


Abstract
The neuropeptide Y (NPY) family peptides NPY, peptide YY (PYY), and pancreatic polypeptide (PP) bind to four G protein-coupled receptors (GPCRs): Y1, Y2, Y4, and Y5. A key step in the desensitization and internalization of GPCRs is the association of the receptor with β-arrestins. In the present study, these receptors were analyzed with respect to their ability to interact with GFP2-tagged β-arrestin 2 using the new bioluminescence resonance energy transfer 2 method. Agonists induced a concentration-dependent association of β-arrestin 2 with all four receptors. Whereas the Y1 receptor exhibited the highest maximum response and rapid association (t½ = 3.4 min), the maximal signals for the association of Y2 and Y4 receptors were less than half of that of Y1, and the association rates were much slower. Interestingly, when evaluated at the Y4 receptor, the Y4 agonist 1229U91 [(Ile,Glu,Pro,Dpr,Tyr,Arg, Leu,Arg,Try-NH2)-2-cyclic(2,4′),(2′,4)-diamide] was unable to provoke the same maximal response as human PP, suggesting that 1229U91 is a partial agonist. When stimulated by PYY, the Y5 receptor responded with a t½ of 4.6 min and a maximal response approximately 60% of what was observed with Y1. Because β-arrestins are key components in GPCR internalization, it is interesting to note that the receptor that is known to internalize rapidly (Y1) exhibits the most rapid association with β-arrestin 2, whereas the receptor that is known to internalize slowly, or not at all (Y2) associates slowly with β-arrestin 2.
Neuropeptide Y (NPY), peptide YY (PYY), and pancreatic polypeptide (PP) have profound central effects on energy intake, mood, and reproduction and act through four G protein-coupled receptors (GPCRs) in primates, namely, Y1, Y2, Y4, and Y5.
The pharmacological profiles and distribution of these receptors have been studied extensively (for review, see Berglund et al., 2003). The Y1, Y2, and Y5 receptors bind NPY and PYY with higher affinity than PP, whereas Y4 displays the reverse rank order of potencies (PP > PYY ≥ NPY). The Y1, Y2, and Y5 receptors can be distinguished from one another by several subtype-selective antagonists such as BIBP3226 (Y1) (Rudolf et al., 1994), BIIE0246 (Y2) (Doods et al., 1999), and CGP71683A (Y5) (Criscione et al., 1998) and the similar compound Novartis 1 (Pronchuk et al., 2002) used here. Furthermore, the Y2 receptor pharmacology is differentiated from that of the other receptors in its ability to bind N-terminally truncated fragments of NPY and PYY such as NPY/PYY13-36 with similar affinity as the native peptides.
All four NPY-family receptors are found both in the brain and in peripheral tissues. Like many peptide GPCRs, the NPY family receptors all couple negatively to adenylyl cyclase, inhibiting cAMP synthesis. Most GPCRs are desensitized shortly after agonist exposure, making them unable to respond to a new signal. This may ultimately lead to internalization of the receptor. The Y1 and Y4 receptors have been shown to internalize rapidly upon agonist binding (Fabry et al., 2000; Parker et al., 2001; Gicquiaux et al., 2002; Parker et al., 2002), whereas it has been suggested that Y2 does not internalize or does so very slowly (Parker et al., 2001; Gicquiaux et al., 2002).
Internalization of GPCRs is mediated by a family of proteins called β-arrestins (for reviews, see McDonald and Lefkowitz, 2001; Pierce and Lefkowitz, 2001). There are four known forms of arrestin. Arrestin 1 and cone arrestin are exclusively expressed in the visual cells of the retina, whereas arrestin 2 and 3 (also known as β-arrestin 1 and 2) have a much broader distribution. β-Arrestins bind to the activated (phosphorylated) form of GPCRs, inhibiting further interaction with G proteins and, instead, link the receptor to the cell surface protein clathrin. Clathrin-coated pits are a key component of the internalization machinery (Takei and Haucke, 2001), and after internalization, GPCRs are either recycled to the cell surface or degraded in lyzosomes. β-Arrestin 2 is relatively promiscuous and can couple to many GPCRs, whereas β-arrestin 1 binds to a more limited portfolio of receptors (Pierce and Lefkowitz, 2001). GPCRs can sometimes be categorized based on how they interact with β-arrestins (Pierce and Lefkowitz, 2001). Class A receptors interact primarily with β-arrestin 2 and unhook from β-arrestins at or near the cell surface before internalization. In contrast, class B receptors remain associated with β-arrestins (1 and 2) during internalization after agonist exposure. Several groups have identified molecular recognition sites in GPCRs for β-arrestins. These include phosphorylation sites in the form of clusters of serines and/or threonines at the C terminus (Oakley et al., 2001) that seem to be essential for class B type interaction. In addition, stretches of basic amino acids in beginning and the end of the third intracellular loop have also been identified as interaction points between β-arrestins and GPCRs (DeGraff et al., 2002).
Bioluminescence resonance energy transfer (BRET) is a method that can be used to explore protein-protein interaction (Xu et al., 1999). It is a natural process that occurs in many organisms that emit light (Xu et al., 1999). When Renilla luciferase (Rluc) catalyzes the reaction coelenterazine → coelenteramide, blue light (λ = 410 nm) is emitted. If present in proximity (within 10 nm; Xu et al., 1999), green fluorescent protein (GFP) can act as an acceptor for the blue photon and reemit light in the green spectra (λ = 515 nM for GFP2, the mutated version of GFP used here). BRET has mainly been used to study GPCR dimerization (for review, see Angers et al., 2002) but also for several other applications (Xu et al., 1999; Boute et al., 2001).
BRET2 differs from BRET in that it uses a mutated variant of GFP, GFP2, and a modified variant of the Rluc substrate to increase the spectral resolution. Although several groups have reported desensitization and internalization of NPY family receptors, nothing has been reported to date about the interaction of these receptors with β-arrestins. In this study, we have used a GFP2-tagged β-arrestin 2 to further explore and compare the interaction of β-arrestin 2 with NPY family receptors.
Materials and Methods
Creation of Vector Constructs and Cell Lines. The cloning of the rhesus Y1, Y2, and Y5 receptors has been described elsewhere (Gehlert et al., 2001). To clone the rhesus Y4 receptor, two primers based on the 5′- and 3′-untranslated sequences of the human Y4 receptor gene were synthesized. The 5′ primer had the sequence 5′-GTCCTGGAATCTTTTCACATCCACT-3′ and the 3′ primer had the sequence 5′-GCAGGGAGAAGACCTAGACCTGG-3′. The PCR cycle (94°C for 30 s, 55°C for 40 s, and 68°C for 1 min 30 s, 35 cycles) using 100 ng of genomic DNA from rhesus monkey (BD Biosciences Clontech, Palo Alto, CA) as template and Pfu polymerase (Invitrogen, Carlsbad, CA) generated a band of the expected length (1.1 kilobases). The band was run on an agarose gel, cut out, and purified using a gel extraction kit (QIAGEN, Valencia, CA). The purified band was sequenced using an ABI377 automated sequencer using the two primers mentioned above as sequencing primers. Based on the sequence of the PCR fragment, one forward primer was synthesized: rhY4fH containing a HindIII site with the sequence 5′-AAGCTTAAGCTTACCATGAACACCTCTCACCTCCT-3′ and one reverse primer rhY4rNSKS with the sequence 5′-CCGCGGTACCAATGGGATTGGACCTGC-3′, containing a KpnI and a SacI site but lacking the stop codon to make the carboxy-terminally tagged constructs. Similarly, expression constructs for Rluc-tagged Y1, Y2, and Y5 receptors were generated using PCR by inserting the coding sequence for each receptor without the stop codon into the vector pRluc-N2 (PerkinElmer Life Sciences, Montreal, QC, Canada) directly after the HindIII site and before the KpnI site of the cloning linker. All constructs were sequenced to confirm the correct sequence of the inserts.
The GFP2-β-arrestin 2-expressing vector was purchased from PerkinElmer Life Sciences and used to generate a human embryonic kidney (HEK) 293 cell line stably expressing GFP2-tagged β-arrestin 2.
Binding Studies. HEK293 cells were grown in Dulbecco's modified Eagle's medium:F-12 mix (3:1; Invitrogen) supplemented with 5% fetal bovine serum (Invitrogen) and 20 mM HEPES (Invitrogen) at 37°C in 5% CO2. LipofectAMINE 2000 (Invitrogen) and 20 μg of DNA for the Y1, Y4, and Y5 receptors and 10 μg for the Y2 receptor was used per 150-mm dish of HEK293 cells to transiently express the various Rluc-tagged receptors. Three days after transfection, cells were harvested by scraping and centrifuged in a swing-out centrifuge. Pellets of Rluc-tagged rhY1-, rhY2-, and rhY4-expressing cells were frozen as aliquots at –80°C, whereas rhY5-Rluc membranes were used immediately to avoid receptor degradation. Radioligand binding assays were conducted on isolated crude membrane homogenates as described previously (Gehlert et al., 1992) using iodinated human (h) PYY (Y1, Y2, and Y5) or hPP (Y4) as radioligands. Non-specific binding was defined as the amount of radioactivity remaining on the filter after incubating in the presence of 0.1 μM hPYY (American Peptide Co., Inc., Sunnyvale, CA) for Y1, Y2, and Y5 or 0.1 μM hPP for Y4 (American Peptide Co., Inc.). For saturation binding analysis, cell homogenates from HEK293 cells expressing the various Rluc-tagged receptors were incubated with 12 different concentrations of radioligand for2hat room temperature. The results were analyzed using the Prism software package (GraphPad Software Inc., San Diego, CA). Protein concentrations were measured using Coomassie Plus protein assay reagent (Pierce Chemical, Rockford, IL) using bovine serum albumin standards (Bradford method). The agonists and antagonists used for the BRET2 studies were also tested for their ability to compete with the radioligands. At the rhY1 receptor, hPYY and BIBP3226, (American Peptide Co., Inc.) were tested against 125I-hPYY binding. At rhY2, hPYY and PYY13-36 (Bachem, King of Prussia, PA) were used and at rhY5, hPYY and the Y5-selective antagonist Novartis 1 (Eli Lilly & Co., Indianapolis, IN) were tested. Human PP (American Peptide Co., Inc.) and the bridged anti-parallel dipeptide compound 1229U91 (Daniels et al., 1995; Parker et al., 1998; Schober et al., 1998) (Eli Lilly & Co.) were tested at the rhY4 receptor, against 125I-hPP binding.
Time Dependence in Agonist-Induced Interaction of Rluc-Tagged Receptor with β-Arrestin 2. HEK293 cells stably expressing GFP2-tagged β-arrestin 2 were grown in 90-mm dishes (Falcon Plastics, Oxnard, CA) and transfected by 10 μg of DNA per plate for Rluc-tagged Y1, Y4, and Y5 receptors and 2.5 μg for the Y2 receptor 72 h before experiment. Cells were detached by washing with 10 ml of BRET2 buffer (phosphate-buffered saline with glucose, 1 g/l), spun for 5 min at 200g, and resuspended in 1 ml of BRET2 buffer. Twenty microliters of cell suspension was dispensed into each well of a 96-well plate and 5 μl of agonist (hPYY at 1 μM final concentration for Y1, Y2, and Y5 or 1 μM hPP for Y4) was added to each well at various time points ranging from 1 to 60 min before the BRET2 assay. For the rhY2 receptor, the longest incubation was 120 min. Each time point as well as unstimulated cells was assayed in duplicate. To assay the BRET2 ratio for all samples as well as for the blank (untransfected HEK293 cells), positive control (BRET + fusion protein consisting of GFP2 in the N terminus and Rluc in the C terminus), and negative control (Rluc in the absence of GFP2), 25 μl of DeepBlueC diluted 1:100 (final concentration 5 μM) was added to each well and the plate was immediately counted in a Fusion (PerkinElmer Life Sciences) instrument. The emission from each well was counted at λ = 410 nm (Rluc optimum) and λ = 515 nM (GFP2 optimum). The BRET2 ratio for each sample was calculated as follows: (sample515 nm – blank515 nm)/(sample410 nm – blank410 nm) – baseline, using the Excel software (Microsoft, Redmond, WA). The baseline signal was defined as the BRET2 ratio from HEK293 cells transiently transfected with pRluc-N2 (i.e., Renilla luciferase expressed in the cytosol). Association rates were calculated using Prism (GraphPad Software, Inc.).
Antagonist Effect on Agonist Induced Interaction between β-Arrestin 2 and Rluc-Tagged rhY1 and rhY5 Receptors. Cells were prepared as described above except that they were resuspended in 2 ml of BRET2 buffer. Cells were preincubated together with antagonist for 20 min and, subsequently, agonist (hPYY at final concentrations ranging from 100 pM to 10 μM) was added to a final volume of 25 μl for another 20 min. For the rhY1 receptor, the antagonist BIBP3226 (Rudolf et al., 1994) was used and for the rhY5 receptor, Novartis 1 (Eli Lilly & Co.), a modified version of the Y5 selective antagonist CGP71683A, was used. BRET2 ratios were assayed as described above.
Agonist-Induced Interaction between β-Arrestin 2 and Rluc-Tagged rhY2 and rhY4 Receptors. Due to the lack of commercially available antagonists for the Y2 receptor and the total lack of an antagonist selective for the Y4 receptor, these receptors were characterized using selective agonists. Human PYY and PYY13-36 at final concentrations ranging from 3 nM to 10 μM were added to cells expressing the rhY2-Rluc. After a 60-min incubation at room temperature, BRET2 ratios were assayed and calculated as described above. At the rhY4-Rluc receptor hPP and 1229U91 were tested at concentrations ranging from 100 pM to 10 μM. BRET2 ratios for the rhY4 receptor were assayed after a 20-min incubation with the agonists at room temperature.
Results
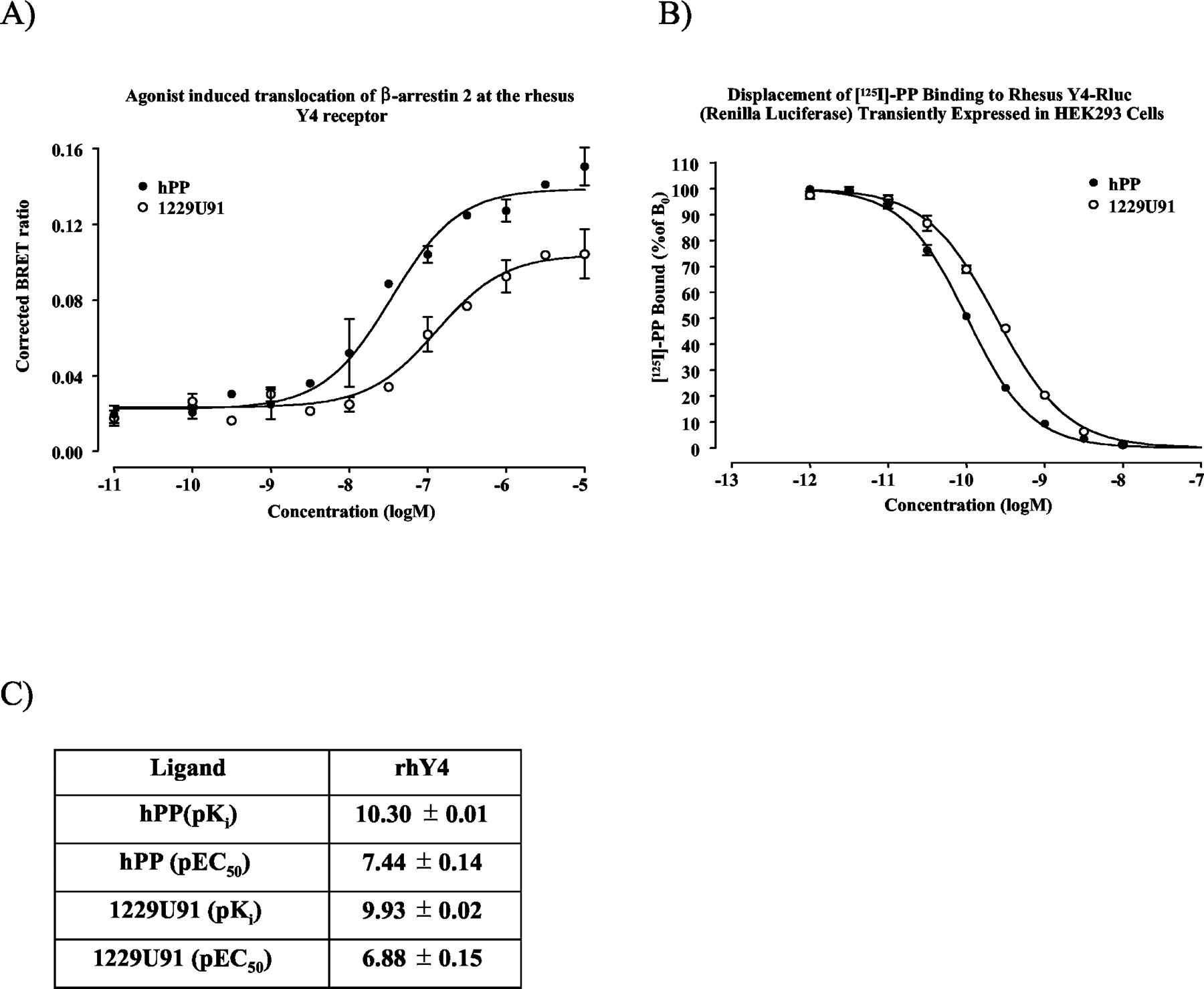
Cloning, Expression, and Pharmacological Evaluation of Rluc-Tagged NPY Family Receptors. The rhesus Y4 receptor was cloned using PCR with primers based on the human Y4 receptor gene and the sequence has been submitted to GenBank with accession number AY149475. The Rluc-tagged rhesus Y1, Y2, Y4, and Y5 receptors were transiently transfected into HEK293 cells for pharmacological evaluation and in HEK293 cells stably expressing GFP2 tagged β-arrestin 2 for studies of agonist induced β-arrestin 2 interaction. The Rluc-tagged Y1, Y2, and Y5 receptors bound 125I-hPYY and Y4 bound 125I-hPP according to saturable one-site models with dissociation constants (Kd) of 118 ± 29, 13.5 ± 2.0, 46.4 ± 1.7, and 1030 ± 120 pM, respectively, and Bmax values of 245 ± 21, 1272 ± 45, 425 ± 5, and 782 ± 47 fmol/mg protein, for Y1, Y2, Y4, and Y5, respectively (Fig. 1, A–D). Human PYY displaced 125I-hPYY binding at the Rluc-tagged rhesus Y1, Y2, and Y5 receptors with pKi values of 9.45 ± 0.03, 9.93 ± 0.05, and 8.32 ± 0.04, respectively (Figs. 3C, 5B, and 4C). Furthermore, BIBP3226 displaced with a pKi value of 8.41 ± 0.04 at the rhY1-Rluc, hPYY13-36 with a pKi value of 9.30 ± 0.04 at the rhY2-Rluc, and Novartis 1 with a pKi value of 6.69 ± 0.06 at the rhY5-Rluc receptor. Human PP and 1229U91 displaced 125I-hPP binding at the Rluc-tagged Y4 receptor with pKi values of 10.30 ± 0.01 and 9.93 ± 0.02, respectively (Fig. 6B).

Saturation binding of 125I-hPYY to Rluc-tagged NPY receptors from rhesus monkey Y1 (A), Y2 (B), and Y5 (D) and 125I-hPP to the Rluc tagged Y4 receptor (C). Data represent the mean ± S.E.M. for four experiments performed in duplicate.

Pharmacological evaluation of the rhY1 receptor. A, BRET2 study using the Y1-selective antagonist BIBP3226 at 0.01, 0.1 and 1 μM exhibits a shift to the right in the concentration-response curve for hPYY at the Y1 receptor to the right. B, Schild plot analysis of the BIBP3226 effect on hPYY-induced interaction with GFP2-tagged β-arrestin 2. C, 11 point displacement curves were used to determine the affinity hPYY and BIBP3226 when competing with 125I-hPYY binding to the Rluc-tagged rhesus Y1 receptor. Each value represents an average of four independent determinations performed in quadruplicate (mean ± S.E.M.). Data are presented in D.

Pharmacological evaluation of the rhY2 receptor. A, human PYY and the Y2-selective agonist PYY13-36 induce a concentration-dependent interaction with GFP2-tagged β-arrestin 2 displayed as an increase in BRET2 ratio. B, 11 point displacement curves were used to determine the affinities of hPYY and PYY13-36 when competing with 125I-hPYY binding to the Rluc-tagged rhesus Y2 receptor. Each value represents an average of four independent determinations performed in quadruplicate (mean ± S.E.M.). Data are presented in C.
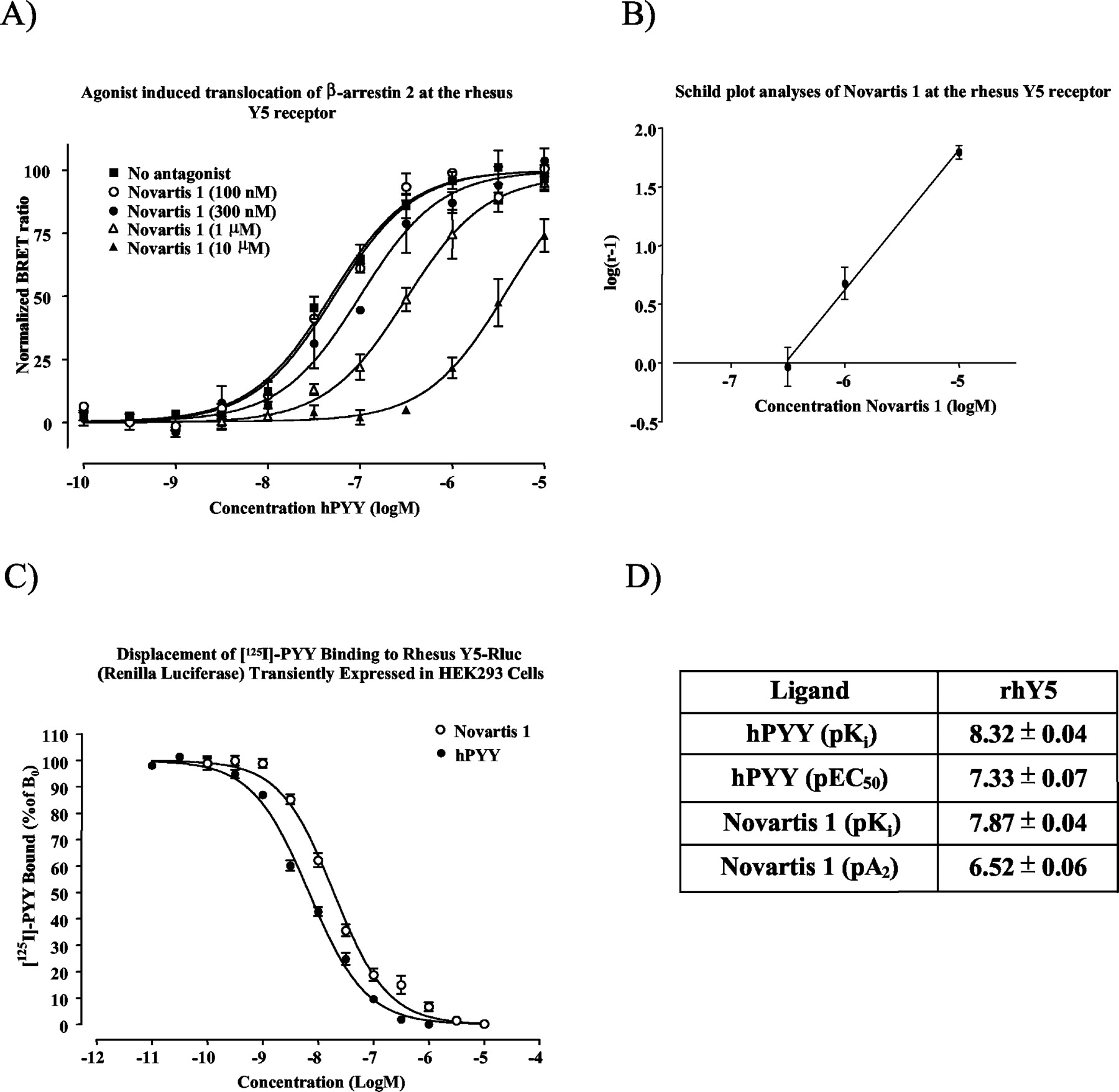
Pharmacological evaluation of the rhY5 receptor. A, BRET2 study using the Y5-selective antagonist Novartis 1 at 0.3, 1, and 10 μM exhibits a shift to the right in the concentration-response curve of hPYY at the Y5 receptor to the right. B, Schild plot analyses of the Novartis 1 effect on hPYY induced interaction with GFP2-tagged β-arrestin 2. C, 11 point displacement curves were used to determine the affinities of hPYY and Novartis 1 when competing with 125I-hPYY binding to the Rluc-tagged rhesus Y5 receptor. Each value represents an average of four independent determinations performed in quadruplicate (mean ± S.E.M.). Data are presented in D.

Pharmacological evaluation of the rhY4 receptor. A, human PP and the Y4 selective agonist 1229U91 induce a concentration-dependent interaction with GFP2-tagged β-arrestin 2 displayed as an increase in BRET2 ratio. B, 11 point displacement curves were used to determine the affinities of hPP and 1229U91 when competing with 125I-hPP binding to the Rluc-tagged rhesus Y4 receptor. Each value represents an average of four independent determinations performed in quadruplicate (mean ± S.E.M.). Data are presented in C.
Time Dependence in Agonist-Induced Interaction of Rluc-Tagged Receptor with β-Arrestin 2. Kinetic studies of the rhesus NPY receptors revealed that stimulation by 1 μM agonist (hPYY for Y1, Y2, and Y5 or hPP for Y4) produced a time-dependent increase in the BRET2 ratios for all four receptors. The t1/2 values were 3.4 ± 0.1, 23 ± 6, 8.7 ± 0.9, and 4.6 ± 0.6 min, and the maximal responses (increase in BRET2 ratio) over baseline were 0.1916 ± 0.0045, 0.0681 ± 0.0065, 0.0803 ± 0.0072, and 0.1217 ± 0.0037, for Y1, Y2, Y4, and Y5, respectively (Fig. 2, A–D).

Kinetic study using BRET2 to assay NPY receptor agonist-induced association of GFP2-tagged β-arrestin 2 with Y receptors. HEK293 cells stably expressing GFP2-tagged β-arrestin 2 were transiently transfected with Rluc-tagged rhesus Y1 (A), Y2 (B), Y4 (C), and Y5 (D) receptors. At the indicated time points, 5 μl of agonist (final concentration 1 μM hPYY for Y1, Y2, and Y5 and 1 μM hPP for Y4) was added to 20 μl of cell suspension in BRET2 buffer. The BRET2 ratio calculated from nonstimulated cells was subtracted from each data point. Graphs represent three to four experiments performed in duplicate. The t1/2 and maximal responses are calculated from each individual experiment and are expressed as mean ± S.E.M.
Schild Plot Analyses of the Rhesus Y1 and Y5 Receptors. Human PYY induced a robust concentration-dependent increase in the BRET2 signal with pEC50 values of 7.32 ± 0.06 and 7.33 ± 0.07 for the Y1 and Y5 receptors, respectively (Figs. 3A and 4A). At the Y1 receptor, the Y1-selective antagonist BIBP3226 at 0.01, 0.1, and 1 μM shifted the concentration-response curve for hPYY 2.8-, 10-, and 62-fold in agreement of a competitive antagonist, yielding a pA2 value of 8.31 ± 0.16 (Fig. 3A). At the Y5 receptor, the antagonist Novartis 1 at 0.3, 1, and 10 μM shifted the concentration-response curve for hPYY 2.1-, 5.9-, and 63-fold, yielding a pA2 value of 6.52 ± 0.06 (Fig. 4A).
Agonist-Induced Interaction between β-Arrestin 2 and Rluc-Tagged rhY2 and rhY4 Receptors. Because of the lack of available antagonists selective for the Y2 and Y4 receptors, these receptors were evaluated using the highly selective agonists PYY13-36 and 1229U91, respectively. PYY13-36 induced a concentration-dependent interaction with GFP2-tagged β-arrestin 2 with a pEC50 values of 5.52 ± 0.09. The pEC50 value for the endogenous peptide hPYY was 6.01 ± 0.10. At the rhY4 receptor, hPP and 1229U91 produced pEC50 values of 7.47 ± 0.14 and 6.88 ± 0.15, respectively. Interestingly, the response produced from 1229U91 did not reach the same maximal response as hPP at the rhY4 receptor (0.1040 ± 0.0053 compared with 0.1388 ± 0.0058; p < 0.005, t test).
Sequence Analyses. The amino acid sequences of third intracellular loop and C terminus of the rhesus NPY receptors were searched for Ser-Thr clusters such as described by Oakley et al. (2001) and for clusters of basic amino acids such as described in DeGraff et al. (2002) with the consensus sequence B-X-X-B-B, where B is either Arg or Lys and X is any amino acid. The rhY1 receptor sequence has one such Ser-Thr cluster in the C terminus (amino acids 360–363, Ser-Lys-Ser-Thr; Fig. 7). The extended third loop of the rhY5 receptor, as well as all other known mammalian Y5 receptors, contains one stretch of four consecutive serines, possibly compensating for the lack of phosphorylation sites in the C terminus of the rhY5 receptor.

Sequence alignments of the third intracellular loop and the C termini of the NPY family receptors. The extended loop of the Y5 receptor is also included. Basic amino acids (Arg/Lys) are underlined and Ser/Thr clusters (as defined in Oakley et al., 2001) are labeled by white text on black.
Discussion
GPCRs are responsible for transmitting most neuronal and endocrine signals between cells. Many signals, especially from neurotransmitters, are of a very transient nature. Proper regulation of receptor densities and attenuation of signals are, therefore, essential for cellular function. To compare the four functional NPY family receptors, we used the BRET2 method to study the effects on β-arrestin 2. Using a GFP2-tagged β-arrestin 2, it was possible to measure changes in the amount of β-arrestin 2 that was in proximity to a Rluc-tagged receptor. Stimulation of all four receptors provoked concentration-dependent shifts in the BRET2 signal. However, the potencies compared with binding affinities as well as the kinetic properties varied remarkably between the receptors.
Because all experiments in a BRET2 study are performed on Rluc-tagged receptors, it was important to confirm that addition of the 259 amino acid Rluc-protein at the C terminus did not significantly alter the binding properties of agonists and antagonists to the NPY family receptors. It was found that the affinities of the radioligands did not differ more than 3-fold from the wild-type receptors (Fig. 1; Gehlert et al., 2001). This is in agreement with findings for the human κ-opioid receptor where tagging the receptor with Rluc did not affect radioligand binding affinity (Ramsay et al., 2002). Furthermore, the displacement curves for the various subtype-selective compounds did not suggest significantly altered pharmacology (Figs. 3C, 4C, 5B, and 6B) at the rhesus Y1, Y2 (Gehlert et al., 2001), and Y4 (M. M. Berglund, unpublished data) receptors. However, hPYY displaced 125I-hPYY with about 10-fold lower affinity than at the native rhY5 receptor. The affinity of the antagonist Novartis 1 for the rhY5 receptor was also lower than published previously (Fig. 4) (Pronchuk et al., 2002). Thus, it cannot be ruled out that the Rluc-tag altered the pharmacology of the rhY5 receptor.
Agonist stimulation of the rhY1 receptor produced the largest and most rapid increase in BRET2 ratio of the NPY family receptors. An increase in the BRET2 ratio indicates association of the Rluc-tagged receptors with GFP2-tagged β-arrestin 2 and implies translocation of β-arrestin 2 to the surface. When searching for the type of class B defining phosphorylation sites, characterized by clusters of serines and threonines in the C terminus (Oakley et al., 2001; Pierce and Lefkowitz, 2001), we found that rhY1 (and all other mammalian Y1 receptors) had one such putative site (-Ser-Lys-Thr-Ser-), whereas none of the other NPY family receptors contained this motif (Fig. 7). This supports the classification of rhY1 as a class B receptor that internalizes together with β-arrestin.
The rhY5 receptor has a very short C terminus (only 17 amino acids; Fig. 7) with a third intracellular loop that is about 100 amino acids longer than in the Y1, Y2, and Y4 receptors. Therefore, it is possible that the third intracellular loop of the rhY5 receptor has taken over functions that normally reside within the C terminus. Thus, it is interesting to note the stretch of four consecutive serines in the third intracellular loop of the rhY5 receptor (Fig. 7). Stimulation of the rhY5 provoked an interaction of β-arrestin 2 almost as rapid as seen with the rhY1 receptor, although the maximal response was lower. Therefore, it is likely that also the rhY5 receptor also fits the definition of a class B receptor. However, there are currently very limited data on Y5 receptor internalization, making such classification difficult. Also, when searching for putative clusters with basic amino acids in the third intracellular loop, these are more abundant in the rhY1 and rhY5 receptors compared with Y2 and Y4 (Fig. 7), again suggesting that the Y1 and Y5 receptors would be more likely to interact with β-arrestin 2.
The rhY2 receptor was the most deviant of the four NPY family receptors with respect to β-arrestin 2 interaction. Whereas Y1 and Y5 responded rapidly to agonists by interacting with β-arrestin 2, the Y2 receptor displayed a very slow association rate and the maximal signal was only one-third of what was observed at the Y1 receptor. Furthermore, the estimated EC50 values for the agonists for hPYY and the highly Y2-selective PYY13-36 were almost 4 orders of magnitude lower than the Ki values observed in equilibrium binding assays. In previous studies, Y2 was shown not to internalize or to internalize very slowly (Parker et al., 2001; Gicquiaux et al., 2002). Based on the present data, Y2 behaves as a slow class B receptor that interacts with β-arrestin 2 and slowly internalizes with β-arrestin 2 still attached to the receptor. Another possibility is that Y2 does not internalize at all and normally does not interact with β-arrestin 2. In the latter case, one could assume that due to overexpression of GFP2-β-arrestin 2, Y2 is forced to interact with β-arrestin 2 but does so with very low affinity.
The rhY4 receptor displayed intermediate properties with regard to kinetics and maximum response (Fig. 1C). At the rhY4 receptor, hPP and 1229U91 could induce a concentration-dependent interaction with β-arrestin 2. Interestingly, the maximal effect over baseline for 1229U91 was about 30% lower than for hPP (Fig. 6A), suggesting that 1229U91 could be a partial agonist at the rhY4 receptor. 1229U91 was first characterized as a Y1 antagonist (Daniels et al., 1995) but was later found to also be a potent agonist at the Y4 receptor (Parker et al., 1998; Schober et al., 1998).
Whereas all agonists responded with potencies much lower than their binding affinities, the nonpeptide Y1-selective antagonist BIBP3226 shifted the concentration-response curve for hPYY with a pA2 value that was very similar to its equilibrium binding affinity for the rhY1 receptor (Fig. 3). In contrast, the Y5-selective antagonist used in this study, Novartis 1, was 10-fold less potent in inhibiting β-arrestin 2 interaction compared with the Ki value from the displacement binding studies (Fig. 4). Also, it was observed that Novartis 1 had very little effect on the β-arrestin association with rhY5 when coincubated with hPYY (data not shown) and thus needed to be preincubated with the receptor to have an effect. This suggests that this ligand may have a very slow association rate. On the other hand, BIBP3226 did not display any differences between preincubation or simultaneous addition with the agonist.
Generally, it is believed that all GPCRs that couple to Gi, and thus inhibit adenylyl cyclase, belong to the class A receptors with respect to β-arrestin interaction (Pierce and Lefkowitz, 2001), i.e., receptors that dissociate from β-arrestin near or at the cell surface. In fact, the same or very similar consensus sites containing basic amino acids that have been suggested to be important for β-arrestin 2 interaction (DeGraff et al., 2002) have also been implicated in Gi interaction (Wade et al., 1999). All four NPY family receptors can couple to Gi but have also been found to evoke an increase in intracellular Ca2+ (Berglund et al., 2003), contradicting the implication that Y1 and possibly Y5 are class B receptors. In addition, several groups have reported that the Y1 receptor internalizes into endosomes, leading to a rapid recycling of the receptor to the cell surface (Fabry et al., 2000; Gicquiaux et al., 2002), a typical feature of class A receptors.
BRET has previously been used to study β-arrestin interaction at the β2-adrenergic receptors (Angers et al., 2000), and recently, BRET2 was used to study β-arrestin interaction at thyrotropin-releasing hormone receptors (Hanyaloglu et al., 2002). In the present study, the binding affinities for agonists were higher than the potency in β-arrestin 2 interaction at all four receptors. Possible explanations could be the constant removal of receptors from the surface due to internalization of receptors followed by decoupling of the receptor-β-arrestin 2 complex, preventing studies of a system at equilibrium when using BRET. Furthermore, overexpression of GFP-β-arrestin 2 compared to the receptors may have changed the stoichiometry of this system and thereby caused the interaction with receptors not normally silenced by β-arrestin 2.
In summary, this study suggests that agonist stimulation of the rhesusY1 and Y5 receptors induces a rapid association of β-arrestin 2 with the receptor. The Y2 receptor responded very slowly and with very low potencies while the Y4 receptors displayed intermediate behavior. These data agree with reports on the rate of Y1 and Y2 internalization and suggest the β-arrestin 2 interaction is an important step in Y1 and Y5 receptor desensitization and internalization. Thus, BRET2 studies using GFP2-tagged β-arrestins can be used to study agonist responses at NPY family receptors and may be a potential screening method for agonists.
Acknowledgments
We acknowledge Dr. Philip Hipskind for synthesis of the Novartis 1 compound.
Footnotes
-
DOI: 10.1124/jpet.103.051227.
-
ABBREVIATIONS: NPY, neuropeptide Y; PYY, peptide YY; PP, pancreatic polypeptide; GPCR, G protein-coupled receptor; BRET, bioluminescence resonance energy transfer; Rluc, Renilla luciferase; GFP, green fluorescent protein; PCR, polymerase chain reaction; HEK, human embryonic kidney; hPP, human pancreatic polypeptide.
- Received March 4, 2003.
- Accepted March 26, 2003.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}