


Abstract
At the spinal level, nociceptin/orphanin FQ (Noc/OFQ) produces pronociceptive and allodynic effects at low doses (picogram range), while causing antinociceptive effects at high doses (microgram range). The discrepancy of pain modulation by Noc/OFQ at low and high doses raised a question whether Noc/OFQ exerted actions through the same Noc/OFQ receptor. In the present study, we examined the involvement of the Noc/OFQ receptor in pain responses with the novel nonpeptide antagonistN-(4-amino-2-methylquinolin-6-yl)-2-(4-ethylphenoxymethyl) benzamide monohydrochloride (JTC-801). Allodynia and hyperalgesia evoked by intrathecal administration of Noc/OFQ (50 pg/mouse) were dose dependently blocked by simultaneous administration of JTC-801 with IC50 values of 32.2 and 363 pg, respectively. JTC-801 did not induce allodynia by itself. Subcutaneous injection of formalin into a hindpaw evoked biphasic pain behaviors such as flinching and biting in mice. Noc/OFQ at 10 pg increased the second-phase pain behaviors evoked by 1% formalin, whereas it strongly inhibited both the first-phase and second-phase pain evoked by 2% formalin at 1 μg. Although the pronociceptive effect by 10 pg of Noc/OFQ was dose dependently blocked by JTC-801 with an IC50 value of 4.58 pg, the antinociceptive effects by 1 μg of Noc/OFQ were not antagonized by JTC-801. Furthermore, both phases of 2% formalin-induced pain behaviors were relieved by JTC-801. These results demonstrate that pronociceptive responses induced by a low dose of Noc/OFQ may be mediated through the Noc/OFQ receptor in the spinal cord and that JTC-801 can be a useful antagonist to examine the involvement of endogenous Noc/OFQ and mediation of the Noc/OFQ receptor under physiological and pathophysiological conditions including pain.
The heptadecapeptide called nociceptin/orphanin FQ (hereafter called Noc/OFQ) was identified as an endogenous ligand of the opioid receptor homolog, designated as ORL1 in humans, and in its rat counterparts as LC132 and ROR-C (Meunier et al., 1995; Reinscheid et al., 1995; Okuda-Ashitaka et al., 1996). Noc/OFQ and its precursor mRNA are found in the superficial dorsal horn of the spinal cord (Okuda-Ashitaka et al., 1996; Ikeda et al., 1998; Neal et al., 1999a;Houtani et al., 2000). The Noc/OFQ receptor is also present in the spinal cord (Ikeda et al., 1998; Neal et al., 1999b; Houtani et al., 2000). In accordance with these localizations, Noc/OFQ is shown to be involved in nociception at the spinal level, but the results in the literature are contradictory, depending on the testing paradigms, animal species, and doses (Meunier, 1997; Ito et al., 2000; Xu et al., 2000). We previously showed that intrathecal (i.t.) administration of Noc/OFQ at very low doses (picogram range) induced allodynia, a state of discomfort and pain evoked by innocuous tactile stimuli, as well as hyperalgesia in conscious mice (Okuda-Ashitaka et al., 1996; Hara et al., 1997). On the other hand, i.t. administration of Noc/OFQ was reported to produce analgesia (Xu et al., 1996), anti-hyperalgesic, and anti-allodynic effects (Hao et al., 1998) at high doses (microgram range). Similarly, whereas it facilitated the second phase of formalin-induced pain at a low dose of 10 pg, Noc/OFQ administered i.t. inhibited both the first-phase and second-phase pain behaviors in the formalin test at high doses of 0.3 to 3 μg/mouse or 2 to 30 μg/rat (Erb et al., 1997; Yamamoto et al., 1997; Nakano et al., 2000). Noc/OFQ was also reported to have no effect on its own (Vanderah et al., 1998). We recently demonstrated that nocistatin, a peptide derived from the same precursor as Noc/OFQ, attenuated allodynia, hyperalgesia, and aggravation of formalin-induced pain by low doses of Noc/OFQ, but it did not affect the inhibitory effect by high doses of Noc/OFQ in the formalin test (Okuda-Ashitaka et al., 1998; Nakano et al., 2000). Some reports showed that antinociceptive effects of Noc/OFQ were sensitive to the opioid antagonist naltrexone (King et al., 1997; Hao and Ogawa, 1998). Therefore, the discrepancy of modulation of pain by low and high doses of Noc/OFQ raised a question whether Noc/OFQ exerted the nociceptive actions through the same Noc/OFQ receptor.
Development of potent and selective antagonists for the Noc/OFQ receptor has enabled us to examine this question. [Phe1ψ(CH2-NH)Gly2]nociceptin(1–13)NH2was reported to behave as a selective antagonist of the Noc/OFQ receptor in isolated tissues (Guerrini et al., 1998), but it turned out to be a full agonist in a cAMP assay of CHO cells transfected with human Noc/OFQ receptor (Butour et al., 1998) and cardiovascular and renal responses in vivo (Kapusta et al., 1999). Recently, an oral active and nonpeptidyl Noc/OFQ receptor antagonist,N-(4-amino-2-methylquinolin-6-yl)-2-(4-ethylphenoxymethyl) benzamide monohydrochloride (JTC-801), has been successfully synthesized (Shinkai et al., 2000). In the present study, we examined the involvement of the Noc/OFQ receptor in pain responses with JTC-801. Some of these data have been presented in abstract form (Muratani et al., 2001).
Materials and Methods
Animals and Intrathecal Injection.
Male ddY mice (Shizuoka Laboratory Center, Hamamatsu, Japan) weighing 20 ± 2 g were used in this study. The animals were housed under conditions of a 12-h light/dark cycle, a constant temperature of 22 ± 2°C, and 60 ± 10% humidity. They were allowed free access to food and water before testing. A 27-gauge stainless steel needle (0.35 mm o.d.) attached to a microsyringe was inserted between the L5 and L6 vertebrae, and 5 μl of drug solution or physiological saline was injected slowly into the subarachnoid space of conscious mice as described previously (Nakano et al., 2000).
Studies on Allodynia.
Studies on allodynia were carried out as described previously (Minami et al., 2000). After i.t. injection of chemicals, each mouse was placed in an individual 13 × 8.5 × 13-cm Plexiglas enclosure with wood chips on the floor and observed. Allodynia was assessed once every 5 min for 50 min by light stroking of the flank of the mice with a paintbrush. The allodynic response was ranked as follows: 0, no response; 1, mild squeaking with attempts to move away from the stroking probe; 2, vigorous squeaking evoked by the stroking probe, biting at the probe, or strong efforts to escape. The maximum possible scores for allodynia of six mice were 2 × 6 = 12 in any 5-min period and 2 × 6 × 10 = 120 for 50 min, and they were taken as 100%. To evaluate the effect of Noc/OFQ antagonists on allodynia, we assessed the effect at the maximal score of allodynia obtained 10 min after i.t. injection of 50 pg of Noc/OFQ.
Studies on Hyperalgesia.
Mice were placed on a hot plate maintained at 55°C, and the elapsed time until the mice showed the first avoidance responses (licking the feet, jumping, or rapidly stamping the paws) was recorded as described previously (Minami et al., 2000). The response time of the mice to the hot plate was measured at 15 min, the point of the maximal hyperalgesic effect obtained with 50 pg of Noc/OFQ.
Formalin Test.
The formalin test was carried out as described previously (Nakano et al., 2000). Approximately 20 min before testing, mice were individually placed in transparent observation chambers for adaptation. The animal was taken out of the chamber, and 20 μl of 1 or 2% formalin in 0.9% saline was injected subcutaneously into the dorsal surface of the right hindpaw. Immediately after the injection, each mouse was returned to the observation chamber. The amount of time spent licking and biting the injected paw was measured with a hand-held stopwatch from 0 to 5 min (the first phase) and from 10 to 30 min (the second phase) after formalin injection and was considered as indicative of pain.
The animals were used only for one experiment. This study was conducted with the approval of the Animal Experimentation Committees of Kansai Medical University and Osaka Medical College, and in accordance with the guidelines of the Ethics Committee of the International Association for the Study of Pain (Zimmermann, 1983).
Chemicals.
Noc/OFQ was obtained from the Peptide Institute (Osaka, Japan). [Phe1ψ(CH2-NH)Gly2]nociceptin(1-13)NH2was supplied by Phoenix Pharmaceuticals (Belmont, CA). Naloxone hydrochloride was purchased from Sigma-Aldrich (St. Louis, MO). JTC-801 was a generous gift from JT Central Pharmacological Research Institute (Osaka, Japan). All chemicals were dissolved in sterile saline on the day of the experiments and kept on ice until used.
Statistics.
Data for allodynia were analyzed by the Mann-Whitney U test between the saline- and Noc/OFQ alone-injected groups. Statistical analysis for the JTC-801-injected group was performed with the Kruskal-Wallis test, followed by Steel's test for multiple comparisons with the Noc/OFQ alone-injected group. Data for hyperalgesia and the formalin test were analyzed byt test for the two-group comparison between the saline- and Noc/OFQ alone-injected groups or by Dunnett's test for multiple comparisons with Noc/OFQ alone-injected groups. P < 0.05 was considered statistically significant. ID50 values with 95% confidence limits (95% CL) were calculated by use of the computer program PRISM.
Results
Effect of the Novel Noc/OFQ Antagonist JTC-801 on Noc/OFQ-Evoked Allodynia.
Before studying the involvement of Noc/OFQ receptor in the Noc/OFQ-induced pain responses, we examined whether [Phe1ψ(CH2-NH)Gly2]nociceptin(1-13)NH2or JTC-801 possessed an agonist activity for induction of allodynia. As reported previously (Hara et al., 1996; Okuda-Ashitaka et al., 1996), i.t. administration of Noc/OFQ (50 pg/mouse) resulted in prominent agitation responses, such as vocalization, biting, and escape from the probe, to tactile stimuli applied to the flank. The Noc/OFQ-induced allodynia was evoked by the first stimulus 5 min after i.t. injection and lasted over the 50-min experimental period (Fig.1A). The i.t. administration of saline did not induce allodynia (data not shown). Whereas mice injected i.t. with JTC-801 showed weak allodynic responses at 5 min but no responses after 15 min, [Phe1ψ(CH2-NH)Gly2]nociceptin(1-13)NH2induced allodynia in a time course similar to that induced by Noc/OFQ. When the allodynic scores obtained for the overall 50-min experimental period were cumulated and expressed as a percentage of the maximum possible score, dose dependence of the Noc/OFQ-induced allodynia showed a bell-shaped pattern from 1 pg to 100 pg, and the maximum effect was observed at 50 pg (Fig. 1B). Whereas JTC-801 negligibly induced allodynia up to 1 ng, dose dependence of [Phe1ψ(CH2-NH)Gly2]nociceptin(1-13)NH2-induced allodynia exhibited a bell-shaped pattern with the maximal effect at 5 pg, one order lower than that of Noc/OFQ (Fig. 1B). These results suggested that [Phe1ψ(CH2-NH)Gly2]nociceptin(1-13)NH2showed a potent agonist activity for induction of allodynia more than Noc/OFQ. Therefore, we used JTC-801 as the Noc/OFQ antagonist for the following experiments.

Effect of i.t. injection of Noc/OFQ and Noc/OFQ receptor antagonists on allodynia. A, time course of allodynia. Mice were injected with 50 pg Noc/OFQ (●), 5 pg [Phe1ψ(CH2-NH)Gly2]nociceptin(1-13)NH2(▵), and 1 pg JTC-801 (○) into the subarachnoid space of conscious mice. B, dose dependence of allodynia. Mice were injected with various doses of Noc/OFQ (●), [Phe1ψ(CH2-NH)Gly2]nociceptin(1-13)NH2(▵), and JTC-801 (○). Allodynia was assessed every 5 min, for 50 min, as described under Materials and Methods. The data (mean ± S.E.M., n = 6) are expressed as a percentage of the maximum possible cumulative score for allodynia evaluated every 5 min (A) and over the 50-min observation period (B).
We first examined the involvement of Noc/OFQ receptor in Noc/OFQ-induced allodynia. When assessed by the maximum value obtained 10 min after Noc/OFQ (50 pg) injection, the allodynia was dose dependently blocked by simultaneous i.t. injection of JTC-801 from 10 pg to 10 ng, with an ID50 value (95% CL) of 32.2 pg (4.01–259 pg) (Fig. 2A).

Effect of i.t. JTC-801 on Noc/OFQ-induced allodynia (A) and hyperalgesia (B). Noc/OFQ (50 pg/mouse) was injected simultaneously with the indicated doses of JTC-801 into the subarachnoid space of conscious mice. A, allodynia was assessed 10 min after i.t. injection as described in the legend for Fig. 1. The allodynic score at 10 min after i.t. injection of Noc/OFQ alone was 83.3% of the maximum possible score and was taken as 100%. The data shown are the mean ± S.E.M. (n = 6). ∗∗,P < 0.01, as compared with the saline-injected group (Mann-Whitney U test). †, P< 0.05, as compared with the Noc/OFQ-injected group (Kruskal Wallis test followed by Steel's test). B, hyperalgesia was assessed 15 min after i.t. injection of Noc/OFQ as described under Materials and Methods. The data shown are the mean ± S.E.M. (n = 10). ∗∗, P < 0.01, as compared with the saline-injected group (t test). ††,P < 0.01, as compared with the Noc/OFQ-injected group (Dunnett's test).
Effect of JTC-801 on Noc/OFQ-Evoked Hyperalgesia.
The thermal hyperalgesia was assessed by the hot-plate (55°C) test. There was no significant difference in the latency period between mice that had not received an i.t. injection (16.4 ± 0.9 s, mean ± S.E.M., n = 10) and those treated with i.t. saline (16.3 ± 0.9 s, at 15 min). Consistent with our previous findings (Hara et al., 1997), i.t. administration of Noc/OFQ (50 pg) shortened the response latency to 9.0 ± 0.6 s at 15 min after i.t. injection, demonstrating that the experimental mice developed increased sensitivity to the thermal stimuli by i.t. Noc/OFQ. The Noc/OFQ-induced hyperalgesia was blocked by JTC-801 with statistical significance at 1 ng and above (Fig. 2B). An ID50 value (95% CL) of JTC-801 was 363 pg (37.8 pg to 3.48 ng), one order higher than that of allodynia. These results demonstrated that both allodynia and hyperalgesia induced by Noc/OFQ were mediated by the Noc/OFQ receptor.
Involvement of Noc/OFQ and Its Receptor in Pain-Like Behaviors in the Formalin Test.
Subcutaneous injection of formalin into the dorsal surface of the hindpaw produced biphasic pain behaviors such as licking and biting, with the first transient phase of 0 to 5 min and the second prolonged phase of 10 to 30 min. We previously showed that Noc/OFQ aggravated the second phase of 1% formalin-induced pain at 10 pg and that the aggravating effect was reversed by 10 pg of nocistatin (Nakano et al., 2000). At 1 μg, Noc/OFQ significantly inhibited both phases of the 2% formalin-induced pain, which were not affected by 1 μg of nocistatin. To clarify the difference in nociceptive effects in the formalin test of Noc/OFQ and nocistatin between low and high doses, here we examined the effects of JTC-801 on the formalin-induced pain. As shown in Fig. 3, Noc/OFQ, at an i.t. dose of 10 pg, given 10 min after subcutaneous injection of 1% formalin, increased the licking and biting time from 23.2 ± 3.3 to 106 ± 10.2 s during the second phase. This aggravation by a low dose of Noc/OFQ was dose dependently reversed by simultaneous administration of JTC-801 from 1 to 100 pg with an IC50 value (95% CL) of 4.58 pg (0.61–50.6 pg).

Effect of i.t. JTC-801 on aggravation by Noc/OFQ of the second-phase pain behavior induced by 1% formalin in mice. The total time spent licking and biting the injected paw was measured during the second phase (10–30 min) after subcutaneous injection of 1% formalin into the hindpaw. Noc/OFQ (10 pg/mouse) was injected simultaneously with indicated doses of JTC-801 10 min after formalin injection. The data shown are the mean ± S.E.M. (n = 8). ∗∗, P < 0.01, as compared with the saline-injected group (t test). ††,P < 0.01, as compared with the Noc/OFQ-injected group (Dunnett's test).
Increase in injection concentration of formalin from 1 to 2% enhanced the licking and biting behaviors from 23.2 ± 3.3 to 99.2 ± 9.9 s during the second phase, comparable with that observed by 10 pg of Noc/OFQ in 1% formalin injection (Figs. 3 and4B). The i.t. injection of JTC-801 significantly decreased the formalin-induced pain behaviors during the second phase at 1 ng and 10 ng. In the saline control group, 2% formalin induced 79.4 ± 7.5 s of licking and biting behaviors during the first phase. JTC-801 administered i.t. significantly decreased the formalin-induced pain behaviors to 38.0 ± 5.3, 42.8 ± 4.7, and 32.2 ± 4.5 s at doses of 0.1, 1, and 10 ng, respectively (Fig. 4A). Consistent with previous studies, a high dose (1 μg/mouse) of Noc/OFQ almost completely suppressed both first and second phases of pain behaviors induced by 2% of formalin in mice (Fig.5). In contrast to the aggravating effect by 10 pg of Noc/OFQ, however, JTC-801 (0.1–10 ng) failed to antagonize the inhibition by 1 μg of Noc/OFQ of either phase of the 2% formalin-induced pain (Fig. 5). Due to the limited solubility of JTC-801, 10 ng was the highest dose employed in this study. Naloxone (1 μg/kg i.t.), a nonspecific opioid antagonist, also could not reverse the inhibition of 1 μg of Noc/OFQ on both phases of the 2% formalin-induced pain (data not shown).

Effect of i.t. JTC-801 on 2% formalin-induced pain in mice. Indicated doses of JTC-801 were injected i.t. 1 min before (A) or 10 min after (B) formalin injection. The total time spent licking and biting the injected paw was measured during the first phase (0–5 min, A) and the second phase (10–30 min, B) after formalin injection. The data shown are the mean ± S.E.M. (n = 8). ∗∗, P < 0.01, as compared with the saline-injected group (Dunnett's test).

Effect of Noc/OFQ and JTC-801 on 2% formalin-induced pain in mice. Noc/OFQ (1 μg/mouse) without or with indicated doses of JTC-801 was simultaneously injected i.t. 1 min before (A) or 10 min after (B) formalin injection. The total time spent licking and biting the injected paw was measured during the first phase (0–5 min, A) and the second phase (10–30 min, B) after formalin injection. The data shown are the mean ± S.E.M. (n = 8). ∗∗,P < 0.01, as compared with the saline-injected group (t test).
Nocistatin (100 pg) given i.t. 1 min before 2% formalin injection decreased the first phase of licking and biting behaviors and when given i.t. 10 min after formalin injection decreased the second-phase behaviors. Simultaneous injection of JTC-801 with nocistatin did not affect the inhibition by nocistatin of either phase of 2% formalin-induced pain (Fig. 6).
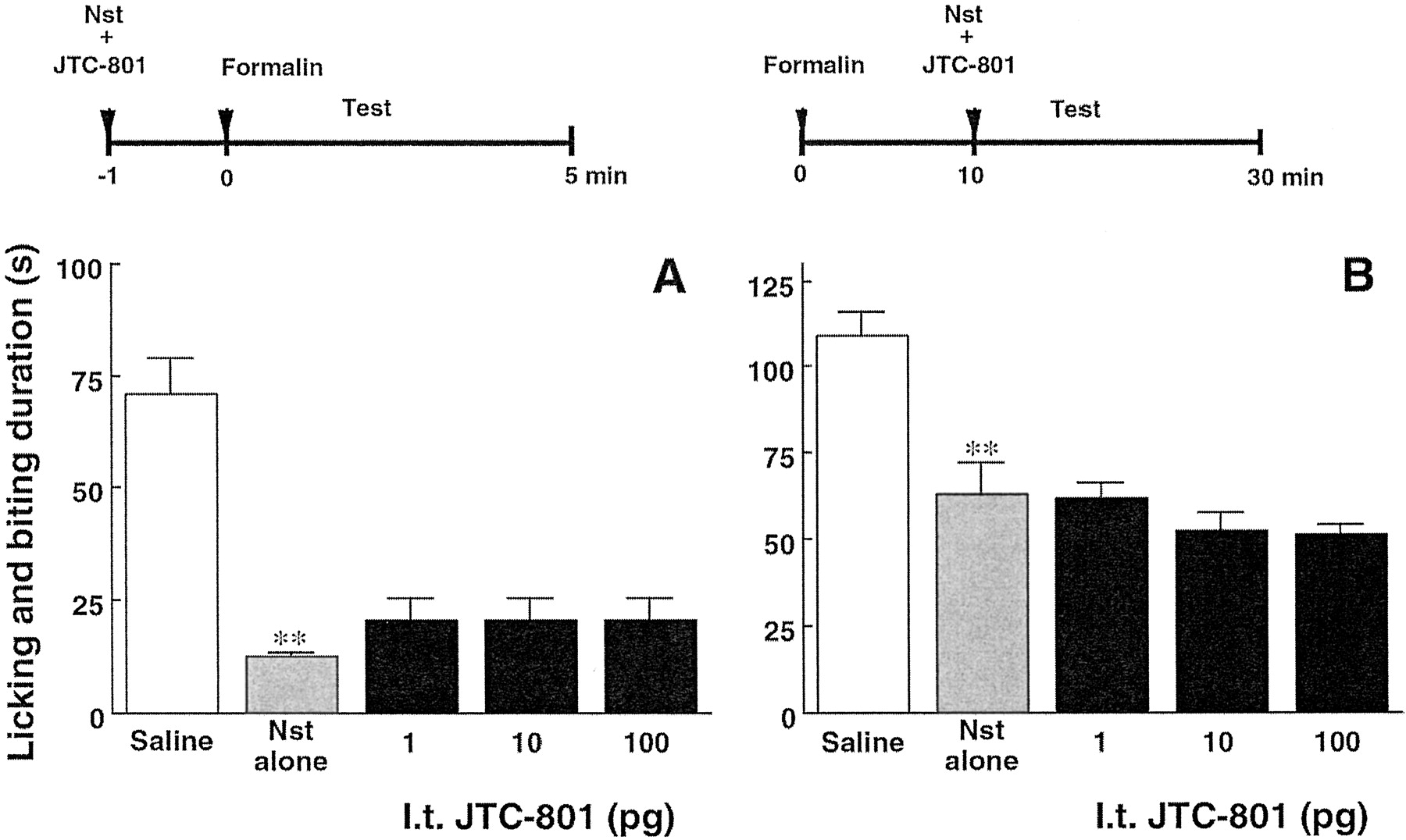
Effect of nocistatin and JTC-801 on 2% formalin-induced pain in mice. A, nocistatin (NST, 100 pg/mouse) without or with indicated doses of JTC-801 was simultaneously injected i.t. 1 min before (A) or 10 min after (B) formalin injection. The total time spent licking and biting the injected paw was measured during the first phase (0–5 min, A) and the second phase (10–30 min, B) after formalin injection. The data shown are the mean ± S.E.M. (n = 8). ∗∗, P < 0.01, as compared with the saline-injected group (t test).
Discussion
A novel nonpeptide Noc/OFQ antagonist JTC-801 was recently synthesized (Shinkai et al., 2000) and shown to inhibit [3H]Noc/OFQ binding to the receptor (Ki = 44.5 nM) and antagonize the suppression of Noc/OFQ on forskolin-induced cAMP formation (IC50 = 2.58 μM) in HeLa cells expressing human Noc/OFQ receptor (Yamada et al., 2002). Whereas [Phe1ψ(CH2-NH)Gly2]nociceptin (1-13)NH2 produced allodynia in a manner similar to that by Noc/OFQ in terms of time course and dose dependence, JTC-801 did not induce allodynia by itself (Fig. 1), confirming that JTC-801 is devoid of an agonist activity in our paradigm. In the present study, we demonstrated that pronociceptive effects by a low dose (10 or 50 pg/mouse) of Noc/OFQ were blocked by JTC-801 with an IC50 of 4.6 to 363 pg, but that the antinociceptive effect by a high dose (1 μg/mouse) of Noc/OFQ was not antagonized by JTC-801 up to 10 ng (Figs. 2-5). In parallel with our studies, the JT Central Pharmacological Research Institute group (Yamada et al., 2002) examined the effects of JTC-801 on the same testing paradigms through different pathways; e.g., orally and intravenously. JTC-801 administered intravenously antagonized the Noc/OFQ-induced allodynia, hyperalgesia, and formalin-evoked pain responses at 10 μg kg−1 and above. JTC-801 administered orally blocked the Noc/OFQ-induced allodynia, hyperalgesia, and formalin-evoked pain responses at ≥30 μg kg−1, ≥1 mg kg−1, and ≥0.3 mg kg−1, respectively. Although Noc/OFQ also elicited a flexor-reflex in picogram doses in the periphery (Inoue et al., 1998), comparison of effective doses of JTC-801 among routes of administration suggested that JTC-801 administered peripherally passed through the blood-brain barrier and exerted analgesic actions in the spinal cord.
Although the spinal effects of Noc/OFQ on pain responses are contradictory, in vivo and in vitro actions of Noc/OFQ have been considered to be inhibitory in the spinal cord (Xu et al., 2000). The effects of Noc/OFQ on pain and analgesia summarized in the review (Meunier, 1997) apparently showed a tendency that, whereas Noc/OFQ administered i.t. causes antinociceptive effects at high doses (nanogram to microgram range), Noc/OFQ produces pronociceptive and allodynic effects at low doses (femtogram to picogram range). Consistent with previous reports in rats (Erb et al., 1997; Yamamoto et al., 1997), the formalin-induced pain behaviors were blocked in mice by 1 μg of Noc/OFQ (Fig. 5). However, we also found that 10 pg of Noc/OFQ facilitated the 1% formalin-evoked pain. Again, the effects of Noc/OFQ on pain-like behaviors are different in the formalin test at low and high doses. Noc/OFQ (10 pg) significantly facilitated the second phase of the 1% formalin-induced pain behaviors in mice, comparable with those induced by 2% formalin (Figs. 3 and 4B). Since Noc/OFQ did not facilitate the second phase of 2% formalin-induced pain behaviors, we previously suggested that Noc/OFQ released in the spinal cord may be involved in the second phase of 2% formalin-induced pain (Nakano et al., 2000). This notion was supported by use of the Noc/OFQ antagonist JTC-801 in the present study. JTC-801 alleviated the second phase of 2% formalin-induced pain behaviors (Fig. 4B) as well as aggravation of 1% formalin-evoked pain by 10 pg of Noc/OFQ (Fig.3). JTC-801 also reduced the first-phase pain responses at 100 pg and above, showing that JTC-801 was 10-fold more potent in suppressing the first phase than the second one in the formalin test (Fig. 4). In this connection, nocistatin was also 10-fold more potent in inhibiting the first phase than the second one (Nakano et al., 2000). It is likely that a small amount of Noc/OFQ released by formalin injection may take part in both phases of pain behaviors through the Noc/OFQ receptor.
Since the dose-response curve for the allodynic effect of Noc/OFQ is bell-shaped (Fig. 1B), one possible explanation for the discrepancy of Noc/OFQ-induced pain responses is that the inhibitory effect of Noc/OFQ at high doses may dominate and override the excitatory effect at low doses (Xu et al., 2000). Whereas 1 μg of Noc/OFQ (approximateMr = 2000)/5 μl (the volume employed for i.t. injection) corresponds to 100 μM, 10 pg of Noc/OFQ/5 μl corresponds to 1 nM. Considering that Noc/OFQ exerts actions through the cloned Noc/OFQ receptor (Kd = 0.1 nM) (Meunier et al., 1995; Reinscheid et al., 1995), a high dose of Noc/OFQ may exert actions through a receptor different from the cloned Noc/OFQ receptor (Ito et al., 2000). This idea was also suggested by the observations that nocistatin antagonized the pronociceptive effects by a low dose of Noc/OFQ, but not analgesic effects by a high dose of Noc/OFQ (Okuda-Ashitaka et al., 1998; Nakano et al., 2000). In fact, inhibition by 1 μg of Noc/OFQ of the 2% formalin-induced pain behaviors was not affected by JTC-801 (Fig. 5). J-113397, another nonpeptidyl Noc/OFQ antagonist, was shown to enhance the second phase of formalin-induced pain behaviors at 3 to 30 μg/10 μl when injected i.t. or intracerebroventricularly (Yamamoto et al., 2001). The concentration employed in the study was 3 to 30 μg/10 μl, corresponding to approximately 1 to 10 mM, the concentrations of which were extremely higher than that (IC50 = 2.3 nM) for the displacement of [125I]Noc/OFQ binding to the cloned human Noc/OFQ receptor (Kawamoto et al., 1999). Although anti-nociceptive effects of Noc/OFQ were shown to be sensitive to the opioid antagonist naltrexone (King et al., 1997; Hao and Ogawa, 1998), the inhibitory effect of a high dose of Noc/OFQ was not antagonized by naloxone in the present study. Interestingly, it was previously reported that Noc/OFQ administered intracerebroventricularly induced both hyperalgesia and analgesia, and that blockade of the antiopioid ς system by haloperidol enhanced the analgesic potency of Noc/OFQ (Rossi et al., 1997). Furthermore, the Noc/OFQ derivative Noc/OFQ(1–11) lost the hyperalgesic activity while retaining analgesic activity. Since Noc/OFQ(1–11) with the 1000-fold lower affinity against [125I]Tyr14-Noc/OFQ binding in mouse brain and transfected cell lines as compared with Noc/OFQ, inhibited forskolin-stimulated cAMP formation in mouse brain with a potency similar to Noc/OFQ, heterogeneity of Noc/OFQ receptors was suggested in mouse brain (Mathis et al., 1997). A mechanism for the anti-nociceptive effect by a high dose of Noc/OFQ remains to be elucidated.
In summary, JTC-801 is a selective small-molecule Noc/OFQ antagonist with good bioavailability and blood-brain barrier permeability (Shinkai et al., 2000). JTC-801 administered i.t. blocked allodynia, hyperalgesia and alleviation of formalin-evoked pain behaviors induced by a low dose of Noc/OFQ, but not the analgesic effect in the formalin test by a high dose of Noc/OFQ. These results demonstrated that pro-nociceptive responses induced by a low dose of Noc/OFQ may be mediated through the Noc/OFQ receptor in the spinal cord and that the anti-nociceptive effect by a high dose of Noc/OFQ in the formalin test may be mediated by an unknown mechanism. JTC-801 can be a good tool to clarify whether or not Noc/OFQ exhibits actions through the cloned receptor and to examine the involvement of endogenous Noc/OFQ in pathophysiological conditions including pain.
Footnotes
-
This work was supported in part by Grants-in-Aids for Scientific Research on Priority Areas, Scientific Research (B), and Encouragement of Young Scientists from the Ministry of Education, Culture, Sports, Science, and Technology of Japan and by grants from the Science Research Promotion Fund of the Japan Private School Promotion Foundation and Jinsenkai Foundation of Osaka Medical College.
-
DOI: 10.1124/jpet.102.036095
- Abbreviation:
- Noc/OFQ
- nociceptin/orphanin FQ
- JTC-801
- N-(4-amino-2-methylquinolin-6-yl)-2-(4-ethylphenoxymethyl) benzamide monohydrochloride
- J-113397
- 1-[(3R,4R)-1-cyclooctylmethyl-3-hydroxymethyl-4-piperidyl]-3-ethyl-1,3-dihydro-2H-benzimidazol-2-one
- Received March 13, 2002.
- Accepted June 25, 2002.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}