


Abstract
The serotonin2C (5-HT2C) receptor couples to multiple effector mechanisms, including phospholipase A2-mediated arachidonic acid (AA) release and phospholipase C-mediated production of inositol phosphates (IP). Agonist relative efficacy differs depending upon which response (AA release or IP accumulation) is measured. In this study, we investigated the characteristics and agonist dependence of rapid desensitization of 5-HT2C receptor-mediated AA release and IP accumulation measured simultaneously from the same cell population. Pretreatment with 5-HT reduced the ability of a maximal concentration of 5-HT to elicit AA release and IP accumulation by about 60%; however, the AA response desensitized more rapidly (t1/2 = 1.3 min) than the IP response (t1/2 = 6.9 min). In addition, desensitization of the IP response was more sensitive (occurred at lower receptor occupancy levels) than the AA response. Moreover, in response to submaximal 5-HT concentrations, after an initial transient desensitization, the AA response was enhanced by up to ∼250%. After maximal desensitization, both responses recovered, but recovery of the AA response was complete and faster than that for IP. Desensitization of both responses was also agonist-dependent, and the capacity of agonists to elicit desensitization was not related to their efficacy to activate signaling. These data suggest that desensitization of the 5-HT2C receptor system is both agonist- and effector pathway-dependent and underscore the need to study multiple cellular responses to multiple agonists to understand receptor-mediated signaling systems.
The serotonin2C (5-HT2C) receptor is a member of the 5-HT2 family of seven transmembrane-spanning (7-TMS) receptors. This receptor couples to phospholipase C (PLC)-mediated phosphatidylinositol (PI) hydrolysis and phospholipase A2(PLA2)-mediated arachidonic acid (AA) release in a pertussis toxin-insensitive manner (Berg et al., 1998). There is considerable evidence that 5-HT2C receptors play important roles in many physiological functions and behaviors, such as sleep, affective state, feeding behavior, and temperature regulation (Zifa and Fillion, 1992; Boess and Martin, 1994; Hoyer et al., 1994). Furthermore, this receptor may be a target for the therapeutic action of the atypical antipsychotic drugs (Herrick-Davis et al., 2000; Rauser et al., 2001) and for hallucinogenic drugs of abuse (Glennon, 1994;Sanders-Bush, 1994). Because the 5-HT2C receptor is a major focus of drug discovery efforts, it is important to develop an understanding of the ways by which the responsiveness of this receptor system can be regulated.
It is well known that continued agonist exposure can reduce the responsiveness of many 7-TMS receptor systems; a process known as desensitization (for reviews, see Freedman and Lefkowitz, 1996;Ferguson, 2001). Rapid (within minutes) desensitization occurs through a variety of mechanisms (frequently involving phosphorylation) that can target the receptor, the transducing molecules (e.g., G protein), and the effectors. Because the ability of a drug to produce a response (efficacy) may be limited by time-dependent desensitization, study of the mechanisms and regulation of desensitization are currently important questions in pharmacology.
It is now generally accepted that most, if not all, 7-TMS receptors can couple to multiple effector pathways through one or more heterotrimeric G proteins or other transducer molecules (e.g., arrestins, small G proteins, and PDZ domain-containing proteins). However, most studies of 7-TMS receptor-mediated desensitization have examined loss of responsiveness of a single pathway only. For example, although many Gq family-coupled receptor subtypes (e.g., 5-HT2A/2C, muscarinic M1,3,5, and α1-adrenergic) known to couple to PLC-PI hydrolysis also couple to PLA2-AA release (Richelson, 1995; Berg et al., 1998; Zhong and Minneman, 1999), typically only changes in the responsiveness of the PLC pathway [with measures of inositol phosphate (IP) accumulation or intracellular calcium levels] have been used to assess desensitization. Because desensitization mechanisms can target specific transducer/effector molecules in addition to the receptor (Ali et al., 1997), characteristics of 7-TMS receptor system desensitization could be response-dependent. Such effector pathway-dependent desensitization would lead to qualitative, as well as quantitative, changes in response to receptor activation over time. Consequently, it is important to study desensitization of each of multiple responses coupled to a receptor.
Recent studies have shown that agonist-relative efficacy is effector pathway-dependent (Berg et al., 1998; Brink et al., 2000; Cordeaux et al., 2000; Watson et al., 2000; Berg et al., 2001a). For example, when expressed in CHO cells, the 5-HT2C and 5-HT2A receptors can couple independently to PLC and PLA2 effector pathways and agonist relative efficacy differs depending upon whether PI hydrolysis or arachidonic acid release is measured (Berg et al., 1998). Because some desensitization mechanisms can be viewed as additional independent effector pathways (e.g., G protein receptor kinase activation), it is possible that the relative efficacy of agonists to elicit desensitization may be different from that for activation of other responses.
In this study, we investigated agonist-mediated, rapid desensitization of PLC-PI hydrolysis and PLA2-AA release responses coupled to the human 5-HT2C receptor stably expressed in CHO cells. To avoid confounds of data interpretation that can occur when responses are measured under different experimental conditions, both IP accumulation and AA release were measured from the same cells, simultaneously. We found that characteristics of desensitization elicited by 5-HT were effector pathway-dependent. Furthermore, the relative efficacy of agonists to promote desensitization also differed and did not seem to be linked with their efficacy to activate either PLC or PLA2.
Materials and Methods
Materials.
The following materials were purchased from commercial sources: [3H]myo-inositol and [3H]arachidonic acid (PerkinElmer Life Sciences, Boston, MA); 5-HT HCl, lysergic acid diethylamide (LSD), bufotenin, quipazine, 1-(3-chlorophenyl)piperazine (mCPP), and 3-trifluotomethylphenyl-piperazine (TFMPP) (Sigma/RBI, Natick, MA); and fetal bovine serum (Gemini Bioproducts, Calabasas, CA). All other tissue culture reagents were purchased from Invitrogen (Carlsbad, CA). All other drugs and chemicals (reagent grade) were purchased from Sigma-Aldrich (St. Louis, MO).
Cell Culture.
CHO-1C19 cells are CHO-K1-derived cell lines that stably express human 5-HT2C receptors at a density of ∼200 fmol/mg protein (Berg et al., 1994). Cells were maintained in α-minimal essential medium supplemented with 5% fetal bovine serum and 300 μg/ml hygromyocin. For these experiments, the cells were seeded into multiwell tissue culture plates at a density of 4 × 104 cells/cm2. After a 24-h plating period, cells were washed with Hanks' balanced salt solution and placed into Dulbecco's modified Eagle's medium/F-12 (1:1) with 5 μg/ml insulin, 5 μg/ml transferrin, 30 nM selenium, 20 nM progesterone, and 100 μM putrescine (serum-free media). Cells were grown in serum-free media for 24 h before experimentation.
IP Accumulation and AA Release Measurements.
Cells were labeled with 1 μCi/ml [3H]myo-inositol in serum-free medium for 24 h and 0.1 μCi/ml [3H]AA for 4 h. Total IP accumulation (IP1, IP2, and IP3, collectively referred to as IP) and AA release were measured as described previously (Berg et al., 1998). Measurements of PLC-mediated IP accumulation and PLA2-mediated AA release were made from the same multiwell, simultaneously. Desensitization studies were conducted essentially as described previously (Berg et al., 2001b) with the exception of a brief wash period between pretreatment and the responsiveness test. To elicit desensitization, cells were exposed to agonist (in the absence of LiCl) for various periods of time (0- to 60-min pretreatment time) in the presence of the radiolabels. After this agonist exposure, cells were washed (three times) rapidly (∼45 s) with Hanks' balanced salt solution containing calcium and magnesium, 20 mM HEPES, and 0.1% bovine serum albumin. Responsiveness of the receptor system was assessed by measuring IP accumulation and AA release in response to application of a maximal concentration of 5-HT (10 μM) for 10 min in the presence of 20 mM LiCl and 0.1% bovine serum albumin. After the 10-min incubation, a 200-μl aliquot of media from each well was added directly to scintillation vials for measurement of 3H content (AA release) with liquid scintillation counting. The remaining media were aspirated quickly and 2 ml of 10 mM formic acid was added to extract the accumulated [3H]IP. The [3H]IP were separated with ion exchange chromatography and quantified with liquid scintillation counting.
Data Analysis.
For desensitization experiments, IP accumulation and AA release data were expressed as a percentage of 5-HT-stimulated values, according to the following equation.
To calculate fractional occupancy the following equation was used.
Time-course data were fit to the following equation using nonlinear regression analysis.
Statistical Analysis.
Analysis of variance was used for statistical comparisons, and the Newman-Keuls post hoc test was used to determine which groups were different. A p value <0.05 was considered significant. Data shown represent mean ± S.E.M. of several independent experiments.
Results
Effector Pathway-Dependent Desensitization and Recovery.
The effect of pretreatment (0–60 min) with 5-HT (10 μM; 99.7% occupancy) on 5-HT-mediated IP accumulation and AA release, measured simultaneously from the same cells, is shown in Fig.1. Both responses desensitized partially to a similar maximal extent, however, the rate of desensitization of the AA response was faster than that for IP accumulation. TheDmax was 62 ± 2 and 61 ± 3% of control for IP accumulation and AA release, respectively, and the Kdes was 0.10 ± 0.02 (t1/2 = 6.9 min) and 0.52 ± 0.08 min−1 (t1/2 = 1.3 min) for IP accumulation and AA release, respectively (mean ± S.E.M., n = 3–6, p < 0.05).

Time course of desensitization of 5-HT2Creceptor-mediated IP accumulation and AA release in response to pretreatment with 5-HT. Cells were pretreated for 0 to 60 min with 10 μM 5-HT (99.7% occupancy) at 37°C. The cells were then washed (three times) rapidly (45 s) and incubated for 10 min with 10 μM 5-HT. AA release and IP accumulation were measured simultaneously from the same multiwell, with the exception of the early time points (1–3 min) in which only AA release was measured. Data, expressed as a percentage of control 5-HT stimulation in the absence of pretreatment, represent the mean ± S.E.M. of three to six experiments. 5-HT-stimulated activity was 969 ± 106 and 553 ± 70% above basal for IP accumulation and AA release, respectively. The AA and IP responses that remained after 60-min pretreatment with 5-HT were blocked by the antagonist mianserin (10 μM). For the AA response, data points after 2 min of agonist pretreatment were not significantly different. Gray line at 100% represents 5-HT response in the absence of pretreatment.
To examine the occupancy dependence of 5-HT-mediated desensitization of IP accumulation versus that of AA release, cells were pretreated for 0 to 60 min with 10 nM (25% occupancy), 30 nM (50% occupancy), or 100 nM (75% occupancy) 5-HT before the responsiveness test. As shown in Fig. 2, pretreatment with 30 and 100 nM concentrations of 5-HT induced desensitization of the IP response (18 ± 5 and 28 ± 1%, 30 versus 100 nM 5-HT, respectively) that was less than the desensitization induced by a maximal concentration of 5-HT (Fig. 1). The IP response did not desensitize when cells were pretreated with 10 nM 5-HT for up to 1 h. In contrast to the IP response, the AA response was not as sensitive to desensitization by lower concentrations of 5-HT. In fact, with pretreatment with 100 nM 5-HT, a modest (44 ± 14%;p < 0.05) and transient (within 5 min) desensitization occurred, which was followed by sensitization of the AA response to 145 ± 28% of control (p < 0.05). Pretreatment with lower concentrations of 5-HT produced only a time-dependent sensitization of the AA response. After exposure to 10 nM 5-HT (25% occupancy), the subsequent response to the 5-HT test stimulus was 238 ± 24% of control (p < 0.05).

Concentration dependence of desensitization of 5-HT2C receptor-mediated IP accumulation and AA release. Cells were pretreated for 0 to 60 min with indicated concentrations of 5-HT at 37°C, washed, and AA release and IP accumulation were measured in response to 10 μM 5-HT (10 min). Values for receptor occupancy (θ) at each concentration are provided in the legend. Data are expressed as a percentage of control 5-HT stimulation and represent mean ± S.E.M. of four to six experiments. 5-HT-stimulated activity was 975 ± 65 and 479 ± 94% above basal IP accumulation and AA release, respectively. Gray line at 100% represents 5-HT response in the absence of pretreatment. Data for the curves for 5-HT (10 μM) are from Fig. 1.
Figure 3 shows the time course for recovery of responsiveness of the 5-HT2C receptor system to stimulation with 5-HT after maximal desensitization by pretreatment with 5-HT (10 μM) for 15 min. Both AA and IP responses recovered partially within 15 min of agonist removal, however, by 60 min only the AA response had recovered fully. 5-HT-stimulated IP accumulation was only about 70% of control after 60 min of recovery.

Recovery of 5-HT2C receptor-mediated responsiveness after agonist washout. Cells were pretreated for 15 min with 10 μM 5-HT, washed, and incubated for 0 to 60 min in the absence of 5-HT. After the recovery period, IP accumulation and AA release were measured in response to 10 μM 5-HT for 10 min. Data, expressed as a percentage of control 5-HT stimulation, represent mean ± S.E.M. of six experiments. 5-HT-stimulated activity was 324 ± 32 and 241 ± 26% above basal for IP accumulation and AA release, respectively. Gray line at 100% represents the control 5-HT response in the absence of pretreatment. ∗, p < 0.05 compared with nondesensitized control; †, p < 0.05 compared with desensitized, no recovery point.
Agonist-Dependent Desensitization.
As illustrated in Fig.4 and Table1, the capacity of different agonists to elicit changes in responsiveness of the IP and AA responses varied widely, in terms of magnitude and time course. Pretreatment with agonists at concentrations that produced greater than 95% occupancy (at least 20 times the KA) reduced the responsiveness of the 5-HT2C-IP pathway. Each of the agonists produced desensitization of the IP response but to different magnitudes and with different rates. The relative efficacy to elicit desensitization of the IP response was not clearly related to the agonist's relative efficacy to stimulate IP accumulation or AA release (Fig. 5; Table1). For example, quipazine, a strong agonist for 5-HT2C-stimulated IP accumulation (0.9) and a moderate agonist for stimulation of AA release (0.6), caused desensitization of the IP response of about 60% that of 5-HT and sensitization of the AA response. Bufotenin, a relatively strong agonist for IP accumulation (0.8) and a full agonist for AA release (1) relative to 5-HT, produced desensitization of the IP response that was about 50% that of 5-HT and did not alter responsiveness of the 5-HT2C-AA pathway. The time course to produce changes in responsiveness also varied. For example, TFMPP produced very rapid desensitization of the IP response (more rapid than 5-HT), whereas the rate of desensitization elicited by quipazine, an agonist with similar relative efficacy as TFMPP for IP accumulation, was much slower than that of 5-HT. For AA release, most agonists seemed to produce a biphasic response characterized by an initial, transient desensitization followed by return of responsiveness or sensitization. The change in responsiveness for 5-HT-stimulated AA release was also not related to agonist relative efficacy for IP accumulation or AA release. Although quipazine and TFMPP have equal relative efficacy to stimulate AA release (0.6) and similar relative efficacy for IP accumulation (0.9 versus 1), TFMPP produced strong desensitization of AA release at 60 min of pretreatment, whereas quipazine produced sensitization.
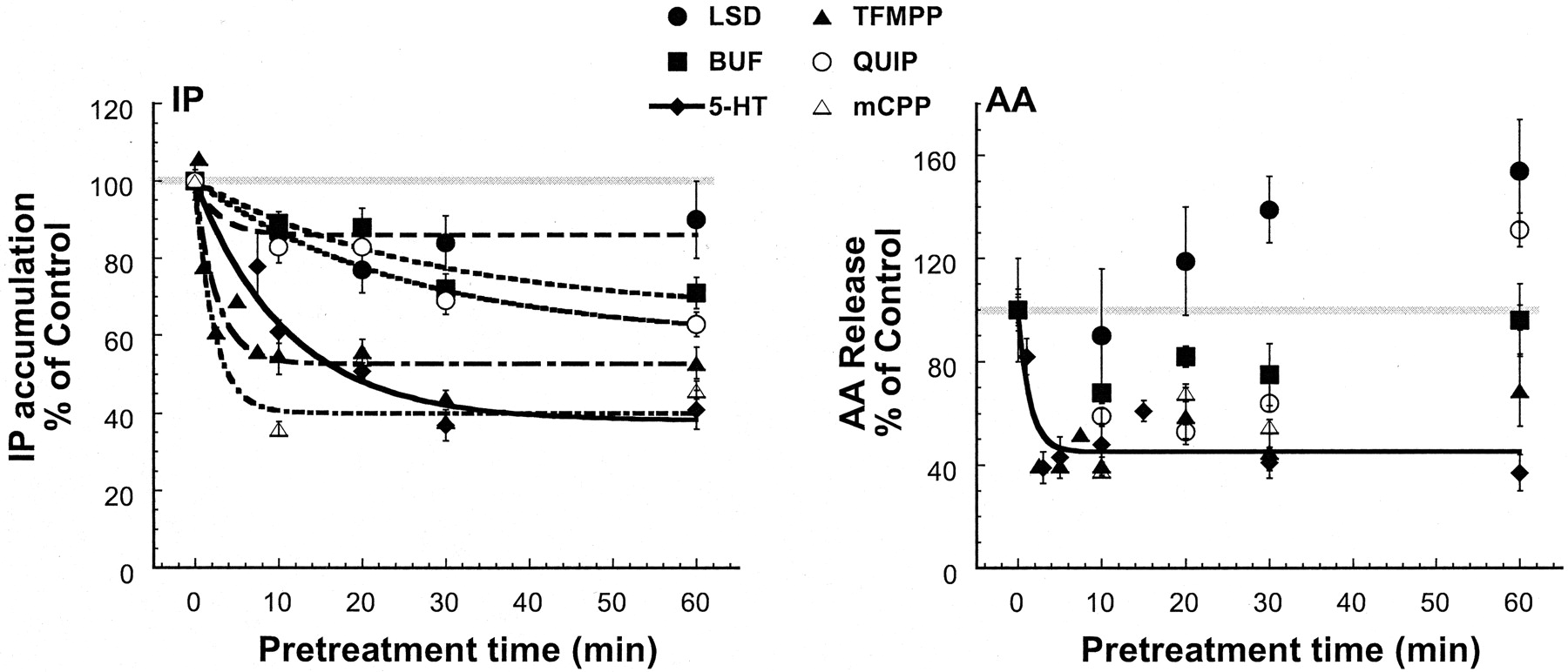
Agonist dependence of desensitization of 5-HT2C receptor-mediated IP accumulation and AA release. Cells were pretreated for 0 to 60 min with maximal concentrations (at least 20 times the KA) of the agonists 5-HT (10 μM), quipazine (30 μM), LSD (100 nM), TFMPP (10 μM), mCPP (20 μM), or bufotenin (10 μM), washed, and AA release and IP accumulation were measured in response to 10 μM 5-HT for 10 min. Data, expressed as a percentage of control 5-HT stimulation, represent the mean ± S.E.M. of three to six experiments. Control 5-HT-stimulated activity was 976 ± 48 and 487 ± 59% above basal for IP accumulation and AA release, respectively.
Agonist relative efficacy for 5-HT2C receptor-mediated stimulation and desensitization of IP accumulation and AA release

Relative efficacies of agonists for stimulation and desensitization of AA release and IP accumulation. Relative efficacy values are with reference to the effect produced by maximal 5-HT. For stimulation of AA release and IP accumulation, values are from Berg et al. (1998), with the exception of mCPP. Values for desensitization indicate that pretreatment with the ligand resulted in enhancement of 5-HT-stimulated AA release.
Discussion
As with many other receptors, the 5-HT2Creceptor couples to more than one cellular signaling pathway. The most well known and best-studied effector pathway activated by 5-HT2C receptors is the PLC-PI hydrolysis pathway. Receptor-mediated activation of this pathway occurs via pertussis toxin-insensitive G proteins and leads to hydrolysis of inositol-containing phospholipids, resulting in the production of inositol phosphates (e.g., IP3) and diacylglycerol. IP3 acts to increase levels of intracellular calcium, whereas diacylglycerol is an endogenous activator of protein kinase C. A second effector pathway coupled to the 5-HT2C receptor is the PLA2-AA pathway. In CHO cells, activation of PLA2 is also pertussis toxin insensitive, is independent of PLC-PI hydrolysis, and results in liberation of AA from various membrane phospholipids (Berg et al., 1998). AA has many cellular actions of its own and is also metabolized by a variety of enzymes to a myriad of bioactive compounds (eicosanoids) such as prostaglandins, thromboxanes, and leukotrienes.
Previous studies of 5-HT2C receptor system desensitization have focused exclusively on the PLC-IP pathway, which desensitizes rapidly, within 10 to 20 min, in SH-SY5Y, NIH/3T3, and CHO cells (Westphal et al., 1995; Briddon et al., 1998; Berg et al., 2001a,b; Porter et al., 2001), and the results presented herein agree with these earlier reports. However, there are no reports that describe desensitization of the PLA2 response coupled to the 5-HT2C receptor. In this study we measured changes in the responsiveness of both PLC and PLA2 effector pathways, simultaneously from the same cells, under identical conditions. We found that although both responses desensitized to a similar maximal extent (∼60%) in response to pretreatment with a maximal concentration of 5-HT (10 μM), there were clear effector pathway-dependent differences in the characteristics of desensitization in response to 5-HT pretreatment. We found that the rate of desensitization of the AA response (t1/2 = 1.3 min) was faster than that for IP accumulation (t1/2 = 6.9 min). This is in agreement with earlier reports that desensitization of the AA response occurred faster than the IP response for agonist activation of gonadotropin-releasing hormone and bradykinin B1 receptors (Poulin et al., 1998; Zhou et al., 2000). In contrast to the desensitization response to pretreatment with a maximal 5-HT concentration, the AA response was not as sensitive as the IP response to desensitization with lower concentrations of 5-HT. Submaximal concentrations of 5-HT desensitized the IP response but produced a biphasic change in responsiveness for the AA pathway, consisting of an initial transient desensitization followed by enhanced responsiveness. Finally, recovery of responsiveness of the IP pathway was slower than that for AA release. Taken together, these data demonstrate that desensitization of the 5-HT2C receptor system is effector pathway-dependent.
Effector pathway-dependent desensitization suggests that the qualitative, as well as the quantitative, response of a cell to 5-HT can change with time. For example, after exposure of a cell to a submaximal concentration of 5-HT that could produce desensitization of the PLC pathway but sensitize the PLA2 pathway, the cellular response to 5-HT could change from one that is dominated by PLC to one in which AA signaling cascade predominates. With the assumption that the PLC-IP and PLA2-AA pathways regulate different aspects of cell physiology, one would expect that such differential effector pathway changes in responsiveness would result in marked, time-dependent differences in the quality of response to 5-HT between a naive cell and a cell that has had prior exposure to 5-HT.
Several studies have reported that agonist efficacy to produce desensitization is associated with their efficacy to activate a signal transduction pathway (January et al., 1997; Kovoor et al., 1998;Oppermann et al., 1999); however, there are reports to the contrary (Yu et al., 1997; Lewis et al., 1998). For example, there was a strong correlation between the efficacy of β-adrenergic receptor agonists to activate adenylyl cyclase with their ability to desensitize the β-adrenergic receptor-adenylyl cyclase pathway and to internalize and phosphorylate the receptor (January et al., 1997). On the other hand,Yu et al. (1997) found that methadone and l-α-acetyl-methadone were more efficacious at causing phosphorylation and producing desensitization of μ-opioid receptors than the stronger agonist morphine, and Lewis et al. (1998) reported that dopamine D1A agonist efficacy to stimulate adenylyl cyclase activity was not a good predictor of their capacity to desensitize that system. Herein, we found that desensitization elicited by pretreatment with different agonists also did not seem to be related to the relative efficacy of the agonist to activate either the PLC-IP or the PLA2-AA pathways. For example, the magnitude of desensitization of the AA and IP responses elicited by TFMPP, an agonist with relative efficacy values of 1 and 0.6 for PLC and PLA2, respectively, was similar to that produced by 5-HT. However, desensitization elicited by quipazine, a drug with relative efficacy for PLC and PLA2 similar to that of TFMPP (0.9 and 0.6, respectively) was markedly different from that produced by TFMPP. Pretreatment with quipazine produced less desensitization of the PLC response and enhanced the responsiveness of the PLA2 pathway. Bufotenin, an agonist with efficacies similar to that of 5-HT to activate PLC and PLA2 (0.8 and 1, respectively), produced much less desensitization of both responses than did 5-HT.
The differences between the relative efficacies of agonists to produce desensitization and those for activation of signaling pathways for the 5-HT2C receptor system is not surprising given that the relative efficacies of agonists to activate the PLC-IP and the PLA2-AA pathways themselves differ substantially (Berg et al., 1998). This effector pathway dependence of relative efficacy has been postulated to be due to ligand-specific receptor conformations that have differential capacity to couple to/activate the individual signaling systems (Kenakin, 1995; Clarke and Bond, 1998). Perhaps ligand-specific receptor conformations also differ in their capacity to activate desensitization mechanisms. In this regard, the phenomenon termed “agonist-directed trafficking of receptor stimulus” may be extended to include desensitization as well as activation of signaling responses.
Alternatively, ligand-specific receptor conformations could serve differentially as targets for desensitization mechanisms that target the receptor. It is known that some desensitization mechanisms, such as G protein receptor kinase and arrestin, seem to be sensitive to receptor conformation because they preferentially interact with agonist-activated receptors (Benovic et al., 1986; Wu et al., 1997). Thus, it may be possible that certain agonists can produce a receptor conformation(s) that is efficiently targeted by desensitization mechanisms (high efficacy) but that is less efficient at activating signal-transducing mechanisms (low efficacy). Consistent with this hypothesis are the data of Thomas et al. (2000) where an angiotensin analog, Sar1,Ile4,Ile8-AngII, was inactive at signaling via the AT1Areceptor-PLC pathway but elicited receptor phosphorylation equivalent to that of a full agonist.
Agonist-dependent desensitization of the 5-HT2Creceptor system could have interesting implications in vivo. Consideration of not only efficacy for activation of individual signaling systems but also for their desensitization may be important in the choice of a drug to activate the 5-HT2Creceptor system. For example, bufotenin, which is a full agonist on the AA pathway and a strong partial agonist for the IP pathway (0.8), weakly desensitized both responses. Consequently, bufotenin may be a better agonist to sustain IP and AA responses over time compared with either 5-HT itself or TFMPP, which produce rapid and strong desensitization of both responses. LSD and quipazine also present interesting possibilities, because they sensitize the AA response with some desensitization of the IP response. Thus, after exposure to LSD or quipazine, PLA2-dependent physiological responses might be enhanced, whereas PLC-dependent physiological changes might be reduced. Thus, a complete understanding of agonist action must include temporal and effector pathway-dependent changes in efficacy.
Acknowledgments
We thank Blythe King and Jodie Cropper for expert technical assistance.
Footnotes
-
This work was supported by U.S. Public Health Service Grants DA 09094 (to K.A.B.) and GM 58652 and the Texas Advanced Research Program (3569-0044; to W.P.C. and K.A.B.).
- Abbreviations:
- 5-HT
- serotonin
- 7-TMS
- seven transmembrane spanning
- PLC
- phospholipase C
- PI
- phosphatidylinositol
- PLA2
- phospholipase A2
- AA
- arachidonic acid
- IP
- inositol phosphates
- CHO
- Chinese hamster ovary
- LSD
- lysergic acid diethylamide
- mCPP
- 1-(3-chlorophenyl)piperazine
- TFMPP
- 3-trifluotomethylphenyl-piperazine
- Received February 5, 2002.
- Accepted March 25, 2002.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}