


Abstract
Our goal was to characterize the 5-hydroxytryptamine (5-HT) receptor(s) mediating contraction in the isolated right renal artery, testing the hypothesis that the 5-HT2A receptor would be the primary and likely only 5-HT receptor involved in contraction. Contraction of arteries was investigated in isolated tissue baths, and expression of 5-HT receptors was measured using immunohistochemical and Western analyses. Compared with endothelium-denuded rat aorta, a tissue with an established 5-HT2A receptor, endothelium-denuded renal artery contracted to 5-HT with a 10-fold greater potency. Surprisingly, the 5-HT2B receptor agonist α-methyl-5-(2-thienylmethoxy)-1H-indole-3-ethanamine hydrochloride (BW723C86) caused a concentration-dependent contraction that was antagonized by the 5-HT2B receptor antagonist 6-methyl-1,2,3,4-tetrahydro-1-[3,4-dimethoxyphenyl) methyl-9H-pyrido[3,4b]indole] hydrochloride (LY272015) and nonselective 5-HT2 receptor antagonist 6-methyl-1-(1-methylethyl)-ergoline-8b-carboxylic acid 2-hydroxy-1-methylpropyl ester maleate (LY53857). Correlation of -log EC50 values with binding affinities (pKi) indicated that contraction of the renal artery elicited by 13 different agonists was likely consistent with activation of a 5-HT2A (r = 0.928) and 5-HT2B (r = 0.843) receptor. 5-HT-induced contraction was shifted by the 5-HT2A receptor antagonist ketanserin (3 and 10 nM) and the 5-HT2B receptor antagonist LY272015 (10 and 50 nM). Higher than expected concentrations of the 5-HT2A/5-HT2B receptor antagonist LY53857 were needed to antagonize 5-HT-induced contraction and the 5-HT2B receptor antagonist 2-amino-4-(4-fluoronaphth-1-yl)-6-isopropylpyrimidine (RS127445) was virtually inactive. Western and immunohistochemical analyses of the renal artery validated the presence of 5-HT2A and 5-HT2B receptor protein. These results suggest that the renal artery possesses a complex 5-HT receptor population, including ketanserin- and LY272015-sensitive receptors. This unique pharmacology may reflect differences in 5-HT receptor coupling between tissues or heterogeneity in the subtype(s) of 5-HT receptors expressed in the renal artery.
5-HT has an interesting history of involvement in modifying arterial tone, a parameter relevant to blood pressure (Turla and Webb, 1989; Martin, 1994). We have recently begun investigation of the renal artery because the kidney is unarguably relevant to determination of blood pressure. 5-HT has been suggested to be made within the kidney (Hafdi et al., 1996) and localized to cells of the kidney (Lincoln et al., 1990). In the isolated rat renal artery, it is established that 5-HT causes a concentration-dependent contraction, but the receptor mechanism by which this occurs is unknown and may be species-dependent.
In the isolated main renal artery of the rabbit, 5-HT causes contraction through activation of a silent 5-HT1B/1D receptor (Choppin and O'Connor, 1994; Hill et al., 2000) and causes conduit renal arteries in the conscious rabbit to spasm (Wright and Angus, 1987). In the dog, 5-HT reduces renal blood flow via activation of a 5-HT2-like receptor (Blackshear et al., 1991; Takahashi et al., 1992). In human renal arteries, 5-HT causes a contraction associated with marked oscillations (Ueda et al., 1982). In many but not all isolated rat arteries, it is the 5-HT2A receptor that is primarily responsible for 5-HT-induced contraction, evidenced by the effectiveness of agonists such as DOI and α-methyl-5-HT, and antagonism of contraction by ketanserin and MDL 100907. Many of these studies were performed before development of drugs that were selective for the 5-HT2B receptor; thus, the potential involvement of this receptor in renal arterial contraction is unknown. This issue is of particular interest because evidence supports that the 5-HT2B receptor, although present in arterial smooth muscle, does not mediate 5-HT-induced contraction under normal, nonhypertensive conditions (Watts, 1997; Watts and Fink, 1999; Banes and Watts, 2002; Russell et al., 2002).
We hypothesized that the 5-HT2B receptor would not be involved in mediating contraction to 5-HT in the renal artery from the normal rat and that, as has been observed in other rat arteries, the 5-HT2A receptor was primarily responsible for mediating 5-HT-induced contraction. We present here studies that potentially negate this hypothesis. Although we have observed that the 5-HT2A receptor certainly plays a major role in 5-HT-induced contraction, the renal artery possesses what seems to be a functioning 5-HT2B-like receptor in the normal rat, or a 5-HT receptor complement with significant differences in receptor/effector coupling compared with other arteries. This study serves to demonstrate two ideas. First, that the pharmacology of the 5-HT receptor(s) mediating contraction in the isolated renal artery is complex and second that the 5-HT2B receptor, or similar receptor, may be differently coupled in normal tissues.
Materials and Methods
Animal Use. Male Sprague-Dawley rats (0.225–0.250 kg; Charles River Breeding Laboratories, Portage, MI) were used. All rats were given free access to standard pelleted rat chow (Harlan/Teklad 8640 Rodent Diet).
Blood Pressure Measurement. Systolic blood pressures of rats were determined in the conscious state by the tail cuff method (pneumatic transducer; Narco, Austin, TX).
Isolated Smooth Muscle Contractility Measurement. Rats were deeply anesthetized with pentobarbital (60 mg kg-1 i.p.) to the point of a loss of eyelid reflex and lack of withdrawal from painful stimuli. Aorta, right renal artery, and stomach fundus were placed in physiological salt solution consisting of 130 mM NaCl, 4.7 mM KCl, 1.18 mM KH2PO4, 1.17 mM MgSO4·7H2O, 1.6 mM CaCl2·2H2O, 14.9 mM NaHCO3, 5.5 mM dextrose, and 0.03 mM CaNa2EDTA, pH 7.2. Aorta and renal artery were cleaned of fat and connective tissue, cut into helical strips, and rubbed with a moistened cotton swab to remove the endothelial cell layer. Two strips from the inner most section of the stomach fundus were dissected. Strips were mounted as longitudinal preparations and placed on stainless steel holders in tissue baths (30 and 50 ml) for isometric force recordings using Grass polygraphs and transducers (Astro-Med, West Warwick, RI) or PowerLab for the Macintosh (ADInstruments, Dover, NH). Tissues were placed under optimum resting force (1500 mg for aorta, 500 mg for renal artery, 4000 mg for stomach fundus; determined in preliminary experiments). Muscle baths were filled with warmed (37°C), aerated (95% O2, 5% CO2) physiological salt solution. Tissues equilibrated for 1 h with frequent exchanges of warmed salt solution. After this hour, arteries were challenged with a maximal concentration of α-adrenergic receptor agonist phenylephrine (10 μM) and stomach fundus with KCl (67 mM). Phenylephrine and KCl were washed out, and tissue tone was returned to baseline. In arterial tissue, functional integrity of the endothelial cells was evaluated by testing relaxation caused by acetylcholine (1 μM) in strips contracted with a half-maximal concentration of phenylephrine (10–100 nM). In all experiments, the endothelial cell layer was functionally removed as evidenced by the lack of relaxation (5 ± 3% relaxation) caused by acetylcholine. One of the following protocols was then followed.
Use of 5-HT Receptor Agonists. In each tissue, two 5-HT receptor agonists (10-9–10-4 M) were used to generate cumulative concentration response curves in isolated arteries. These agonists were randomized and separated by a wash period of 1 h, with a minimum of 10 buffer exchanges, with the exception of BW723C86 and α-methyl-5-HT. Both these agonists were difficult to wash out, so when they were administered as the first agonist, the tissues were not used to generate a second agonist curve. However, there were tissues in which the response to BW723C86 and α-methyl-5-HT was generated as a second agonist. In comparing responses of first and second agonists, we could not detect a difference in potency or maximum response to individual agonists tested, and thus these data are pooled.
Use of 5-HT Receptor Antagonists. Tissues were exposed to either the appropriate vehicle (water or 0.1% dimethyl sulfoxide) or antagonist and incubated for 1 h without washing. A cumulative concentration response curve to 5-HT or BW723C86 (10-9–10-4 M) was then performed.
Western Analysis. Tissues were isolated directly from the animal, cleaned, and placed directly into liquid nitrogen. In liquid nitrogen, tissues were ground to a powder, and ice-cold homogenization buffer was added [125 mM Tris (pH 6.8), 4% SDS, 20% glycerol, 0.5 mM phenylmethylsulfonyl fluoride, 1 mM orthovanadate, 10 μg/ml aprotinin, 10 μg/ml leupeptin]. Homogenates were vortexed, sonicated briefly and transferred to a plastic centrifuge tube and spun at 4°C to pellet debris. Supernatant was separated from the pellet and analyzed for protein concentration (BCA protein kit; Sigma-Aldrich, St. Louis, MO). Fifty micrograms of total protein was heated at 100°C for 5 min with standard 4:1 sample buffer. Proteins were separated on 1 mm-thick 10% SDS polyacrylamide gels using a Mini Bio-Rad III apparatus. Membranes were blocked for 3 h in 4% chick egg ovalbulmin [4°C, Tris-buffered saline (TBS)/0.1% Tween + 0.025% NaN3]. Primary antibody (0.5 μg/ml, mouse antibody for 5-HT2B receptor; BD PharMingen, San Diego, CA) was incubated with blots overnight at 4°C. Blots were then rinsed thrice in TBS + 0.1% Tween with a final rinse in TBS and incubated with horseradish peroxidase-linked anti-mouse secondary antibody (1:10,000; Amersham Biosciences Inc., Piscataway, NJ) for 1 h at 4°C with rocking. ECL reagents (Amersham Biosciences Inc.) were used to visualize bands. Gels were stained with Gel Code Blue (Pierce Chemical, Rockford, IL) to verify protein loading, and blots were reprobed with smooth muscle α-actin primary antibody (1:1000; Oncogene Research Products, Boston, MA) to ensure equal protein loading.
Immunohistochemical Analysis. Renal arteries were snap frozen in OCT compound and stored at -70°C until use. Arterial sections (8 μm) were cut and air-dried (overnight, room temperature). Samples were cold acetone fixed, washed three times with phosphate-buffered saline, and endogenous peroxidase blocked [0.3% H2O2 in phosphate-buffered saline (PBS) for 30 min]. Sections were blocked for nonspecific binding for 2 h in PBS containing 1.5% of competing serum. In a humidified chamber, samples then incubated 24 h with antibody (5-HT2A receptor; Santa Cruz Biotechnology, Inc., San Diego, CA; 4°C, 5 μg/ml with 1.5% blocking serum in PBS) or antibody neutralized with 5-fold excess of competing peptide. The remaining steps were carried out in accordance to the manufacturer's instructions (Vector Laboratories, Burlingame, CA). Sections were washed three times with PBS and incubated with a peroxidase-conjugated secondary antibody (30 min, room temperature). Samples were washed and incubated with Vectastain ABC Elite reagent (30 min, room temperature) followed by 3,3′-diaminobenzidine/H2O2. Reaction was stopped with washing, and sections were air-dried, hematoxylin-stained, mounted, and photographed using a Spot 2 digital camera on a Leica light microscope with filters.
Materials. Drugs were made daily and in water unless otherwise specified. 5-HT creatinine sulfate, BW723C86, α-methyl-5-HT, mCPP, (±)8-OH-DPAT, 2-methyl-5-HT, 5-methoxytryptamine, (±)-DOI, 5-carboxamidotryptamine, phenylephrine hydrochloride, acetylcholine chloride, ketanserin tartrate (dimethyl sulfoxide), and LY53857 were purchased from Sigma-Aldrich. LY344864 was a gift from Eli Lilly & Co. (Indianapolis, IN), CP93129 was from Pfizer (Groton, CT), (+)-norfenfluramine was from SRI International (Menlo Park, CA), sumatriptan was from GlaxoSmithKline (Stevenage, Hertfordshire, UK), and RS127445 (dimethyl sulfoxide) was from Roche (San Francisco, CA). The 5-HT2B receptor antibody was obtained from BD PharMingen, whereas 5-HT2A receptor antibody and competing peptide was purchased from Santa Cruz Biotechnology, Inc. Vector kits (Vector Laboratories) were used for immunohistochemical analysis.
Data Analysis. Data are presented as means ± standard error of the mean for the number of animals. Contraction is reported as a percentage of response to phenylephrine (10 μM) or KCl (67 mM KCl). -log EC50 values (negative logarithm of the agonist concentration necessary to produce a half-maximal response [M]) were determined using nonlinear regression analysis in GraphPad Prism (GraphPad Software Inc., San Diego, CA). Apparent antagonist dissociation constants (apparent KB values, reported as pKB) were calculated using the method of Arunlakshana and Schild (1959). When comparing two groups, an unpaired Student's t test was used or, in concentration-response curves, analysis of variance with repeated measures. Analyis of variance followed by Student-Newman-Keuls post hoc test was performed when comparing three or more groups. In all cases, a p value less than or equal to 0.05 was considered statistically significant and is indicated with an asterisk (*). Band density was quantified using the public domain program NIH Image (version 1.62).
Results
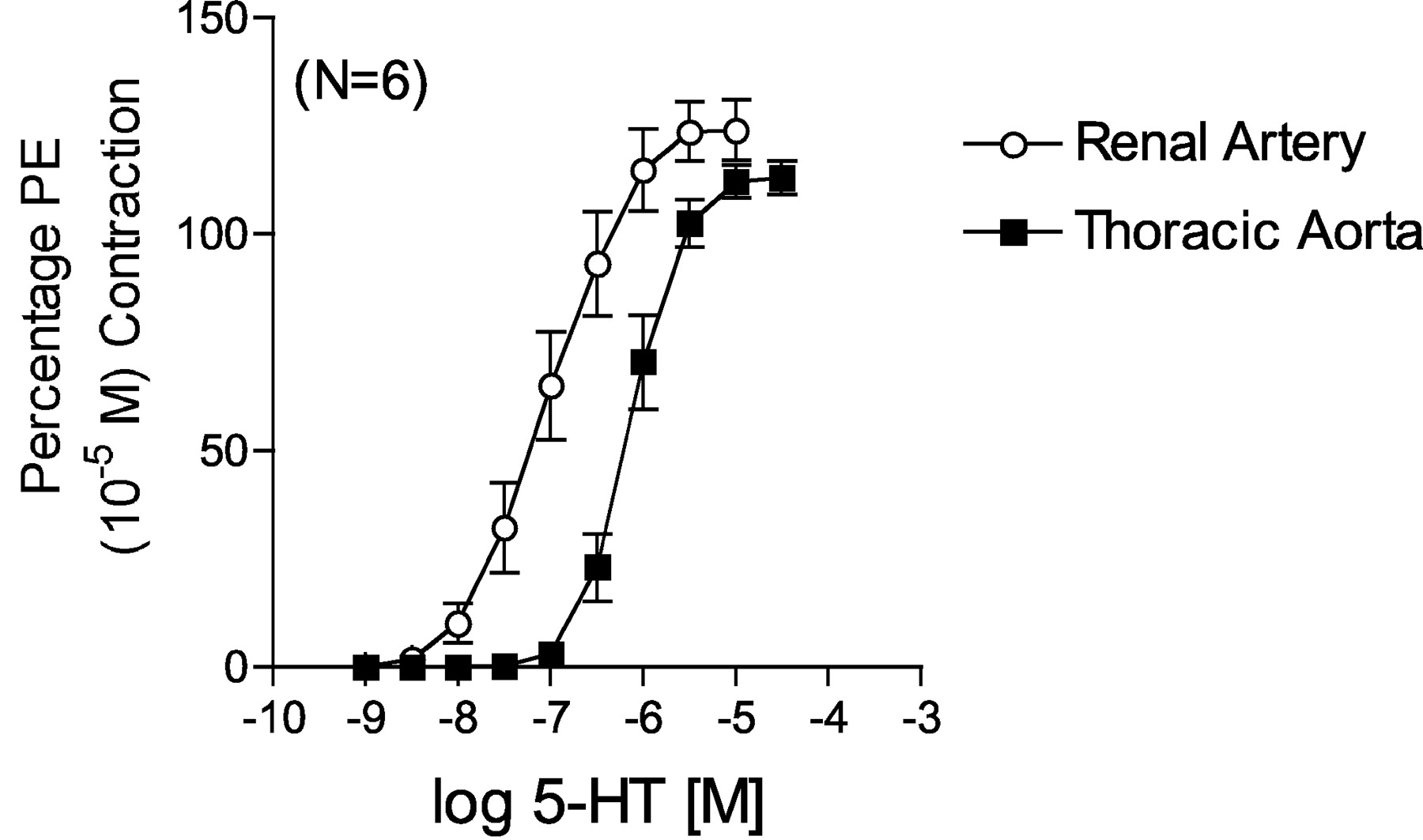
Comparison of 5-HT-Induced Contraction in Rat Aorta and Right Renal Artery.Figure 1 demonstrates a difference between 5-HT-induced contraction in the thoracic aorta and main right renal artery dissected from the same normotensive Sprague-Dawley rats (systolic blood pressure of 120 ± 2 mm Hg). The rat thoracic aorta is an established model of 5-HT2A receptor-mediated contraction, as evidenced by the potency of 5-HT and the repeated demonstration that antagonists that block contraction have a high affinity for the 5-HT2A receptor (for review, see Martin, 1994). 5-HT possessed a 10-fold greater potency in contracting the renal artery (–log EC50 value [M] = 7.03 ± 0.02) compared with aorta (6.08 ± 0.08).
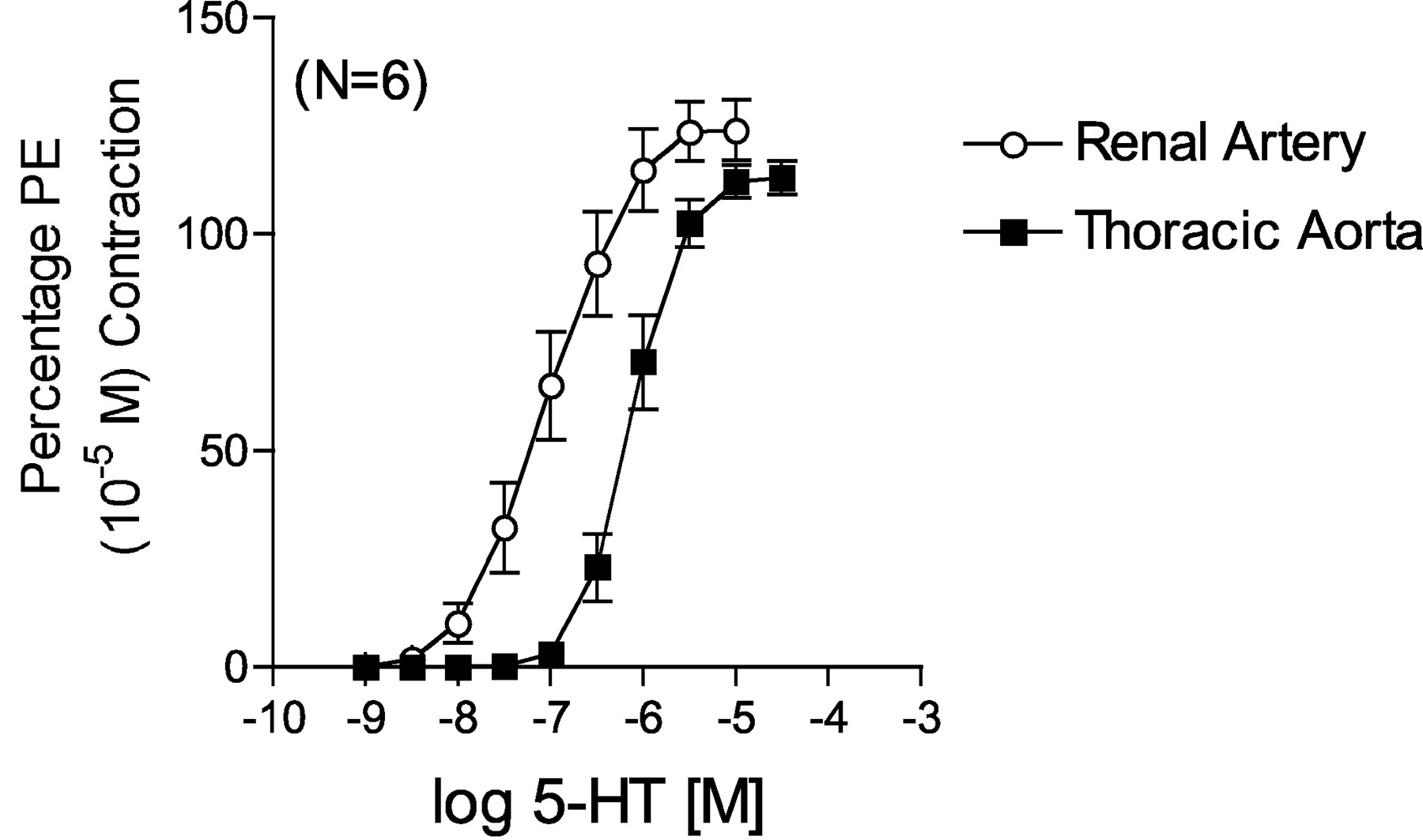
Comparison of 5-HT-induced contraction in endothelium-denuded aorta and right renal artery from the normotensive rat. Points represent means ± S.E.M. for the number of animals in parentheses. PE, phenylephrine.
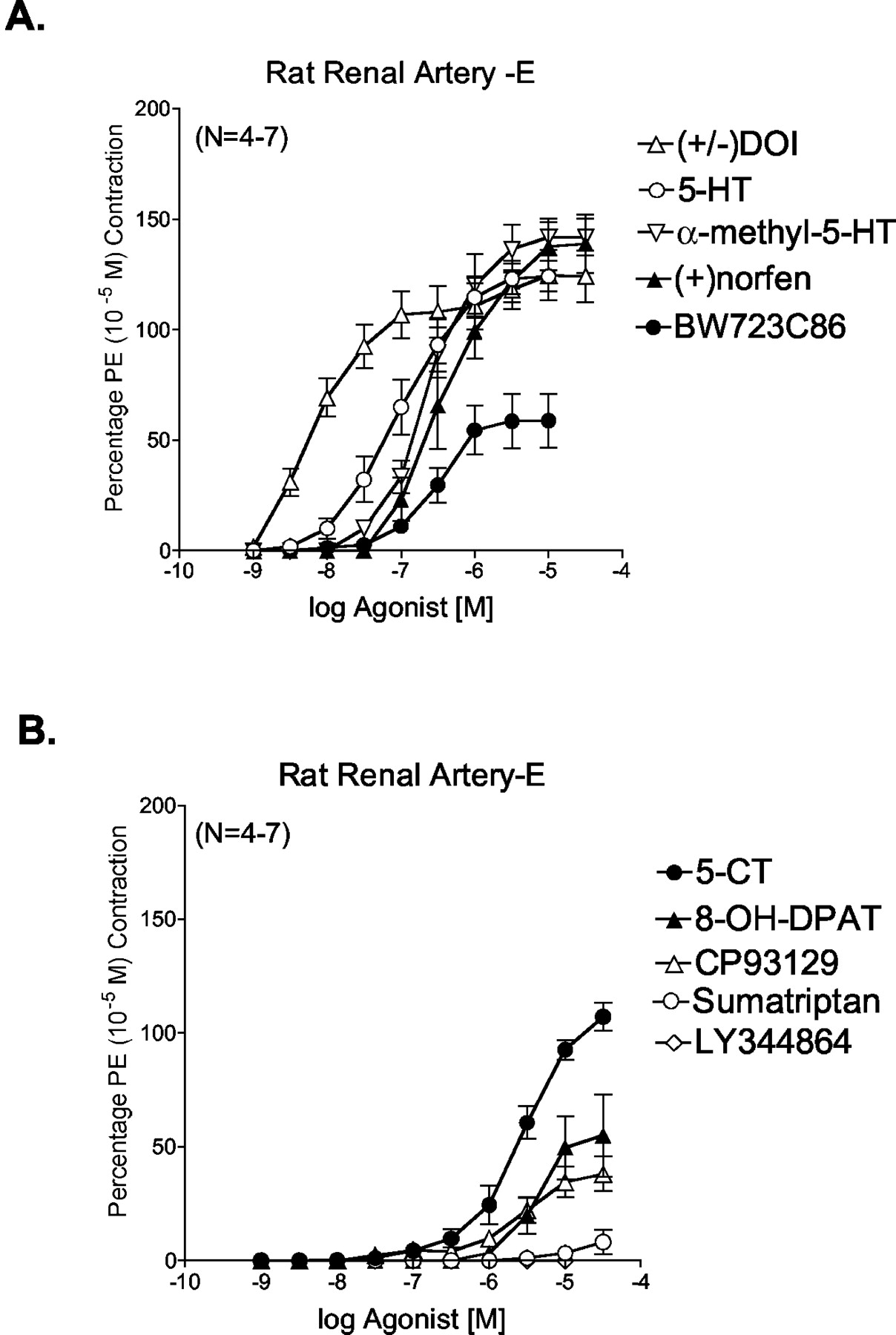
Use of 5-HT Receptor Agonists in Rat Renal Artery.Figure 2, A and B, depicts some of the cumulative concentration-response curves derived from testing 13 different 5-HT receptor agonists. Data for agonists for which curves are not shown can be found in Table 1. These have been divided into those with higher potency and higher maximum response (Fig. 2A) and those with lower potency and lower maximum response (Fig. 2B). The following rank order of potency was observed and is detailed in Table 1: DOI > 5-HT = 5-methoxytryptamine > mCPP = α-methyl-5-HT = BW723C86 > (+)-norfenfluramine > 2-methyl-5-HT > CP93129 = 5-carboxamidotryptamine > (±)8-OH-DPAT > sumatriptan > LY344864 (inactive). Because the 5-HT1F receptor agonist LY344864 was inactive, this suggests that 5-HT1F receptors do not play a role in renal arterial contraction. Notably, the 5-HT2B receptor agonist BW723C86 caused a concentration-dependent contraction in the renal artery, although with lower maximum response than that of 5-HT (Fig. 2A) and could be antagonized by LY272015 and LY53857 (Fig. 3), although not in a strictly competitive manner. Additionally, (+)-norfenfluramine and mCPP, two agonists with appreciable affinity for the 5-HT2B receptor, contracted the rat renal artery.

Effect of 5-HT receptor agonists in contracting the endothelium-denuded right renal artery of the normotensive rat. Agonists are divided into agonists that are potent and cause a large maximum contraction (A) and agonists that are less potent and cause a smaller maximum contraction (B). Points represent means ± S.E.M. for the number of animals in parentheses indicated in parentheses. PE, phenylephrine.
Pharmacological parameters for 5-HT receptor agonist-induced contraction in endothelium-denuded rat renal artery Values are means ± S.E.M. for the number of animals in parentheses.
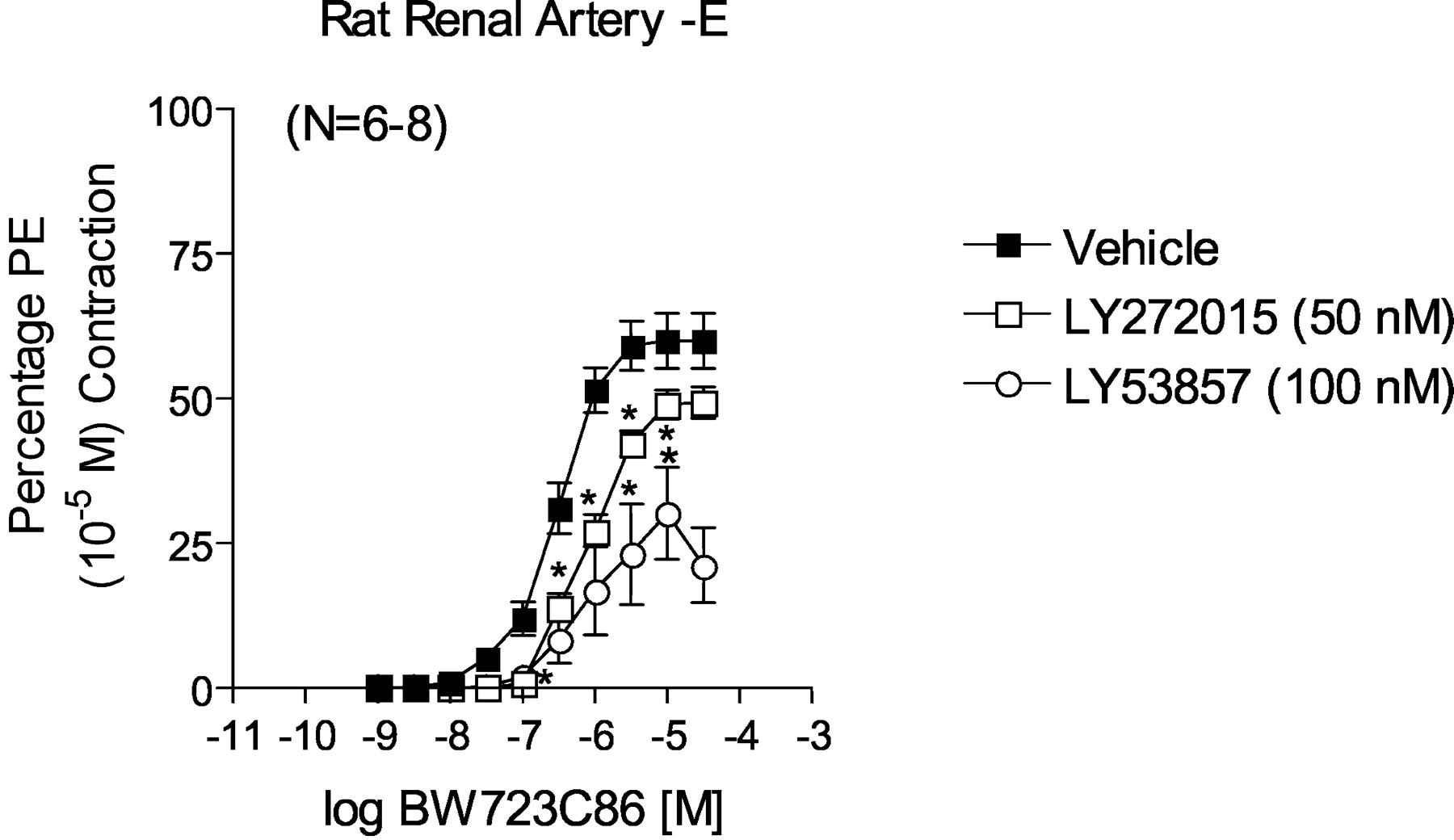
Effect of LY272015 and LY53857 on BW723C86-induced contraction in endothelium-denuded right renal artery. Points represent means ± S.E.M. for the number of animals indicated in parentheses. The asterisk (*) indicates significant differences (p < 0.05) from vehicle. PE, phenylephrine.
The -log EC50 values listed in Table 1 were correlated with binding affinities derived from radioligand binding experiments performed in rat tissues and reported in the investigator-driven Physician Scientist Development Program Drug database (http://pdsp.cwru.edu/pdspf.asp). These binding affinities include measures of affinity of 12 of the 13 agonists tested with the 5-HT1B, 5-HT1F, 5-HT2A and 5-HT2B receptors. These receptors were chosen as these are the primary receptors implicated in promoting arterial smooth muscle contractility. Correlations of -log EC50 values [M] were significant for the 5-HT2A (r = 0.928) and 5-HT2B receptor binding affinities (r = 0.843). Correlations with the 5-HT1B and 5-HT1F receptor were not significant or negative.
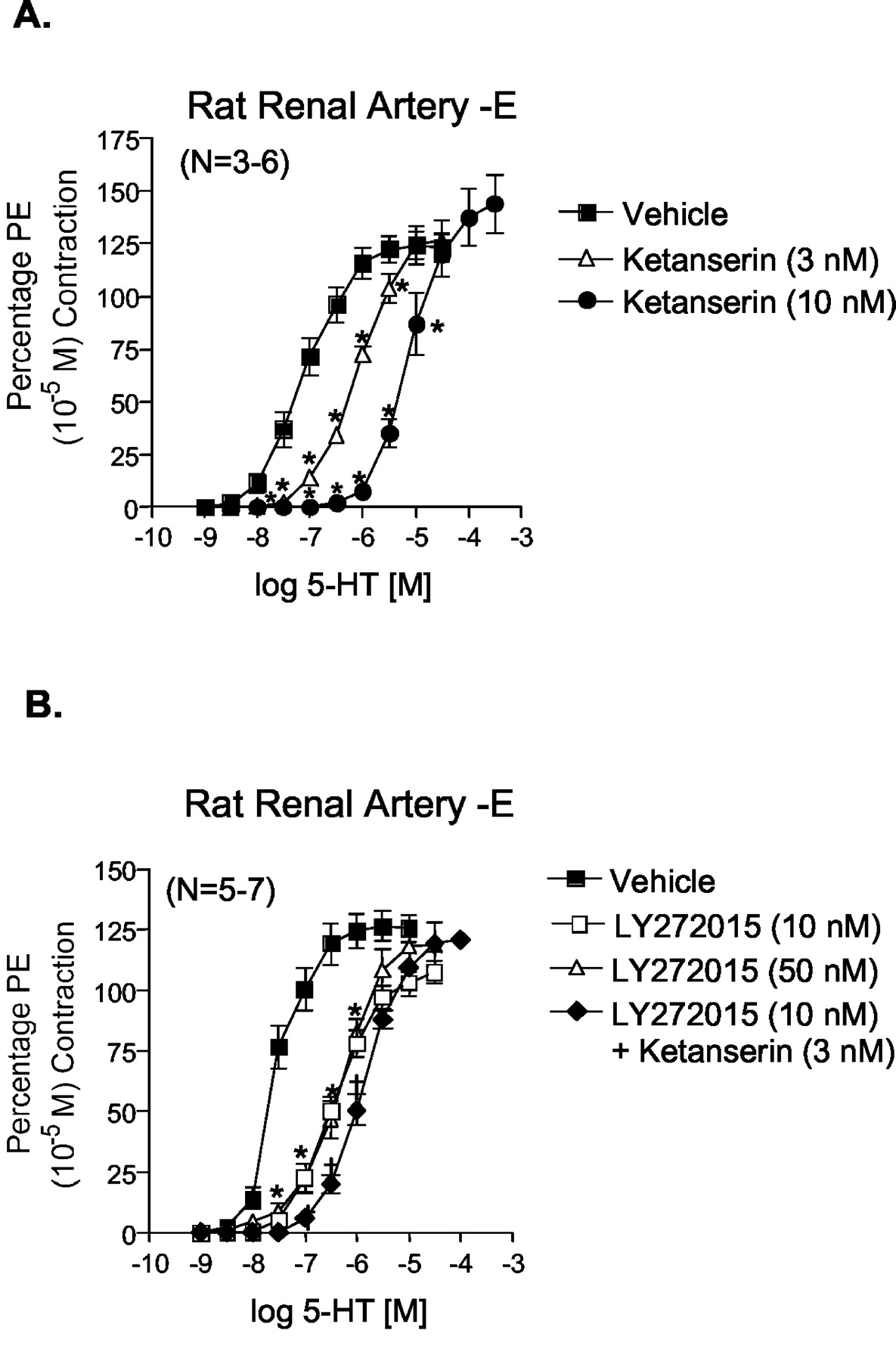
Use of 5-HT Receptor Antagonists in Rat Renal Artery. To validate the involvement of the 5-HT2A and 5-HT2B receptors in 5-HT-induced contraction, we tested the ability of 5-HT receptor antagonists with known affinity for the 5-HT2A and/or the 5-HT2B receptor to inhibit renal arterial 5-HT-induced contraction. Figure 4 demonstrates that the 5-HT2A/5-HT2C receptor antagonist ketanserin (3, 10 nM; pKB = 9.9 ± 0.02) and 5-HT2B receptor antagonist LY272015 (10 nM; pKB = 8.59 ± 0.03) significantly shifted 5-HT-induced contraction in a parallel and competitive fashion. The pKB value determined for ketanserin was significantly higher than expected when anticipating interaction with 5-HT2A receptors. The concentration of LY272015 used (10 nM) was sufficiently low to be assured that it does not interact with 5-HT2A receptors (Cohen et al., 1996), and rightward shifted 5-HT-induced contraction in the isolated stomach fundus with a pKB value of 9.73 ± 0.05 (data not shown). Interestingly, a higher concentration of LY272015 (50 nM) was unable to further shift 5-HT-induced contraction. However, combination of ketanserin (3 nM) and LY272015 (10 nM) caused a shift that was greater than that caused by either antagonist alone (Fig. 4B), indicating the two receptor antagonists likely block different receptor populations.

Effect of the 5-HT2A/2C receptor antagonist ketanserin (3 and 10 nM; A) and 5-HT2B receptor antagonist LY272015 (10 and 50 nM; B) with or without ketanserin (3 nM) on 5-HT-induced contraction in endothelium-denuded right renal artery. Points represent means ± S.E.M. for the number of animals indicated in parentheses. The asterisk (*) indicates significant differences (p < 0.05) from vehicle; †, statistically significant differences (p < 0.05) from LY272015 (50 nM)-incubated responses. PE, phenylephrine.
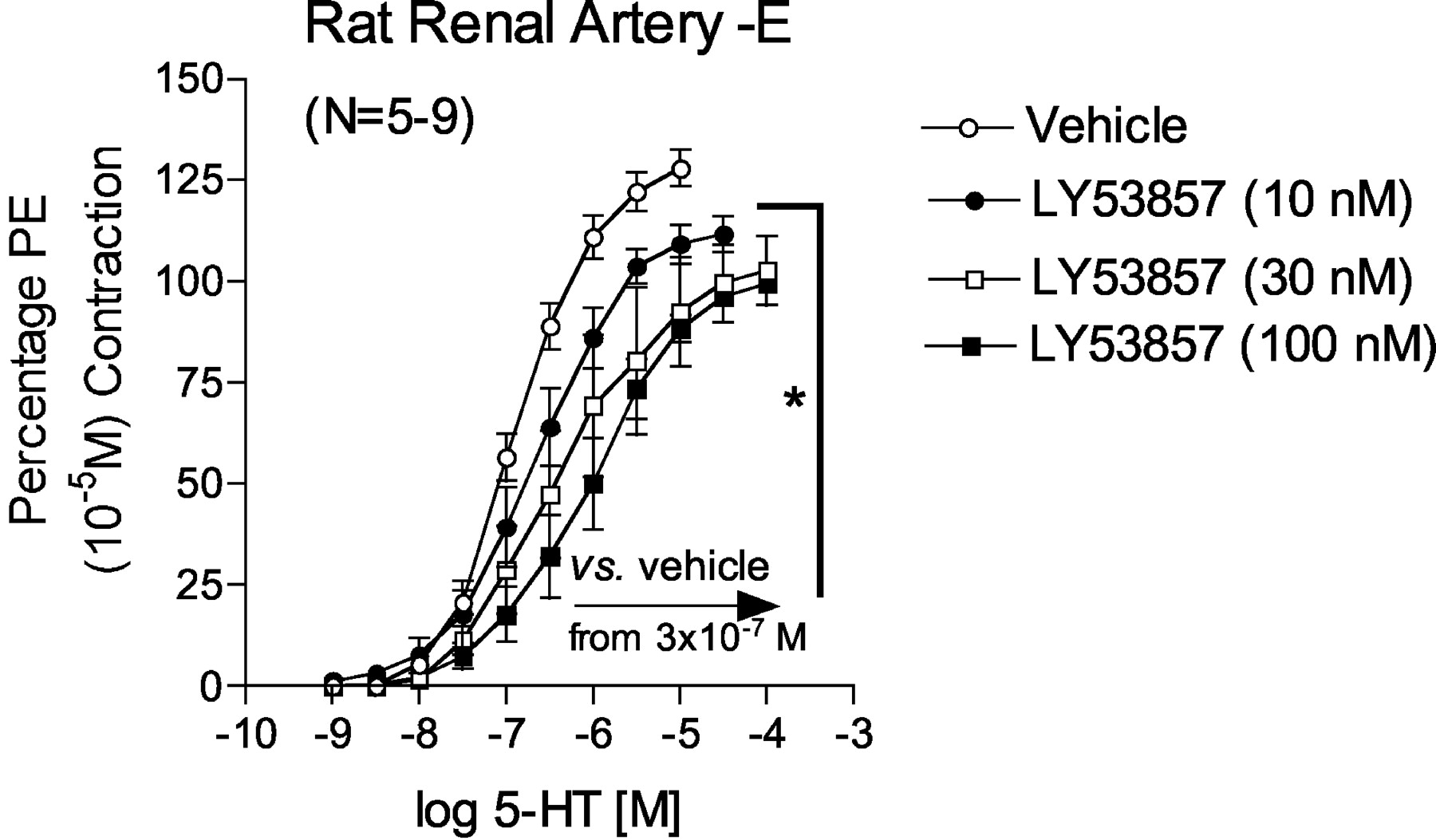
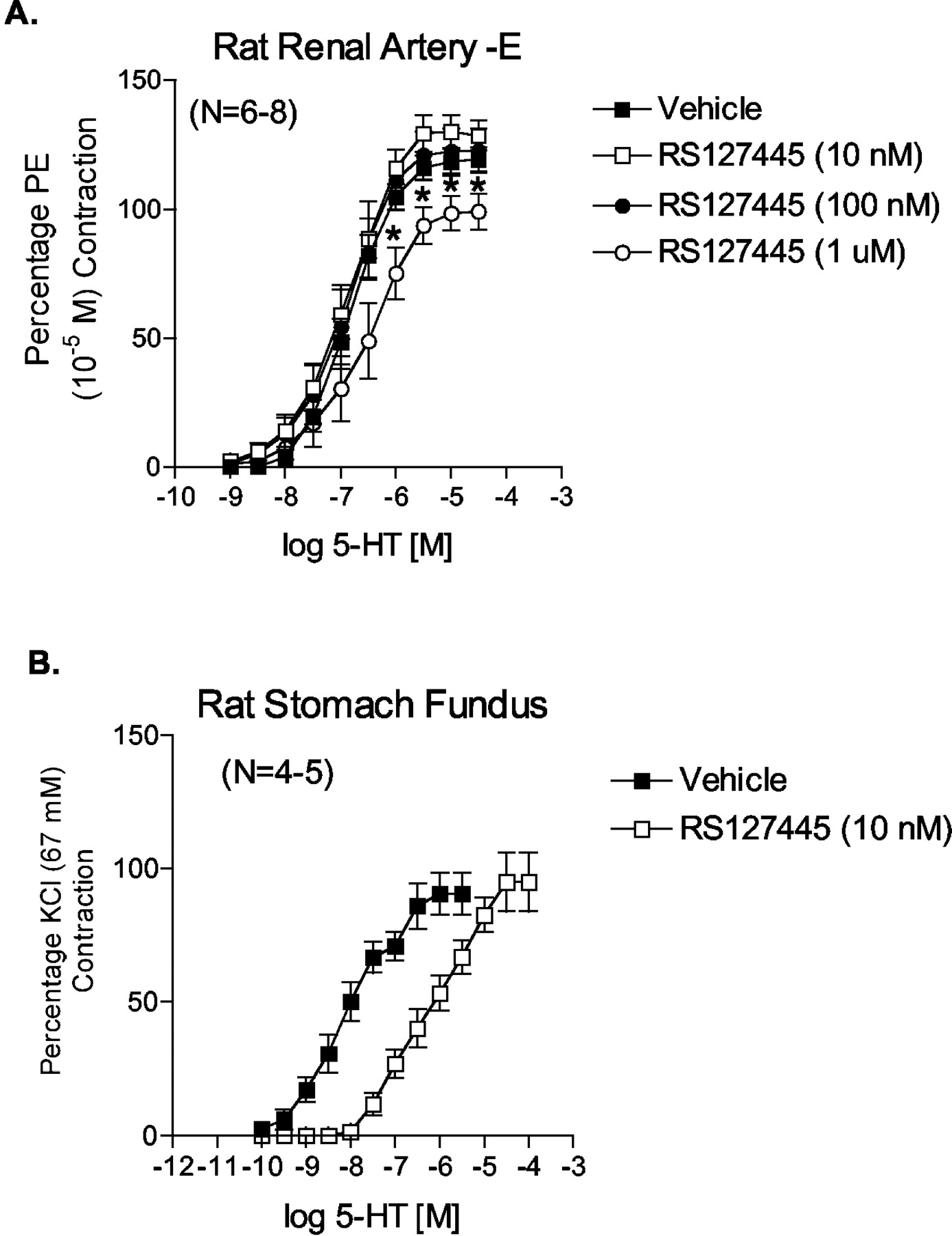
The nonselective 5-HT2 receptor antagonist LY53857 caused a nonparallel and noncompetitive antagonism of 5-HT-induced contraction (Fig. 5). The pEC50 of 5-HT and slope of the linear portion of the curves were as follows: vehicle pEC50 = 6.88 ± 0.03, slope = 60.64 ± 4.47; 10 nM LY53857 pEC50 = 6.64 ± 0.04, slope = 45.9 ± 0.78; 30 nM LY53857 pEC50 = 6.45 ± 0.07, slope = 33.33 ± 1.85; and 100 nM LY53857 pEC50 = 6.01 ± 0.06, slope = 36.53 ± 1.67. From the 5-HT concentration of 3 × 10-7 M and above, all concentrations of LY53857 significantly reduced contraction compared with vehicle [marked with an asterisk (*) and arrow]. A different 5-HT2B receptor antagonist, RS127445 (Fig. 6A), was unable to inhibit 5-HT-induced contraction in concentrations demonstrated in our laboratory to shift competitively 5-HT-induced contraction in the isolated rat stomach fundus, a tissue with a bona fide 5-HT2B receptor (pKB = 9.78 ± 0.13; Fig. 6B). The concentration of RS127445 had to be raised to 1 μM to inhibit 5-HT-induced contraction significantly (Fig. 6A).

Antagonism exerted by the nonselective 5-HT2 receptor antagonist LY53857 against 5-HT in the endothelium-denuded right renal artery. Points represent means ± S.E.M. for the number of animals indicated in parentheses. The asterisk (*) indicates significant differences (p < 0.05) from vehicle for all concentrations starting at the arrow. PE, phenylephrine.

Antagonism exerted by the 5-HT2B receptor antagonist RS127445 against 5-HT in the endothelium-denuded right renal artery (A) and stomach fundus (B). Points represent means ± S.E.M. for the number of animals indicated in parentheses. The asterisk (*) indicates significant differences (p < 0.05) from vehicle. PE, phenylephrine.
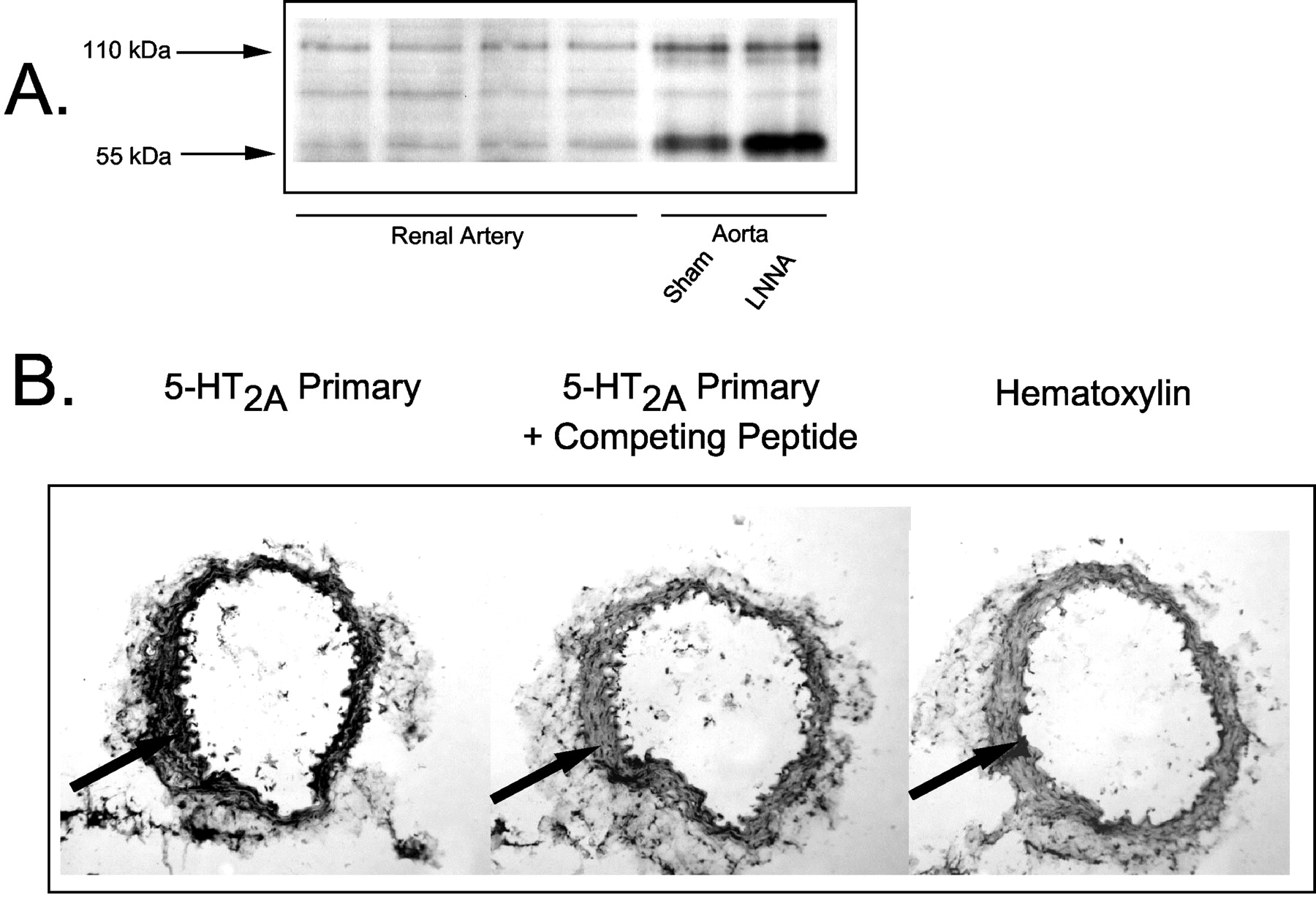
Verification of the Presence of 5-HT2A and 5-HT2B Receptor Protein in Rat Renal Artery. We next determined whether both 5-HT receptor proteins were present in the rat renal artery. Two immunoreactive 5-HT2B receptor bands in both renal and aortic homogenates were identified in Western analyses (Fig. 7A) at approximately 55 and 110 kDa; the band in between these two was observed only spuriously. Positive controls of aortic homogenates from sham normotensive and hypertensive rats were used, where a higher expression of the 5-HT2B receptor in the aorta of the rats made hypertensive by the nitric-oxide synthase inhibitor Nω-nitro-l-arginine was expected and observed (confirmed here, lower 55-kDa band densitometry sham = 5142 arbitrary units, Nω-nitro-l-arginine = 9500; published in Russell et al., 2002). The same amount of total protein (50 μg) was loaded for the renal artery and aorta. The 5-HT2B-immunoreactive protein in renal artery homogenates was expressed at a considerably lower density as compared with the aorta. We estimate that, per microgram of total protein, the renal artery expresses approximately 12% that of the normal aorta (616 ± 81 arbitrary densitometry units calculated for the lower band only because this is likely the monomeric form of the 5-HT2B receptor). Because antibodies useful in Western analyses for the 5-HT2A receptor are not currently available, we took an immunohistochemical approach to demonstrate the presence of this receptor in the renal artery. Figure 7B demonstrates that immunohistochemically detectable and specific 5-HT2A receptor immunoreactivity was observed in the smooth muscle of the renal artery (compare first with second panel). Thus, these studies demonstrate that immunoreactive 5-HT2A and 5-HT2B receptor are present in renal artery.

Western analyses demonstrating the presence of an immunorecognized 5-HT2B receptor in right renal artery (A) and immunohistochemical analysis of right renal artery detecting the presence of an immunoreactive 5-HT2A receptor (B). Experiments representative of four separate experiments performed with samples from different animals. Arbitrary densitometry units were measured using NIH Image version 1.62. Arrow points to specific binding in the area of arterial smooth muscle.
Discussion
Work by investigators has demonstrated that 5-HT acts as a pressor in the isolated perfused kidney (McGregor and Smirk, 1970; Collis and Vanhoutte, 1977; Tuncer and Vanhoutte, 1993) and constricts renal vasculature. Outside of these works, the role of 5-HT in the kidney is not well understood or studied, especially the receptor mechanism by which 5-HT exerts its effects. We present data characterizing the 5-HT receptor(s) in the right renal artery of the normotensive rat.
Expected Pharmacology. We took a classical pharmacological approach to determine the receptor(s) through which 5-HT stimulated renal arterial contraction. A minimum of two receptor populations has the potential to mediate 5-HT-induced contraction. This is evidenced by 1) the rank order of agonists in contracting the artery; 2) the correlation of potency values with literature-derived rat receptor binding affinities; 3) the antagonism exerted by carefully chosen concentrations of antagonists selective for different 5-HT2 receptors; and 4) the biochemical identification of proteins recognized by antibodies specific for the 5-HT2A or 5-HT2B receptor. It is fair to acknowledge the limitations of potency correlations with radioligand binding affinities because agonists, alone, cannot be used to characterize a receptor. Demonstration of equilibrium conditions and lack of measurable desensitization are necessary, and we have not proven these definitively occur in our system. Moreover, agonist potency is not purely defined by affinity. As elegantly discussed by Kenakin (2002), efficacy and potency are associated in complex and interesting manners. Even given these limitations, these correlations allow us to gather agonist data as a whole and evaluate the role of particular 5-HT receptors in contraction. The 5-HT1B and 5-HT1F receptors were excluded, and the 5-HT2A and 5-HT2B receptors seem to play the strongest role in mediating contraction. Importantly, mRNA for both the 5-HT2A and 5-HT2B receptor in rat renal artery has been reported (Ullmer et al., 1995), and the kidney is one of the sites in which the human 5-HT2B receptor mRNA was found to be highly expressed (Bonhaus et al., 1995). The role of these two 5-HT receptor subtypes deserves comment.
First, 5-HT was more potent in the renal artery than the aorta, suggesting that the mechanisms by which 5-HT contracts the two arteries is different. On the basis of affinity alone, the potency of 5-HT in contracting the renal artery was not consistent with interacting with a classical 5-HT2A receptor which, using [3H]-ketanserin as a radioligand, ranges anywhere from 500 nM to 2950 nM in the rat cortex (http://pdsp.cwru.edu/pdspf.asp). Similarly, micromolar affinity values for 5-HT have been observed when [3H]spiperone was used as a radioligand. One can speculate that the 5-HT2A receptors in the renal artery may be more efficiently coupled to their signaling elements than are 5-HT2A receptor in the aorta, or the renal artery expresses a unique 5-HT2A receptor for which 5-HT has a greater than normal affinity. Without a selective 5-HT2A receptor agonist, which to our knowledge is currently unavailable, this is difficult to test. However, another explanation is that a different receptor for which 5-HT possesses a higher affinity is present, and this may be a 5-HT2B-like receptor for which 5-HT possess a 300-fold higher affinity than the 5-HT2A receptor in the rat (Wainscott et al., 1993).
The ability of the 5-HT2B receptor agonist BW723C86, currently the only relatively selective 5-HT2B receptor agonist available, to cause contraction of the renal artery lends significant support to the idea of a 5-HT2B receptor being involved in 5-HT-induced contraction. This is further supported by the fact that other agonists with affinity for the 5-HT2B receptor [mCPP, DOI, (+)-norfenfluramine (Fitzgerald et al., 2000; Rothman et al., 2000)] also contracted the renal artery. Moreover, the 5-HT2B receptor antagonist LY272015 reduced not only the contraction to 5-HT but to BW723C86 as well. Finally, immunoreactive 5-HT2B receptor protein was present, although expressed at low levels compared with the aorta. However, it seems as if activation of 5-HT2B or 5-HT2B-like receptor mediates only a modest portion of contraction to 5-HT as 10 nM LY272015 was maximal in its ability to antagonize 5-HT-induced contraction, whereas ketanserin caused parallel and large shifts in contraction.
Unexpected Pharmacology. We found that antagonists that were predicted to shift a 5-HT2B receptor-mediated response, as based on their reported radioligand binding affinity, did not do so. LY53857 has nearly equimolar affinity for the 5-HT2A and 5-HT2B receptor (Wainscott et al., 1993) and was predicted to produce a large and parallel rightward shift in 5-HT-induced contraction. This is true if one considers interaction of LY53857 with the 5-HT2A or 5-HT2B receptor alone, and Fig. 5 demonstrates this was not observed. Moreover, high concentrations of LY53857 (100 nM) were necessary to antagonize contraction to BW723C86. The pKB value of the 5-HT2B receptor antagonist LY272015 in the renal artery (8.59 ± 0.03) was different from that in the fundus (9.73 ± 0.05). This suggests that the renal 5-HT receptor and fundus receptor are not the same in terms of the affinity LY272015 possesses for the receptors. Thus, the renal receptor may not be a classical 5-HT2B receptor. The lack of similarity may also be influenced by the complement of 5-HT receptors present in the renal artery or indicative of tissue heterogeneity in receptor pharmacology.
It is not only the response to LY53857 that was unexpected but the relative inability of RS127445 to block 5-HT-induced contraction. This compound has been well described as a 5-HT2 receptor antagonist (Bonhaus et al., 1999), and in our hands exerts 5-HT2B receptor antagonism as evidenced by a competitive shift in 5-HT-induced contraction in the rat stomach fundus. Thus, it is reasonable to reiterate that the fundus 5-HT receptor is different from the renal 5-HT receptor(s). Blockade by neutral antagonists should not be dependent on coupling efficiency, and we know of no evidence that suggests that either LY272015, LY53857, or RS127445 exerts any inverse agonism. Thus, the receptor or receptor unit must be of a different character.
Another explanation for these findings, which is purely speculative, is that the combined presence of the 5-HT2A and 5-HT2B receptor may modify the ability of receptor antagonists to inhibit contraction. Such receptor interaction and consequent modification of receptor pharmacology has been reported for endothelin receptors (Lodge et al., 1995; Adner et al., 2001; Watts et al., 2002) and more recently for the 5-HT1B and 5-HT1D receptors (Xie et al., 1999). In this interaction, receptor proteins heterodimerize to form a pharmacologically distinct and functional unit. We are currently unable to test the idea that the 5-HT2A and 5-HT2B interact physically because antibodies for the 5-HT2A receptor that are amenable to immunoprecipitation/coimmunoprecipitation, and Western studies are not available. Thus, the idea of 5-HT2A and 5-HT2B receptor interaction must remain a speculation.
Perspectives. Some of these questions may not be answered until the 5-HT receptor(s) is/are cloned from the renal artery, or until the issue of receptor interaction and efficiency is addressed. The 5-HT2B receptor in the rat stomach fundus was cloned in 1992 (Foguet et al., 1992; Kursar et al., 1992), pharmacologically described in 1993 (Wainscott et al., 1993), and is a member of the heptahelical superfamily. The human and rat 5-HT2B receptor differ in terms of the affinity possessed by ketanserin for each receptor (Wainscott et al., 1996). Regardless of the outcome of such studies, there is sufficient evidence to suggest the presence of a functional 5-HT2B-like receptor in the rat renal artery that is part of the unique pharmacology of the 5-HT receptors in the renal artery.
These findings are physiologically relevant given the relatively sparse research performed in understanding the role of 5-HT in control of renal function. The finding of a unique pharmacology may present a similarly unique means by which to affect renal arterial function. 5-HT, as does mCPP, increases perfusion pressure in the in situ autoperfused rat kidney (Moran et al., 1997). It is important to note that the two receptors identified in the renal artery, the 5-HT2A and 5-HT2B or 5-HT2B –like, also serve the demonstrable function of 5-HT-mediated cellular mitogenesis, growth, and tissue formation. Studies in the vasculature demonstrate the ability of the 5-HT2A receptor to mediate 5-HT-induced mitogenesis (Grewal et al., 1999), as measured by [3H]thymidine uptake, and work using the 5-HT2B receptor knockout mouse underscores the importance of this receptor to the normal formation of the heart (Nebigil et al., 2000) and gastrointestinal system (Fiorica-Howells et al., 2000). The 5-HT2B receptor has also been described as necessary for development of pulmonary hypertension induced by hypoxia (Launay et al., 2002) and for the pathology associated with fenfluramine-induced damage of cardiac valves (Fitzgerald et al., 2000). Although not studied here, both receptor subtypes could potentially contribute to renal arterial narrowing and thus contribute to renal disease.
In summary, these findings present evidence of a complex pharmacology of 5-HT receptors mediating contraction to 5-HT in the normal right renal artery of the rat. Although the pharmacology of this receptor is curious, the finding of such a receptor(s) response raises questions as to whether 5-HT receptor coupling is tissue-dependent, whether 5-HT receptor heterogeneity exists, or whether coexistence of two 5-HT2 receptor subtypes enables a unique pharmacology.
Footnotes
-
This work was supported by National Institutes of Health Grant HL58489.
-
Article, publication date, and citation information can be found at http://jpet.aspetjournals.org.
-
DOI: 10.1124/jpet.103.062562.
-
ABBREVIATIONS: 5-HT, 5-hydroxytryptamine; DOI, 1-(2,5-dimethoxy-4-iodophenyl)-2-aminopropane; BW723C86, α-methyl-5-(2-thienylmethoxy)-1H-indole-3-ethanamine hydrochloride; (±)8-OH-DPAT, (±)-8-hydroxy-2-(di-n-propylamino)tetralin; TBS, Tris-buffered saline; PBS, phosphate-buffered saline; mCPP, 1-(m-chlorophenyl)piperazine; LY53857, 6-methyl-1-(1-methylethyl)-ergoline-8b-carboxylic acid 2-hydroxy-1-methylpropyl ester maleate; LY272015, 6-methyl-1,2,3,4-tetrahydro-1-[3,4-dimethoxyphenyl)methyl-9H-pyrido[3,4b]indole] hydrochloride; LY344864, (R)-N-[3-dimethylamino-2,3,4,9-tetrahydro-1H-carbazol-6-yl]-4-fluorobenzamide; RS127445, 2-amino-4-(4-fluoronaphth-1-yl)-6-isopropylpyrimidine; CP93129, 1,4-dihydro-3-(1,2,3,6-tetrahydro-4-pyridinyl)-5H-pyrrol[3,2-b]pyridin-5-one dihydrochloride; MDL 100907, (±)-2,3-dimethoxyphenyl-1-[2-(4-piperidine)-methanol].
- Received November 5, 2003.
- Accepted December 8, 2003.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}