


Abstract
This study characterizes the sumatriptan-sensitive [5-hydroxytryptamine (5-HT)1B/1D] receptor in rabbit saphenous vein and basilar artery. (S)-(-)-1-{2-[4-(4-Methoxy-phenyl)-piperazin-1-yl]-ethyl}-isochroman-6-carboxylic acid methylamide (PNU-109291), a 5-HT1D subtype-selective agonist (human Ki = 2.5 ± 0.07 nM), did not contract either tissue, whereas o-methoxyphenylpiperazide derivative 4F (MPPA-4F), a 5-HT1B subtype-selective antagonist (human Ki = 4.6 ± 0.6 nM) potently inhibited sumatriptan-induced contraction in the saphenous vein and basilar artery. These results suggested that sumatriptan-induced contraction was mediated via the 5-HT1B receptor in these blood vessels. 5-HT1B receptor-mediated contraction was then compared in endothelium-intact and denuded vessels to evaluate the role of the endothelium in regulating sumatriptan-induced contractility in these tissues. The presence of an intact endothelium inhibited 5-HT1B-induced contraction in both tissues. Endothelial denudation or nitric-oxide synthase inhibition with Nω nitro-l-arginine methyl ester (l-NAME) (100 μM) increased the efficacy and potency of sumatriptan in the saphenous vein and basilar artery. Surprisingly, in endothelial-denuded vascular tissues, l-NAME (100 μM) also significantly increased the maximal 5-HT1B receptor-induced contraction in both tissues, with no effect on potency of sumatriptan. The effect of l-NAME after endothelial denudation may reflect the presence of a low density of residual endothelial cells as estimated by CD31 antibody staining combined with the modulating effect of nitric oxide released from nonendothelial cells in vascular tissue. Endothelial modulation was specific to 5-HT1B receptors because removal of the endothelium did not significantly alter contraction to norepinephrine, histamine, prostaglandin, or potassium chloride in the saphenous vein or basilar artery.
Sumatriptan is an antimigraine drug (Humphrey et al., 1988) with high affinities for vasocontractile serotonin 5-HT1B and 1D receptors (Peroutka and McCarthy, 1989). Cerebral vascular contraction is postulated, at least in part, to underlie the therapeutic efficacy of triptans (sumatriptan and zolmitriptan) in migraine headaches (Hamel, 1999). However, 5-HT1B/1D receptors are also widely distributed in the peripheral vasculature, raising the liability of coronary vasoconstriction, which has been associated with activation of 5-HT1B/1D receptors (MaassenVanDenbrink et al., 1998).
The vascular endothelium can critically modulate serotoninergic contractility via release of nitric oxide (Furchgott and Zawadzki, 1980; Valentin et al., 1996) and other endothelium-derived hyperpolarizing factors (McGuire et al., 2001). For example, in atherosclerotic animal models, sumatriptan produced significantly greater maximal contraction compared with controls (Ishida et al., 2001). Furthermore, the presence of endothelium was inhibitory to sumatriptan-induced vasoconstriction in vitro and in vivo (MacLean et al., 1994; Valentin et al., 1996; Carel et al., 2001). Thus, endothelial integrity is proposed to play a significant role in determining the therapeutic window of a 5-HT1B/1D receptor agonist such as sumatriptan.
The rabbit saphenous vein and basilar artery are convenient in vitro models for studying vascular contractility as surrogates for human peripheral and central vasculature (Cohen et al., 1997; Bhattacharya et al., 2003). Furthermore, mRNA for both 5-HT1B (high-density) and 5-HT1D (low-density) receptors are present in the saphenous vein (Wurch et al., 1997) and in cerebral vessels (Bouchelet et al., 1996), raising the possibility that both receptors may play a role in contractility in these tissues to varying degrees. The apparent affinity of GR127935 in the rabbit saphenous vein was in agreement with that reported for the human 5-HT1B receptor (Cohen and Schenck, 2000), although Valentin et al. (1996) suggested 5-HT1D to be the contractile receptor in a similar experiment with the same tissue. Furthermore, significant correlation existed between contractile potency of a series of triptans to their respective binding affinities at either 5-HT1B or 5-HT1D receptors, suggesting that either or both subtypes may modulate sumatriptan-induced contractility in saphenous vein (Cohen and Schenck, 1999). The rabbit basilar artery has not been characterized for the sumatriptan-sensitive receptor(s), although 5-HT1B and 5-HT1D receptors are thought to contribute to rabbit basilar artery contraction (Parsons et al., 1998). Interestingly, sumatriptan displayed distinct differences in amplification mechanisms between saphenous vein and basilar artery (Bhattacharya et al., 2003), indicating the possibility that different receptor subtypes may mediate contraction in the two tissues.
In this article, we 1) characterized the receptor activated by sumatriptan in rabbit saphenous vein and basilar artery using the selective 5-HT1B receptor antagonist MPPA-4F (Jorand-Lebrun et al., 1997) and the selective 5-HT1D receptor agonist PNU-109291 (Ennis et al., 1998); 2) studied the modulatory role of the endothelium by comparing sumatriptan-induced contraction in the presence and absence of the endothelium and in the presence of l-NAME, a nitric-oxide synthase inhibitor; and 3) addressed the specificity of endothelial modulation to 5-HT1B/1D receptor-induced contraction. Our results indicate that whereas 5-HT1D receptor activation failed to initiate vascular contraction, contraction in the saphenous vein and basilar artery was antagonized by selective blockade of 5-HT1B receptors. 5-HT1B receptor-induced vascular contraction was inhibited in the presence of the endothelium, and conversely, l-NAME increased the maximal response to sumatriptan, suggesting an important role of the endothelium in modulating contractility to sumatriptan in both vessels. Furthermore, l-NAME-sensitive factors regulated sumatriptan-induced contractility, even after endothelial removal, an effect that may reflect nonendothelial sources of NO and/or a residual very low density of endothelial cells. The effect of l-NAME and endothelium was specific to 5-HT1B receptors because a similar modulatory role did not occur with nonserotoninergic contractile agonists.
Materials and Methods
Receptor Binding. Serotonin receptor binding assays were performed and affinity constants were calculated for PNU-109291 and MPPA-4F using cells transfected with human 5-HT receptors as detailed in Zgombick et al. (1991) and Rasmussen et al. (2000).
Isolation of Rabbit Saphenous Vein and Basilar Artery. Male New Zealand White rabbits (2.0-3.0 kg) (Harlan, Indianapolis, IN) were sacrificed by injection of sodium pentobarbital (65-100 mg/kg) into the ear vein according to animal use protocols approved by the Lilly Animal Care and Use Committee. The saphenous vein and/or basilar artery was dissected free of connective tissue, cannulated with 99.9% platinum wire (32-gauge for saphenous vein; 36-gauge for basilar artery) in petri dishes containing Krebs' bicarbonate buffer (Bhattacharya et al., 2003). When appropriate, tissues were denuded of vascular endothelium by rotating the platinum wires along the luminal surface of the blood vessels. Endothelial integrity was determined by carbamylcholine (1 μM)-induced relaxation of PGF2α (3 μM)-induced contraction of the saphenous vein and histamine (1 μM)-induced contraction of the basilar artery.
Tissues were mounted in organ baths containing 10 ml of modified Krebs' solution of the following composition: 118.2 mM NaCl, 4.6 mM KCl, 1.6 mM CaCl2·2H2O, 1.2 mM KH2PO4, 1.2 mM MgSO4, 10.0 mM dextrose, and 24.8 mM NaHCO3. Tissue bath solutions were maintained at 37°C and aerated with 95% O2 and 5% CO2 (pH 7.4). An initial optimum force of 4 and 0.5 g was applied to the rabbit saphenous vein and basilar artery, respectively, resulting in baseline tensions of 1 and 0.25 to 0.30 g in the two tissues (Bhattacharya et al., 2003). Isometric contractions were recorded as changes in grams of force as measured with Sensotec transducers coupled to MP100 data acquisition software (BIOPAC Systems, Inc., Santa Barbara, CA).
Isolated Tissue Contractile Studies. Each tissue was initially challenged with KCl (67 mM) to confirm tissue viability. Cumulative concentration-response curves were generated to sumatriptan, PNU-109291 (5-HT1D agonist), or norepinephrine. No tissue was used more than once to generate a concentration-response curve. The nitric-oxide synthase inhibitor l-NAME, the 5-HT1B receptor antagonist MPPA-4F, or vehicle was incubated with the vessels for 30 min, followed by generation of the sumatriptan concentration-response curve.
Contractile Data Analysis and Material Sources. All results are expressed as mean ± S.E., where n represents the number of animals. Contractile data are expressed as a percentage of the response to a maximal contractile concentration of KCl (67 mM) administered initially in each tissue. EC50 values (-log EC50 or pEC50) were determined by a three-parameter logistic nonlinear model. Apparent estimates of MPPA-4F dissociation constant (pKB) were calculated for surmountable and insurmountable antagonism by the method of Arunlakshana and Schild (1959) and by Gaddum (1957), respectively. Statistical significance (P < 0.05) was determined by Student's t test using SigmaStat software.
Norepinephrine, histamine, PGF2α carbamylcholine, and l-NAME were purchased from Sigma-Aldrich (St. Louis, MO). Sumatriptan, PNU-109291, and MPPA-4F were provided by the Lilly Research Laboratories (Eli Lilly & Co., Indianapolis, IN). All compound solutions were made fresh daily before the start of each experiment.
Immunohistochemistry. Tissues were fixed overnight at 4°C in 4% neutral-buffered paraformaldehyde, followed by routine histological processing and sectioning onto charged microscope slides (Probe-On Plus; Fisher Scientific, Pittsburgh, PA). After deparaffinization, the slides were incubated in target retrieval solution (DAKO, Santa Barbara, CA) for 15 min at 95-99°C. Endothelial cell distribution and integrity was examined using the DAKO autostainer, incubating with biotinylated mouse anti-rat CD31 (BD Biosciences PharMingen, San Diego, CA) in a dilution of 1:100 at 4°C overnight. Negative controls were run for all sections using biotinylated isotype rat anti-mouse antibodies. Antibody binding was then localized using a peroxidase-labeled streptavidin-biotin complex (DAKO) followed by diaminobenzidine (DAKO) as the chromogen. Photomicrographs of tissue sections stained with CD31 were taken with a Leica DMR microscope equipped with a digital Spot camera. For analysis of percentage of endothelium (Fig. 7), the total length of discrete CD31-positive sections was measured and then calculated as a percentage of the total perimeter of the tissue. Percentage of relaxation was estimated by carbamylcholine-induced relaxation for every tissue and was assessed before tissue fixing for immunohistochemistry.

Representative picture of the rabbit saphenous vein immunolabeled with an antibody for the endothelial CD31 protein. CD31 stained the endothelium in an endothelium-intact saphenous vein (top), whereas endothelial denudation of saphenous vein resulted in small yet significant amount of punctuate labeling (middle). Correlation between endothelium and relaxation (bottom). Endothelial percentage was calculated by normalizing CD31 staining counts to tissue perimeter, whereas relaxation was assessed by carbamylcholine (1 μM)-induced loss of tone. The dashed line (bottom) depicted 95% predictive interval for the correlation plot.
Results
Binding Affinities of Sumatriptan, PNU-109291, and MPPA-4F. PNU-109291 and MPPA-4F (Fig. 1) were used to characterize whether the rabbit saphenous vein and the basilar artery expressed contractile 5-HT1B or 5-HT1D receptor subtypes. We measured the affinity of PNU-109291 and MPPA-4F in cell lines expressing human 5-HT receptor subtypes (Table 1). PNU-109291 displayed an affinity of 2.5 ± 0.07 nM for the h5-HT1D receptor. It was about 600-fold selective for this subtype compared with the h5-HT1A and h-5-HT2A receptors. Affinities of PNU-109291 for other 5-HT receptors, including 5-HT1B were greater than 10 μM. Thus, PNU-109291 was a highly selective ligand for the 5-HT1D receptor. Affinity values of MPPA-4F were 4.6 ± 0.6 and 62 ± 10.8 nM for h5-HT1B and h5-HT1D receptors, respectively (Table 1), demonstrating high affinity and selectivity for the 5-HT1B receptor.
Structures of C-[3-(2-dimethylamino-ethyl)-1H-indol-5-yl]-N-methyl-methanesulfonamide (sumatriptan), MPPA-4F, and PNU-109291.
Binding affinities (Ki) MPPA-4F and PNU-109291 at h5-HT receptors
Data are the mean ± standard error from three to five independent experiments.
Effect of PNU-109291 and MPPA-4F in the Rabbit Saphenous Vein and Basilar Artery. Endothelial denuded rabbit saphenous vein and basilar artery were challenged with sumatriptan or the high-affinity 5-HT1D receptor agonist PNU-109291 (Fig. 2). Whereas sumatriptan induced vascular contractility with potency similar to that reported previously (Bhattacharya et al., 2003), PNU-109291 failed to elicit significant contraction, in concentrations as high as 100 μM. Tissues that did not contract to PNU-109291 were viable as assessed by their contraction to KCl (67 mM) (data not shown). Thus, the rabbit saphenous vein and the basilar artery did not functionally contract to 5-HT1D receptor activation.

Effect of sumatriptan and PNU-109291 on rabbit saphenous vein and basilar artery. Increasing concentrations of sumatriptan resulted in contraction of the saphenous vein (top) and basilar artery (bottom). PNU-109291 did not elicit any significant contraction. Contractile effects were measured as percentage of KCl (67 mM)-induced maximal force. Points are mean values and vertical lines are standard error of means for appropriate number of animals (in parentheses).
In the rabbit saphenous vein, MPPA-4F concentration dependently attenuated sumatriptan-induced contractility (Fig. 3, top) without suppression of the maximal response, suggesting surmountable antagonism of sumatriptan-induced contraction by MPPA-4F. MPPA-4F possessed high affinity for the 5-HT1B receptor in the rabbit saphenous vein, with a dissociation constant of approximately 20 nM. In contrast, although MPPA-4F potently antagonized sumatriptan contractility in the basilar artery, it produced apparent insurmountable antagonism of sumatriptan-induced contraction with an antagonist dissociation constant of 2.5 nM (Fig. 3, bottom). These result strongly suggested that the contraction in the saphenous vein and basilar artery was mediated by the 5-HT1B receptor, although differences may exist in MPPA-4F-mediated antagonism of sumatriptan-induced contraction.
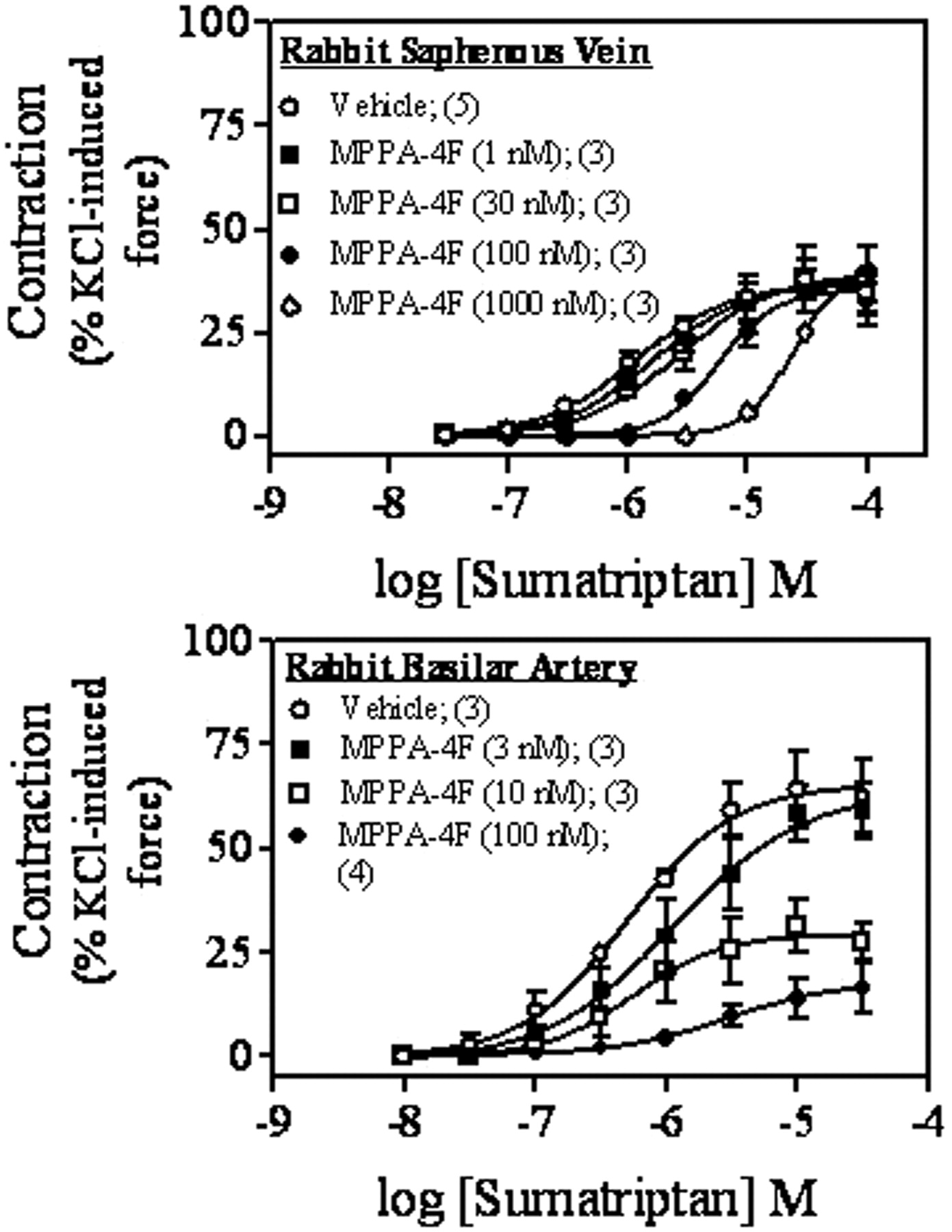
Effect of MPPA-4F on sumatriptan-induced vasoconstriction. MPPA-4F attenuated sumatriptan-induced contraction of the rabbit saphenous vein (top) and the basilar artery (bottom). Contractile effects were measured as percentage of KCl (67 mM)-induced maximal force. Points are mean values and vertical lines are standard error of means for appropriate number of animals (in parentheses).
Effect of Endothelium on the 5-HT1B Receptor-Mediated Vasocontractility. To assess the role of endothelial mediators in modulating 5-HT1B receptor contractility, sumatriptan-induced contraction was compared in the presence or absence of functional endothelium (Fig. 4, top). Endothelial integrity was assessed by the ability or inability of carbamylcholine (1 μM) to relax vascular tissues (Fig. 4, bottom). Sumatriptan-induced contraction of both saphenous vein and basilar artery was inhibited in the presence of an intact endothelium. Interestingly, endothelial integrity modulated the 5-HT1B response more in the basilar artery than in the saphenous vein. For example, basilar arteries with endothelium only modestly contracted to 10 μM sumatriptan, whereas similar concentrations of sumatriptan produced about 65 to 70% contractility when the endothelium was removed (Fig. 4, top). On the other hand, contraction to 10 μM sumatriptan in the saphenous vein was only increased to approximately 30% contractility when the endothelium was removed (Fig. 4, top). In the saphenous vein, endothelial denudation also increased the potency (pEC50) of sumatriptan from 4.9 ± 0.6 to 5.7 ± 0.1. These data indicate that endothelial factor(s) play an important role in attenuating sumatriptan-induced contractile signal amplification.

Effect of vascular endothelium on sumatriptan-induced contraction. Presence of a functional endothelium (+ endothelium) was inhibitory to sumatriptan-induced contraction in both saphenous vein and basilar artery (top). Functional presence or absence of endothelium in vascular tissues as assessed by the ability of carbamylcholine (1 μM) to relax PGF2α (3 μM)-contracted saphenous vein or histamine (1 μM)-contracted basilar artery (bottom). Points are mean values and vertical lines are standard error of means for appropriate number of animals (in parentheses). *, P < 0.05 (t test) for tissues with and without endothelium.
Effect of l-NAME on Sumatriptan-Induced Vascular Contraction. To further characterize the role of endothelium, we used l-NAME (100 μM) in endothelium-intact rabbit saphenous vein and basilar artery (Fig. 5). In the presence of l-NAME, sumatriptan increased the maximal contraction in both tissues (Table 2). Interestingly, l-NAME increased the potency (pEC50) of sumatriptan in the saphenous vein from 4.9 ± 0.4 to 5.8 ± 0.2 (Table 2), identical to the effect observed with endothelial denudation (Table 2; Fig. 4). However, in both the saphenous vein and basilar artery, l-NAME increased the maximal contraction to a greater extent than simple removal of the endothelium (compare responses from Figs. 4 and 5). These results indicated that nitric oxide (NO) release inhibited 5-HT1B receptor-mediated contraction in both tissues.

Effect of l-NAME on endothelium-intact rabbit saphenous vein and basilar artery. Sumatriptan-induced contraction of the saphenous vein (top) or the basilar artery (bottom) in the presence of endothelium and l-NAME (100 μM). Points are mean values and vertical lines are standard error of means for appropriate number of animals (in parentheses). *, P < 0.05 (t test) for tissues with and without l-NAME.
Pharmacology of sumatriptan in saphenous vein and basilar artery, with and without endothelium
Data are mean values with standard error of means from three to five independent experiments.
In endothelium denuded vascular rings, l-NAME (100 μM) produced significant increases in the maximal sumatriptan contraction, suggesting nonendothelial l-NAME-sensitive factor(s) as inhibitors of 5-HT1B receptor-mediated vascular contractility (Fig. 6). In endothelium-denuded tissues, l-NAME did not cause significant increases in the potency of sumatriptan (Table 2). The degree to which l-NAME affected the efficacy of sumatriptan was very similar in endothelium-denuded saphenous vein and basilar artery. In the denuded saphenous vein, l-NAME increased the maximum response to sumatriptan from 38.8 ± 3.2 to 69.2 ± 5.5% contractility. Likewise, in the denuded basilar artery, efficacy of sumatriptan increased from 57.8 ± 4.7 to 109.5 ± 8.2% contractility

Effect of l-NAME on endothelium-denuded rabbit saphenous vein and basilar artery. Sumatriptan-induced contraction of the saphenous vein (top) or the basilar artery (bottom) in the absence of functional endothelium but in the presence of l-NAME (100 μM). Points are mean values and vertical lines are standard error of means for appropriate number of animals (in parentheses). *, P < 0.05 (t test) for tissues with and without l-NAME.
Anti-CD31 Antibody Staining as an Estimate of Endothelial Integrity. The pronounced effect of l-NAME after endothelial removal (assessed by the inability of carbamylcholine to induce relaxation) raised the possibility that remaining endothelial cells may release l-NAME-sensitive factors that inhibited tissue contractility. Representative endothelial staining with anti-CD31 antibody in saphenous vein, both with intact endothelium (top) and after endothelium denudation (middle) demonstrated modest residual CD31 staining in endothelial-denuded tissue (Fig. 7). Thus, loss of relaxation as estimated by loss of cholinergic relaxation may not necessarily translate to a complete absence of endothelium. This was further confirmed by a positive correlation between relaxation to carbamylcholine and the density of endothelium as measured by the intensity of CD31 staining (Fig. 7, bottom). The data indicated that tissues that do not significantly relax to carbamylcholine may still have residual endothelium.
Effect of the Endothelium and l-NAME on Nonserotonergic Contractile Agonists. We next examined the specificity of endothelium/l-NAME-dependent effects by using norepinephrine as the contractile agonist in rabbit saphenous vein (Fig. 8). Neither l-NAME (100 μM) nor endothelial removal had significant effects on norepinephrine-induced contraction, suggesting that relaxant or inhibitory factor(s) is modulated by the 5-HT1B receptor system specifically in these tissues. Likewise, as shown in Table 3, the presence of an intact endothelium did not have significant effects on the amplitude of contraction to KCl (67 mM), histamine (1 μM), or PGF2α (3 μM).

Effect of norepinephrine on contraction of the rabbit saphenous vein both in the presence or absence of endothelium or l-NAME. Contractile effects were measured as percentage of KCl (67 mM)-induced maximal force. Points are mean values and vertical lines are standard error of means for appropriate number of animals (in parentheses).
Effect of potassium chloride, histamine, prostaglandin, and carbamylcholine on saphenous vein and basilar artery
Data are mean values with standard error of means from three to eight independent experiments.
Discussion
Sumatriptan contracted the rabbit saphenous vein and basilar artery and had similar high affinity (Ki) for the h5-HT1B (7.9 nM), h5-HT1D (5.0 nM), and h5-HT1F (25.1 nM) receptors (Bard et al., 1996; Cohen and Schenck, 1999), suggesting that sumatriptan-induced vascular contraction could be mediated via activation of 5-HT1B, 5-HT1D, and/or 5-HT1F receptors. In this study, we characterized the sumatriptan-sensitive receptor in the rabbit saphenous vein and basilar artery as the 5-HT1B receptor based primarily on the pharmacology of PNU-109291 and MPPA-4F.
PNU-109291 displayed high affinity (2.5 ± 0.07 nM) and selectivity (>4000-fold) as an agonist (Ennis et al., 1998) for the h5-HT1D receptor subtype compared with h5-HT1B and h5-HT1F receptors (Table 1). The inability of PNU-109291 to contract the saphenous vein or basilar artery (Fig. 2) strongly suggested that sumatriptan-induced vascular contractility was mediated by either 5-HT1B or 5-HT1F receptors, but not the 5-HT1D receptor subtype. This observation was in agreement with the inability of PNU-109291 to contract bovine and human cerebral arteries (Bouchelet et al., 2000).
The possibility that 5-HT1B receptors mediated serotonergic contractility was tested in the saphenous vein and basilar artery by using MPPA-4F, a 5-HT1B receptor-selective antagonist, to inhibit sumatriptan-induced contraction (Fig. 3). MPPA-4F significantly inhibited vascular contractions to sumatriptan. MPPA-4F did not elicit contraction by itself, ruling out the possibility that the attenuation of contraction to sumatriptan was due to desensitization of sumatriptan-sensitive receptors. In the rabbit saphenous vein, MPPA-4F displayed surmountable antagonism with an antagonist dissociation constant of 20 nM. In the basilar artery, MPPA-4F displayed insurmountable antagonism with an antagonist dissociation constant of 2.5 nM. Although the dissociation constants for MPPA-4F at rabbit serotonin receptors are not known, the dissociation constants at the h5-HT1B and h5-HT1D receptors were 4.6 ± 0.6 and 62 ± 10.8 nM, respectively (Table 1). Thus, based on the affinities of MPPA-4F, the 5-HT receptor subtype mediating vascular contraction in both the saphenous vein and basilar artery closely resembled the 5-HT1B receptor. At this time, we do not fully understand the difference in the nature of antagonism by MPPA-4F between the saphenous vein and basilar artery. It is possible that slow dissociation of MPPA-4F from the 5-HT1B receptors in the basilar artery, but not in the saphenous vein, caused insurmountability. It is notable that GR127935, another 5-HT1B/1D antagonist, caused insurmountable antagonism in the dog basilar artery (Skingle et al., 1996), yet the antagonism was surmountable in the rabbit saphenous vein (Valentin et al., 1996; Cohen and Schenck, 2000). It is plausible that cerebral 5-HT1B receptors bind MPPA-4F and GR127935 at a noncompetitive site, distinct from the competitive site (with respect to sumatriptan) in the saphenous vein. Another possibility is that the coupling mechanism of 5-HT1B receptors in the rabbit saphenous vein and basilar artery is different. Subtle differences in the amplification of sumatriptan-induced contraction also occurred between the two tissues (Bhattacharya et al., 2003).
The 5-HT1F receptor was ruled out as a mediator of vasoconstriction because 1) PNU-109291 did not contract the vascular tissues even in high concentrations of 100 μM; 2) in these blood vessels, the affinity estimates for MPPA-4F, a 5-HT1B receptor-selective antagonist (Jorand-Lebrun et al., 1997), did not match its binding affinity to the 5-HT1F receptor (Table 1; Fig. 3); and 3) Cohen and Schenck (1999) failed to elicit saphenous venoconstriction with a high-affinity 5-HT1F receptor agonist. Therefore, sumatriptan-induced vascular contraction was not mediated by the 5-HT1F receptor in either the peripheral or brain blood vessels.
The influence of a functional endothelium that modulated 5-HT1B receptor-induced contraction was studied by comparing responses in endothelium-intact and -denuded saphenous vein and basilar artery as assessed by cholinergic relaxation (Fig. 4, bottom). Endothelial denudation increased the potency and maximal response to sumatriptan (Fig. 4; Table 2), suggesting that a functional endothelium inhibited sumatriptan-induced contractility in both the saphenous vein and basilar artery. Furthermore, l-NAME, which inhibits NO formation, increased the potency and maximal response to sumatriptan in endothelium-intact tissues (Fig. 5), and the effect of l-NAME to increase the maximal contraction was greater than the effect of endothelial denudation in both vessels. The potency of sumatriptan in the presence of l-NAME or in the absence of endothelium in the rabbit saphenous vein and in the basilar artery (Table 2) was in agreement with previous data (Cohen and Schenck, 2000; Bhattacharya et al., 2003). Interestingly, in the absence of the endothelium or in the presence of l-NAME, sumatriptan was more potent and produced a greater maximal contraction in the basilar artery than in saphenous vein. Likewise, the potency (pEC50) of sumatriptan was greater in dog basilar artery (6.8 ± 0.06) than in the dog saphenous vein (6.1 ± 0.13) (Gupta et al., 1999). This is consistent with either 1) higher reserve for 5-HT1B receptors in the cerebral artery than in the saphenous vein or 2) differences in receptor coupling between the two tissues as observed previously (Bhattacharya et al., 2003).
In the absence of a functional endothelium (as assessed by carbamylcholine-induced relaxation), l-NAME further increased the efficacy of sumatriptan with no effect on its potency in both tissues (Table 2; Fig. 6). This novel observation raised the possibility that nonendothelial NO and/or a low density of residual endothelial cells could release sufficient NO to markedly alter contractility to sumatriptan. This latter possibility is supported by the fact that vascular smooth muscle cells expressed functional nitric-oxide synthase (Boulanger et al., 1998; Hecker et al., 1999). Thus, sumatriptan-induced vascular contractility may be modulated by both endothelial and nonendothelial sources of NO. In addition, the potentiating effect of l-NAME (Fig. 6) after apparent endothelial removal could be due to endothelial NO released from residual endothelial cells that remained after endothelial denudation. Preliminary data also support this possibility because tissues that did not relax to cholinergic stimulation (and by that measure lacked functional endothelium) expressed endothelial markers, although at low levels (Fig. 7). The possibility that sumatriptan may be activating release of NO from residual endothelial cells as well as from nonendothelial cells is consistent with the greater increase in maximal vascular contractility to sumatriptan with the combination of endothelial removal and l-NAME than with either intervention alone. Regardless, it is worth noting that the modulatory role of NO seems to be specific to the activation of 5-HT1B receptor, because the potency and efficacy of norepinephrine remained unchanged after endothelial denudation or in the presence of l-NAME (Fig. 8). Likewise, KCl, histamine, or prostaglandin-induced contraction of the basilar artery and saphenous vein was unaltered after endothelial denudation (Table 3).
Because the rabbit 5-HT1B receptor is >90% identical to the human counterpart (Bard et al., 1996), one might expect that under conditions of compromised vascular integrity such as atherosclerosis (Ishida et al., 2001), both peripheral and cerebrovascular contractility to sumatriptan, and possibly other triptans, may be enhanced. This study raises the importance of sumatriptan's ability to affect the release of NO from endothelial and nonendothelial vascular sources as a mechanism to modulate vascular contractility and hence, efficacy and toxicity of this antimigraine agent.
Footnotes
-
DOI: 10.1124/jpet.103.062653.
-
ABBREVIATIONS: 5-HT, 5-hydroxytryptamine; MPPA-4F, o-methoxyphenylpiperazide derivative 4F; PNU-109291, (S)-(-)-1-{2-[4-(4-methoxyphenyl)-piperazin-1-yl]-ethyl}-isochroman-6-carboxylic acid methylamide; l-NAME, Nω-nitro-l-arginine methyl ester; PGF2α, prostaglandin F2α; h5-HT, human 5-hydroxytryptamine; NO, nitric oxide; GR127935, N-[methoxy-3-(4-methyl-1-piperazinyl)phenyl]-2′-methyl-4′-(5-methyl-1,2,4-oxadiazol-3-yl)[1,1-biphenyl]-4-carboxamide.
- Received December 1, 2003.
- Accepted December 31, 2003.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}