50 dose of RTI-112 observed in drug pretreatment studies were determined with the PET ligand [11C]ZIENT. Both ZIENT and FECNT are nortropane derivatives that exhibit similar physiochemical properties appropriate for PET imaging of transporter occupancy. ZIENT is a potent ligand for imaging the SERT (Ki = 0.05 nM) with high selectivity over DAT and NET (Goodman et al., 2003). The kinetic analysis was performed using the same programs and methods that were used to analyze the [18F]FECNT data (Wilcox et al., 2002; Votaw et al., 2002).
Data Analysis. A generalized reference tissue method was used to analyze the data (Votaw et al., 2002). Data were collected in two sections with the assumption that drug infusion changed only the k3 (Bmaxkon) rate constant (i.e., drugs compete with the FECNT for binding the DAT and decrease the apparent Bmax) from ka3 (predrug infusion) to kb3 (postdrug infusion). Five rate constants (R, k2, ka3, kb3, and k4) and the time-activity curve from the reference region (cerebellum) were used to model the putamen time-activity curve. In the modeling, it was assumed that the competing ligand did not affect the blood-brain barrier parameters (flow or extraction; R or k2) or the transporter-FECNT kinetic properties (kon, koff = k4). Thus, these parameters were kept constant over the entire experiment. A best fit (least-squares sense) set of parameters was determined using the Powell routine (Press et al., 1988). The off rate was constrained to fit within a factor of 2 of the value determined by RTI-55 drug-chase experiments (Goodman et al., 2000). After optimization, the covariance matrix was numerically determined and used to estimate the uncertainty in the fit values. From this, it was found that the variance in the k3/k4 ratio is much less than the variance in either parameter alone due to their nonvanishing covariance. Because of this, the k3/k4 ratio was taken as the measure of DAT density. The fraction of transporters occupied by the competing ligand was then estimated as 1 - (kb3/k4)/(ka3/k4). The chi-squared function (Press et al., 1988) was used to determine the goodness of fit while adjusting the model parameters of the generalized reference tissue model (Votaw et al., 2002). The error in regional uptake as determined from repeated 2-min measurements of [O-15]water has been reported to be approximately 5% (Li and Votaw, 1998). Using this as a conservative guide, the error was assumed to be 8% of the measured value for frames 2 min in duration and inversely proportional to the scan duration for other durations. Initially, the data pre- and postchase were fit assuming one binding potential for the entire scan (null hypothesis). If this fit was rejected at the p < 0.05 level and then the data were refit using separate binding potentials for pre- and post-chase. In practice, when the data are consistent with a binding potential change of more than 10%, the null hypothesis is rejected, which gives the sensitivity of this technique.
Results
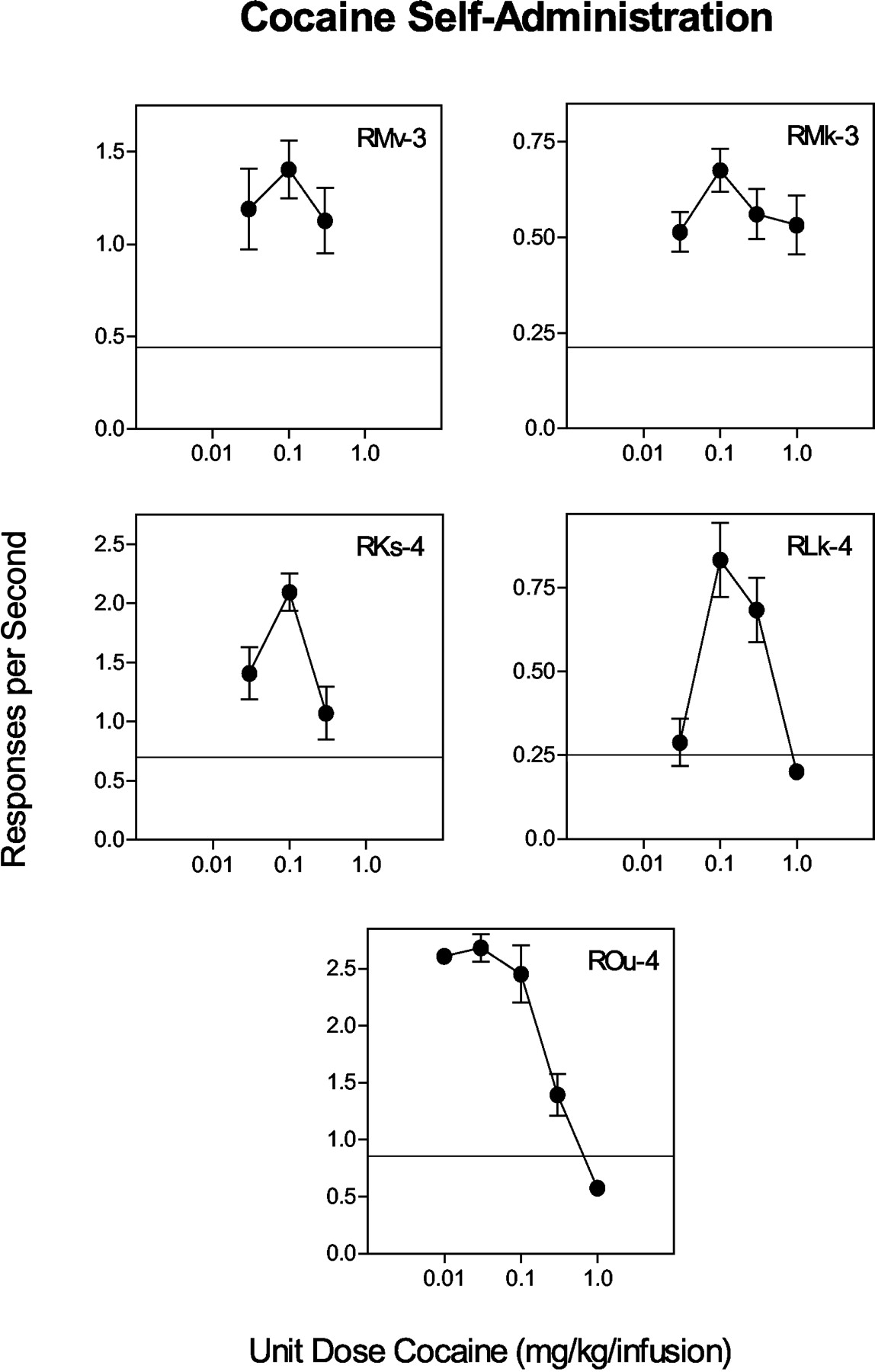
Drug Pretreatment Studies. Dose-effect curves for cocaine self-administration alone generally were characterized as inverted U-shaped functions in individual subjects (Fig. 1). Unit doses of 0.1 and 0.3 mg/kg/infusion were selected for drug pretreatment experiments because they were positioned on the peak and descending limb of the dose-effect curves, respectively. Pretreatments with the DAT inhibitors caused dose-dependent reductions in cocaine-maintained responding in all subjects tested (Figs. 2, 3, 4), and varying the maintenance dose of cocaine had no influence on the effectiveness of drug pretreatments (Figs. 2 and 3). Therefore, data were pooled for each maintenance dose of cocaine to determine ED10 and ED50 doses for drug pretreatments. Comparisons of ED50 values indicated that RTI-112 was approximately an order of magnitude more potent than RTI-177 which was approximately an order of magnitude more potent than GBR 12909 (Table 2). DAT inhibitor-induced reductions in cocaine-maintained responding were accompanied by significant reductions in cocaine intake (Table 3). No adverse behavioral effects were observed in any subject, and daily food intake and body weight were normal.

Mean (±S.D.) rates of responding (responses per second) maintained by cocaine self-administration in individual subjects. Data were derived from the last 5 days of each condition that met stability criterion of <20% variation. Solid lines indicate the upper limit for responding during saline extinction.

GBR 12909-induced reductions in cocaine self-administration: relationship to DAT occupancy. The line graph (left ordinate) depicts rates of cocaine-maintained responding. Open symbols represent rates of responding maintained by 0.1 mg/kg/infusion (inf) (circles) or 0.3 mg/kg/inf (squares). Closed symbols represent rates of responding for 0.1 mg/kg/inf (circles) or 0.3 mg/kg/inf (squares) after pretreatments of GBR 12909 as indicated on the abscissa. Each data point represents one or two pretreatment experiments, each consisting of three consecutive daily cocaine self-administration sessions performed under the influence of the indicated pretreatment dose of GBR 12909. Error bars represent the standard error of the mean for group data (A) and the standard deviation for data from individual subjects (B). All pretreatment doses of GBR 12909 tested had significant effects on responding maintained by 0.1 and 0.3 mg/kg/inf (p < 0.001). The bar graphs (right ordinate) depict percentage of DAT occupancy by the doses of GBR 12909 indicated on the abscissa. The asterisks represent occupancy values that were below the limit of detection.

RTI-177-induced reductions in cocaine self-administration: relationship to DAT occupancy. Otherwise, as in Fig. 2. Pretreatments with RTI-177 had a significant effect on cocaine-maintained responding (p < 0.05). Post hoc comparisons made using Dunnett's method revealed that only the highest pretreatment dose of RTI-177 (0.3 mg/kg) had significant effects on responding maintained by 0.1 and 0.3 mg/kg/infusion (inf) (p < 0.01). The bar graphs (right ordinate) depict percentage of DAT occupancy by the doses of RTI-177 indicated on the abscissa. The asterisks represent occupancy values that were below the limit of detection.

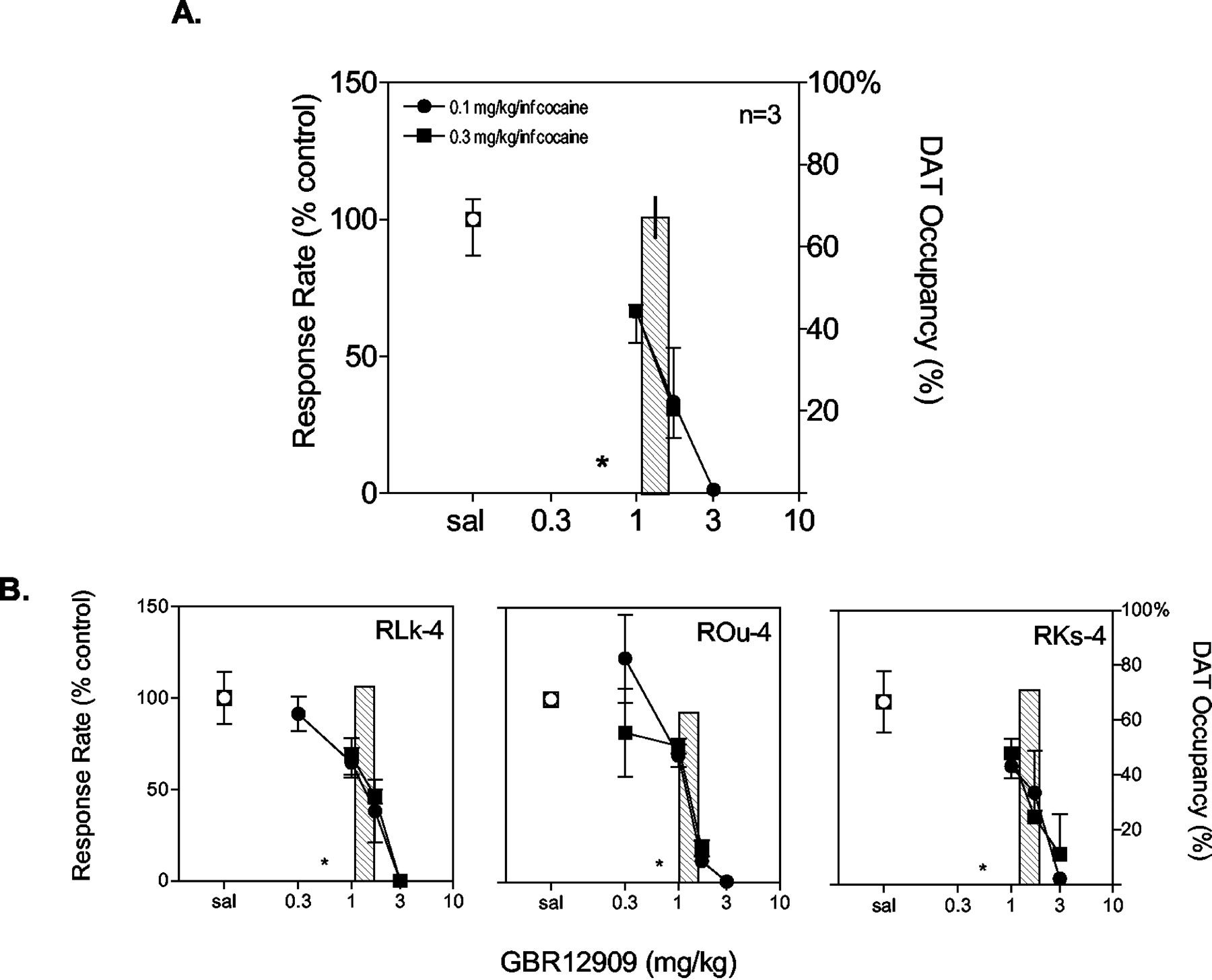
RTI-112-induced reductions in cocaine self-administration: relationship to DAT occupancy. Otherwise, as in Fig. 2. Pretreatments with RTI-112 had a significant effect on cocaine-maintained responding (p < 0.01). Post hoc analysis using Dunnett's method revealed that a pretreatment with 0.03 mg/kg RTI-112 had a significant effect on cocaine-maintained responding (p < 0.01). The bar graphs (right ordinate) depict percent DAT occupancy by the doses of RTI-112 indicated on the abscissa. The asterisks represent occupancy values that were below the limit of detection.
Relationship between reductions in cocaine self-administration and DAT occupancy
Mean ± S.D. cocaine intake (infusions per session)
PET neuroimaging experiments were conducted to determine the percentage of DAT occupancy associated with the ED10 and ED50 doses of drug pretreatments derived from behavioral experiments. The ED10 and ED50 values were determined for individual subjects rather than using the same group mean for all subjects. The ED10 doses of GBR 12909 and RTI-177 resulted in DAT occupancies below the threshold (<10%) of detection using this methodology (Figs. 2 and 3; Table 2). In contrast, the ED50 doses of GBR 12909 and RTI-177 resulted in DAT occupancies of 67 and 73%, respectively, for the group. The ED10 dose of RTI-112 was not tested in PET studies because the ED50 dose resulted in DAT occupancy below the threshold of detection (Fig. 4; Table 2). However, the highest dose of RTI-112 tested in each subject resulted in DAT occupancy of 73% for the group. Time-activity curves obtained in subject RMv-3 illustrate uptake of [18F]FECNT and its displacement by RTI-112 (Fig. 5). Given that the ED50 dose of RTI-112 resulted in DAT occupancy below the threshold of detection, additional PET neuroimaging experiments were conducted in the same subjects to determine the percentage of SERT occupancy. The ED50 dose of RT-112 resulted in SERT occupancies of 99, 88, and 64% for subjects ROu-4, RKs-4, and RMv-3, respectively (group mean 84% ± 13).

Typical [18F]FECNT time-activity curves for the putamen and cerebellum in subject RMv-3. The data have been decay-corrected to the time of injection. Note displacement of activity from the putamen after a high dose of RTI-112 (0.03 mg/kg) but not by the ED50 dose of RTI-112 (0.01 mg/kg).
Drug Substitution Studies. The DAT inhibitors were substituted for cocaine in drug self-administration studies in order to characterize their reinforcing effects. Both GBR 12909 and RTI-177 reliably maintained drug self-administration at levels greater than those maintained by saline in all subjects (Fig. 6). Moreover, the shape of the dose-effect curves resembled an inverted-U shape function typical of psychomotor stimulants. However, rates of responding were lower than those maintained by the training dose of cocaine (0.1 mg/kg/infusion) in two of three subjects substituted with GBR 12909 and in all three subjects substituted with RTI-177. Interestingly, RTI-112 failed to maintain robust drug self-administration in any subject (Fig. 6). In subject ROu-4, responding was intermediate between that maintained by saline and the training dose of cocaine. In subject RSu-4, only the highest dose tested maintained responding greater than that maintained by saline. In subject RLk-3, no dose maintained responding greater than that maintained by saline.

Self-administration of GBR 12909, RTI-177, and RTI-112: relationship to DAT occupancy. Response rates (responses per second) for GBR 12909, RTI-177, and RTI-112 as a function of drug dose under a second-order schedule of i.v. drug self-administration in individual subjects. The unbroken lines indicate mean rates of responding maintained by the training dose of cocaine (0.1 mg/kg/infusion). Dashed lines indicate the upper limit for responding during saline extinction. Each data point was determined on a single occasion and is the mean (±S.D.) of the last five sessions in a condition. Numbers in parentheses indicate percentage of DAT occupancy at doses that maintained peak rates of responding.
PET neuroimaging experiments were conducted to determine the percentage of DAT occupancy associated with doses of GBR 12909 and RTI-177 that maintained peak rates of responding. Similar experiments were not conducted with RTI-112 due to its marginal reinforcing effects and the outcome of earlier experiments showing no significant DAT occupancy at the ED50 dose in drug pretreatment studies. Unit doses of GBR 12909 and RTI-177 that maintained peak rates of responding were identified for individual subjects and then the average total dose the animal received during its self-administration sessions was determined and administered as a bolus injection in PET neuroimaging experiments (Table 4). For GBR 12909, the DAT occupancy was 57% (range 41–69%), and for RTI-177, the DAT occupancy was 92% (range 85–98%) for the group (Fig. 6). Hence, the percentage of DAT occupancy at doses that maintained peak rates of responding was greater for RTI-177 compared with GBR 12909.
Relationship between drug self-administration and DAT occupancy
Effect of Isoflurane Anesthesia on DAT Occupancy. PET neuroimaging experiments were conducted in a separate group of three subjects to determine the effect of isoflurane anesthesia on measures of percentage of DAT occupancy. The analysis was restricted to RTI-177 and was conducted in the same subjects both awake and anesthetized on separate occasions. There was no significant difference in measures of DAT occupancy by RTI-177 during awake and anesthetized states (Table 5). The largest difference in DAT occupancy for any animal tested was 9% (range 2–9%).
Effects of isoflurane anesthesia on DAT occupancy by RTI-177 A separate group of three rhesus monkeys participated in PET neuroimaging experiments in both awake and anesthetized states. Since these animals did not participate in behavioral studies, the dose of RTI-177 (0.11 mg/kg) was determined using the mean ED50 derived from behavioral experiments in other subjects (Table 2). No significant difference in measures of DAT occupancy was observed between the awake and anesthetized states
Discussion
The present study compared the effects of several DAT inhibitors with varying selectivity for monoamine transporters in drug self-administration protocols. Direct, within-subject comparisons were made between drug effects on behavior and in vivo DAT occupancy measured with PET neuroimaging. Two drugs with selectivity for DAT over SERT (GBR 12909 and RTI-177) exhibited high levels of DAT occupancy at doses that produced robust decreases in cocaine self-administration. In contrast, a mixed action inhibitor of DAT and SERT (RTI-112) did not exhibit levels of DAT occupancy above the threshold of detection at a dose that significantly reduced cocaine self-administration. Importantly, the same dose of RTI-112 exhibited high levels of SERT occupancy, showing apparent in vivo selectivity for SERT over DAT at a behaviorally relevant dose. In drug substitution studies, GBR 12909 and RTI-177 reliably maintained drug self-administration in all subjects. In contrast, RTI-112 did not function as a robust reinforcer in any subject. Collectively, the results indicate that the behavioral profile of DAT inhibitors may be influenced by actions at other monoamine transporters.
Each of the DAT inhibitors investigated produced dose-related reductions in cocaine self-administration, consistent with a variety of studies in nonhuman primates. For example, the phenyltropane RTI-113 effectively decreased cocaine self-administration in squirrel monkeys (Howell et al., 2000) and rhesus monkeys (Wilcox et al., 2002) trained under a second-order schedule. RTI-113 maintained its effectiveness when the unit dose of cocaine was increased, indicating that reductions in cocaine self-administration could not be surmounted by a higher dose of cocaine. In the present study, RTI-177 also maintained its effectiveness at multiple unit doses of cocaine. Similar results have been observed with the phenyltropane PTT in rhesus monkeys trained under a fixed interval schedule (Nader et al., 1997). The effectiveness of DAT inhibitors to reduce cocaine self-administration extends to the phenylpiperazine derivative GBR 12909. In rhesus monkeys trained under a fixed ratio schedule, GBR 12909 dose dependently decreased response rates and drug intake (Glowa et al., 1995). Last, nonselective monoamine transporter inhibitors, including cocaine, have been effective in reducing cocaine self-administration in rhesus monkeys. Administration of cocaine as a continuous infusion during self-administration sessions reduced cocaine-maintained responding in a dose-related manner (Glowa and Fantegrossi, 1997), although consistent effects were not observed in another study (Panlilio et al., 1998). Indatraline is another example of a nonselective monoamine transporter inhibitor that produced dose-dependent decreases in cocaine self-administration over a broad range of cocaine doses (Negus et al., 1999).
High levels of DAT occupancy were observed for doses of GBR 12909 and RTI-177 that produced robust decreases in cocaine self-administration. At doses that decreased rates of responding by 50%, DAT occupancy was approximately 70% for both compounds. Similar levels of DAT occupancy have been reported for RTI-113 at doses that suppressed cocaine self-administration (Wilcox et al., 2002). Likewise, doses of GBR 12909 that decreased cocaine self-administration in rhesus monkeys (Glowa et al., 1995) resulted in DAT occupancies greater than 50% in baboons (Villemagne et al., 1999). Interestingly, DAT occupancy for the ED50 dose of RTI-112 was below the threshold of detection in the present study. A higher dose resulted in DAT occupancy greater than 70%, clearly demonstrating that RTI-112 can effectively displace [18F]FECNT. It is unlikely that pharmacokinetic considerations were responsible for the results obtained given that RTI-112 and RTI-I77 have similar onset and duration of action in rodent locomotor assays (Kimmel et al., 2001). Importantly, the ED50 dose of RTI-112 resulted in SERT occupancy of approximately 84%. Hence, reductions in cocaine self-administration induced by intermediate doses of RTI-112 were likely mediated through its actions at the SERT. In support of this interpretation, selective SERT inhibitors can attenuate cocaine-induced stimulant and reinforcing effects (Kleven and Woolverton, 1993; Howell and Byrd, 1995; Czoty et al., 2002), increases in extracellular dopamine (Czoty et al., 2002), and activation of prefrontal cortex (Howell et al., 2002). Note that a higher dose of RTI-112 resulted in DAT occupancy greater than 70%. Therefore, it is reasonable to speculate that DAT-mediated effects also contributed to the marked reductions in cocaine self-administration observed at the highest dose of RTI-112.
Given that RTI-112 also has high affinity for the NET and is approximately equipotent in inhibiting the uptake of dopamine and norepinephrine (Kuhar et al., 1999), noradrenergic mechanisms may have contributed to RTI-112-induced decreases in cocaine self-administration. However, GBR 12909 and RTI-177 differ in their selectivity for DAT over NET, yet they had a similar profile of behavioral effects and significant DAT occupancy at ED50 doses that decreased cocaine self-administration. Moreover, selective norepinephrine transporter inhibitors typically fail to reduce cocaine self-administration in nonhuman primates. Pretreatment with desipramine in rhesus monkeys trained under a second-order schedule had inconsistent effects (Mello et al., 1990). In another study, pretreatment with desipramine in rhesus monkeys trained under multiple fixed ratio schedules of cocaine and food delivery had no effect on cocaine self-administration (Kleven and Woolverton, 1993). Accordingly, it is more likely that the serotonergic effects of RTI-112 contributed to the interactions observed in the present study.
The results of neuroimaging studies with RTI-112 high-light the importance of conducting in vivo occupancy studies at behaviorally relevant drug doses. The potency of RTI-112 at DAT and SERT is nearly identical based on in vitro uptake assays. However, the ED50 dose of RTI-112 derived from behavioral studies exhibited a high level of SERT occupancy with no detectable DAT occupancy. This apparent in vivo selectivity for SERT over DAT may contribute significantly to the unique profile of behavioral effects observed for RTI-112 compared with GBR 12909 and RTI-177. Interestingly, the DAT occupancy dose-effect curves for all drugs were steep compared with in vitro binding and uptake assays, which generally encompass two log units. However, the behavioral dose-effect curves were also steep, consistent with the in vivo occupancy data observed in the present study. Reasons for the discrepancies often observed between in vitro and behavioral assays are unclear, but they likely involve a number of factors, including drug disposition, complex regulatory mechanisms, and multiple pharmacological targets.
Although the present study did not determine the selectivity of drug pretreatment effects on cocaine-maintained behavior, published studies with monoamine transporter inhibitors have yielded equivocal results. Pretreatment with the DAT inhibitor GBR 12909 selectively attenuated cocaine-maintained behavior compared with food-maintained behavior at a low unit dose of cocaine but not at a higher unit dose (Glowa et al., 1995). Similarly, administration of cocaine as a continuous infusion had selective effects on cocaine-maintained behavior compared with food-maintained behavior only at low unit doses of self-administered cocaine (Glowa and Fantegrossi, 1997). Several other studies with monoamine transporter inhibitors have demonstrated nonselective reductions in cocaine-maintained behavior and behavior maintained by alternative reinforcers, including food and stimulus-shock termination (Kleven and Woolverton, 1993; Glowa and Wojnicki, 1996; Howell et al., 2000). Whereas selectivity of effects on drug-maintained behavior is desirable, the nature of the alternative reinforcer and the behavior engendered are important considerations. The types of side effects that are viewed as tolerable should be evaluated in the context of effectiveness in reducing drug use.
A possible limitation to the use of DAT inhibitors as medications for treatment of cocaine addiction is their potential for abuse given their documented reinforcing effects (Howell and Wilcox, 2001). RTI-113 reliably maintained self-administration in squirrel monkeys (Howell et al., 2000) and rhesus monkeys (Wilcox et al., 2002). Similar results have been reported for other tropane analogs of cocaine (Bergman et al., 1989; Weed et al., 1995) and for GBR 12909 (Bergman et al., 1989; Howell and Byrd, 1991). In the present study, GBR 12909 and RTI-177 reliably maintained drug self-administration in all subjects. DAT occupancies at doses of GBR 12909 and RTI-177 that maintained peak rates of responding were 57 and 92%, respectively. Previous studies have reported levels of DAT occupancy between 65 and 76% for peak doses of cocaine and between 94 and 99% for peak doses of RTI-113 in rhesus monkeys (Wilcox et al., 2002). Similarly, studies in cocaine abusers found DAT occupancies between 60 and 77% for doses of cocaine within the range commonly used (Volkow et al., 1997). Interestingly, GBR 12909 and RTI-177 maintained lower rates of responding compared with cocaine across a broad range of doses, even though DAT occupancy was equal to or greater than that observed for cocaine. In behavioral studies in rodents and nonhuman primates, these compounds have slower onset and longer duration of action compared with cocaine (Howell et al., 2000; Kimmel et al., 2001). Hence, reinforcing effectiveness and the pattern of drug self-administration may be influenced by pharmacokinetics, in addition to steady-state levels of DAT occupancy. In addition, the mixed action transporter inhibitor RTI-112 did not function as a robust reinforcer in any subject. In addition to its pharmacokinetic profile of slow onset and long duration of action (Kimmel et al., 2001), its high affinity at other monoamine transporters may have limited its reinforcing effectiveness. It is important to acknowledge, however, that the present study focused on response rate measures maintained by a second-order schedule of drug delivery. There are multiple protocols available to measure the reinforcing effectiveness of drugs, and outcomes may vary depending on the methods used. For example, a recent study using a progressive-ratio schedule in rhesus monkeys failed to show a clear relationship between reinforcing effectiveness and drug time course of action (Lile et al., 2003). Accordingly, a more thorough evaluation of pharmacokinetics, selectivity, and reinforcing effectiveness is clearly warranted.
Direct comparisons were made between measures of DAT occupancy obtained for RTI-177 in the awake versus anesthetized states to investigate the potential influence of isoflurane anesthesia on DAT occupancy. It is well established that isoflurane can affect variables relevant to dopaminergic function that could influence DAT occupancy in anesthetized subjects (Adachi et al., 1999; Tsukada et al., 1999). However, within-subject comparisons in the present study demonstrated that DAT occupancy obtained during anesthesia did not differ statistically from occupancy obtained while awake. Although absolute quantification of DAT density may be sensitive to conditions of anesthesia, the relative measures reported in the present study were not influenced by isoflurane anesthesia. These results support the use of the generalized reference tissue method as a reliable means to measure relative occupancy in anesthetized subjects (Votaw et al., 2002).
In conclusion, the DAT inhibitors investigated were effective in reducing cocaine self-administration at doses that had no adverse behavioral effects. The reductions did not seem to result from an enhancement of the reinforcing effectiveness of cocaine because drug pretreatments were equally effective at both unit doses of cocaine, and cocaine intake was minimal at high pretreatment doses. For the drugs with selectivity for DAT over SERT, a high level of DAT occupancy was observed for doses that produced robust decreases in cocaine self-administration. In contrast, a mixed action inhibitor of DAT and SERT reduced cocaine self-administration at a high level of SERT occupancy with no detectable DAT occupancy. Collectively, the results indicate that high occupancy at DAT or SERT can induce significant decreases in cocaine self-administration. Regarding medications development to treat cocaine abuse, selective SERT inhibitors do not exhibit cocaine-like behavioral effects and do not seem appropriate as substitute agonist pharmacotherapies. For example, fluoxetine was ineffective for treatment of cocaine dependence in a placebo-controlled, double-blind trial (Grabowski et al., 1995). In contrast, selective DAT inhibitors clearly exhibit cocaine-like behavioral effects and seem promising as substitute agonist pharmacotherapies based on preclinical evaluations. However, their robust reinforcing effects in animal models indicate they may exhibit high abuse liability that could limit their clinical utility. The limited reinforcing effectiveness of the mixed action inhibitor in the present study is encouraging and suggests that dual actions at DAT and SERT could lead to compounds with modest cocaine-like properties appropriate for substitute agonist pharmacotherapies but with limited abuse liability. Overall, the results support the development of DAT inhibitors as potential pharmacotherapies in the treatment of cocaine abuse and indicate that mixed action inhibitors of DAT and SERT warrant consideration.
Acknowledgments
We gratefully acknowledge the technical assistance of Tango Howard, Olga Epstein, and Peggy Plant.
Footnotes
-
This work was supported in part by U.S. Public Health Service Grants DA10344 (to L.L.H.), DA00517 (to L.L.H.), DA06042 (to K.P.L.), DA06051 (to K.M.W.), DA05477 (to F.I.C.), and RR00165 (Division of Research Resources, National Institutes of Health). The Yerkes National Primate Research Center is fully accredited by the Association for the Assessment and Accreditation of Laboratory Animal Care International.
-
DOI: 10.1124/jpet.103.060293.
-
ABBREVIATIONS: DAT, dopamine transporter; PET, positron emission tomography; SERT, serotonin transporter; FECNT, 8-(2-[18F]fluoroethyl)-2β-carbomethoxy-3β-(4-chlorophenyl)nortropane; ZIENT, 2β-carbo[11C]methoxy-3β-(4′-((Z)-2-iodoethenyl)phenyl)nortropane; NET, norepinephrine transporter.
- Received September 18, 2003.
- Accepted February 19, 2004.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}