


Abstract
Recent studies have established the functional and molecular presence of a high-affinity peptide transporter, PEPT2, in whole tissue rat choroid plexus. However, the precise membrane location and directionality of PEPT2-mediated transport is uncertain at present. In this study, we examined the transport kinetics of a model dipeptide, glycylsarcosine (GlySar), along with the protein expression of PEPT2 using primary cell cultures of choroidal epithelium from neonatal rats. GlySar accumulation and transepithelial transport were 3 to 4 times higher when introduced from the apical as opposed to the basal side of the monolayers. GlySar apical uptake was also stimulated by an inwardly directed proton gradient. The uptake of GlySar was inhibited by di/tripeptides, carnosine, and α-amino cephalosporins but was unaffected by amino acids, cephalosporins lacking an α-amino group, and organic anions and cations. The Michaelis constant (Km) of GlySar was 59.6 μM for apical uptake and 1.4 mM for basal uptake; this is consistent with the high-affinity properties of PEPT2 at the apical membrane. Immunoblot analyses and immunofluorescent confocal microscopy demonstrated the presence of PEPT2, but not PEPT1, in rat choroid plexus epithelial cells. Moreover, PEPT2 was present in the apical and subapical regions of the cell but was absent in the basolateral membrane. These findings demonstrate, for the first time, that PEPT2 protein is present at the apical membrane of choroidal epithelial cells and that it is functionally active at this membrane surface. The results suggest that PEPT2 may have a role in the efflux of peptides and/or mimetics from cerebrospinal fluid to the blood.
Proton-coupled peptide transporters, PEPT1 and PEPT2, mediate the uptake of di- and tripeptides via an electrogenic transport system driven by the presence of an inwardly directed proton gradient and negative membrane potential (Fei et al., 1994; Liu et al., 1995; Nussberger and Hediger, 1995;Leibach and Ganapathy, 1996). PEPT1, a low-affinity and high-capacity transporter, is mainly expressed in the small intestine and at low levels in the proximal tubule of the kidney (Terada et al., 1997; Shen et al., 1999). PEPT2, a high-affinity and low-capacity transporter, is mainly expressed in the kidney (Shen et al., 1999). These transporters serve important physiological and pharmacological functions since small peptides and peptidomimetic drugs are absorbed in the intestine or reabsorbed in the kidney via these carriers. More recently, two peptide/histidine transporters, PHT1 (Yamashita et al., 1997) and PHT2 (Sakata et al., 2001), were cloned from rat brain and shown to transport histidine and small peptides with high affinity and in a proton gradient-dependent manner. Whereas PHT1 is expressed strongly in the brain and eye, PHT2 is expressed primarily in the lymphatic system and detected faintly in the brain. PHT1 and PHT2 share an amino acid identity of 49%, but homology to either rat PEPT1 or PEPT2 is less than 25%. More importantly, their physiological role in the brain has yet to be elucidated.
In contrast to the intestine and kidney, little is known about the cellular and molecular mechanisms of peptide and peptidomimetic transport between the blood and brain or cerebrospinal fluid (CSF). The brain is an unusual organ in that it is protected from circulating drugs, toxins, and xenobiotics by tight junctions in the blood-brain barrier (i.e., the cerebral capillary endothelial cells) and blood-CSF barrier (i.e., the choroid plexus epithelial cells). As a result, the paracellular transport of hydrophilic agents into the brain is restricted. Similar to other epithelia, choroid plexus cells are polar with distinct apical (CSF-facing) and basolateral (blood-facing) surfaces. Each polar membrane has distinct characteristics as well as transporters that are uniquely distributed between the two surfaces. These transporters may play an important role in allowing blood-to-brain influx or brain-to-blood efflux of nutrients, neurotransmitter metabolites, and various neuroactive drugs.
The presence of peptide transporters within the brain has generated considerable interest as to their precise anatomical location, role in neuropeptide homeostasis, significance in peptide trafficking, and potential as a drug delivery system through the blood-brain and/or CSF barriers. In this regard, a peptide transporter was cloned from rat brain and found to be identical to that of rat kidney PEPT2 (Wang et al., 1998). Using in situ hybridization, PEPT2 mRNA was expressed throughout the brain including the epithelial cells of choroid plexus (Berger and Hediger, 1999). Subsequently, the presence of PEPT2, but not PEPT1, protein was confirmed in whole tissue rat choroid plexus by immunoblot analysis (Novotny et al., 2000). The functional evidence for PEPT2-mediated transport of dipeptides was also demonstrated in this tissue (Teuscher et al., 2000). However, whole tissue choroid plexus studies are limited in that they are unable to differentiate membrane sidedness and directionality for peptide transporter activity.
With this in mind, we used rat choroid plexus epithelial cells in primary culture to investigate the peptide-mediated transport mechanisms of a model dipeptide, glycylsarcosine (GlySar), at the blood-CSF interface and the role of PEPT2 in this process.
Experimental Procedures
Materials.
[14C]GlySar (106 mCi/mmol) was purchased from Amersham Biosciences (Piscataway, NJ) and [3H]mannitol (19.9 Ci/mmol) was purchased from PerkinElmer Life Sciences (Boston, MA). Amino acids (glycine, sarcosine, l-histidine), cephalosporins (cefadroxil, cephalexin, cephaloridine, cephalothin), peptides (GlySar, glycylproline, glycylglycylhistidine, carnosine), organic acids (SITS, PAH), and organic bases (TEA, NMN) were purchased from Sigma-Aldrich (St. Louis, MO). Other chemicals were obtained from standard sources and were of the highest quality available.
Primary Culture of Choroidal Epithelial Cells.
Primary cultures of epithelial cells from 1- or 2-day-old rat choroid plexuses were prepared using the method described by Strazielle and Ghersi-Egea (1999). In brief, Sprague-Dawley rat pups (either sex) were killed, the brains were exposed, and choroid plexuses from lateral ventricles were rapidly dissected under a stereomicroscope and kept warm (37°C) in culture medium consisting of Dulbecco's modified Eagle's medium/F-12 (1:1) supplemented with 10% (v/v) fetal bovine serum, 2 mM glutamine, and 50 μg/ml gentamicin (all reagents were obtained from Invitrogen, Carlsbad, CA). Choroidal epithelial cells were isolated and enriched through a series of steps, including tissue digestion, sedimentation and centrifugation, and differential attachment on plastic dishes. Cells were then collected and seeded on laminin-coated Transwell-Clear filter inserts (12-mm diameter, 0.4-μm pore size; Costar Plastics, Cambridge, MA) at a density of 0.65 cm2/plexus for functional activity studies. For immunohistochemical studies, cells were plated at the same density on 10-mm Anocell tissue culture inserts (Whatman Laboratories, Maidstone, UK) containing Anopore membranes (pore size of 0.2 μm) precoated with laminin.
The medium was changed every 48 h and consisted of Dulbecco's modified Eagle's medium/F-12 (1:1) supplemented with 10% (v/v) fetal bovine serum, 2 mM glutamine, 25 μg/ml gentamicin, 5 μg/ml insulin, 5 μg/ml transferrin, 5 ng/ml sodium selenite, 10 ng/ml epidermal growth factor, 2 μg/ml hydrocortisone, and 5 ng/ml basic fibroblast growth factor. Unless otherwise stated, experiments were performed 10 days after seeding. Transepithelial electrical resistance (TEER) measurements were performed immediately prior to experimentation (using a Millicell-ERS; Millipore Corp., Bedford, MA) as a measure of monolayer integrity.
Permeability Studies.
Mannitol permeability was measured in rat choroid plexus cell monolayers grown on 12-mm laminin-coated filters. The uptake buffer contained 10 mM Tris/MES, 147 mM sodium chloride, 2.4 mM KCl, 0.5 mM KH2PO4, 1.1 mM CaCl2, 0.85 mM MgCl2, 0.5 mM Na2SO4, and 5.0 mM glucose (pH 7.4). Culture inserts were rinsed twice with uptake buffer (pH 7.4) on both sides before initiating the study. Then 0.4 ml of uptake buffer (pH 7.4) containing [3H]mannitol (2 μM) was added to the apical side, with 1.2 ml of unlabeled buffer (no mannitol, pH 7.4) added to the opposite side. The incubation proceeded for the indicated period of time at 37°C. Laminin-coated filters without cells were also run in duplicate at the same time. The radioactivity of a 100-μl aliquot from each well was determined by liquid scintillation counting.
GlySar Intracellular Accumulation and Transepithelial Transport.
Cells were incubated with a low-sodium Tris/MES buffer containing 10 mM Tris/MES, 147 mM choline chloride, 2.4 mM KCl, 0.5 mM KH2PO4, 1.1 mM CaCl2, 0.85 mM MgCl2, 0.5 mM Na2SO4, and 5.0 mM glucose (pH 7.4). This buffer has been shown to be advantageous in studying the PEPT2-mediated transport of dipeptides in choroid plexus (Teuscher et al., 2000, 2001).
The transepithelial transport of [14C]GlySar was measured in Transwell chambers, as previously described (Shu et al., 2001). Cell monolayers were preincubated apically and basolaterally with 0.4 and 1.2 ml, respectively, of low-sodium Tris/MES buffer for 10 min at 37°C. The buffer was then removed, and fresh uptake buffer containing [14C]GlySar and [3H]mannitol (2 μM each), with and without 1 mM inhibitors, was added to the apical (0.4 ml) or basolateral (1.2 ml) side; control buffer (no GlySar or mannitol) was added to the opposite side. The incubation proceeded for the indicated period of time at 37°C. To measure transepithelial transport, an aliquot (100 μl) of the buffer was taken from the opposite side, and the radioactivity was counted. To measure intracellular accumulation, the media were aspirated at the end of the incubation period, and the monolayers were rapidly washed four times on both sides with ice-cold uptake buffer. The filters with monolayers were detached from the chambers, and cells were solubilized in 0.5 ml of 0.2 M NaOH and 1% SDS. The radioactivity of the collected buffer and the solubilized cells was determined by liquid scintillation counting. The protein content of the solubilized cell monolayers was determined by the method of Bradford (1976), using the Bio-Rad protein assay kit (Bio-Rad, Hercules, CA) with bovine serum albumin as a standard.
The experimental conditions for GlySar accumulation and transepithelial transport were optimized based on the result of preliminary studies. In particular, a 15-min incubation time was found to be appropriate for estimating the initial rate uptake of GlySar in subsequent kinetic analyses. All data were corrected for the extracellular content and paracellular transport of GlySar, as estimated by mannitol.
GlySar Intracellular Efflux.
For efflux measurements, the cell monolayers were incubated with [14C]GlySar (1 mM) for 60 min at 37°C and washed four times on both sides with ice-cold uptake buffer. The monolayers were then incubated at 37°C with control buffer (no GlySar) in the apical and basolateral chambers. Aliquots (100 μl) of the uptake buffer were taken from both sides at specified times, and the radioactivity was counted. Efflux was expressed as a percentage of the initial GlySar concentration in the cells after loading for 60 min.
Immunoblot Analysis.
Apical membrane vesicles were prepared from choroid plexus cell cultures of neonatal rats and choroid plexus whole tissue (i.e., lateral ventricles) of adult rats, as described previously for rabbit renal brush border membrane vesicles (Akarawut et al., 1998; Lin et al., 1999); this method is similar to that used for apical membrane vesicles of rabbit choroid plexus (Ross and Wright, 1984). Membrane pellets were then solubilized in sample loading buffer (1% SDS, 50 mM Tris-HCl, pH 7.0, 20% glycerol, 5% mercaptoethanol, 0.01 mg/ml bromphenol blue) and heated at 100°C for 3 min. Samples (20–100 μg of protein/lane) were subjected to 7.5% SDS-polyacrylamide gel electrophoresis, and resolved proteins were transferred to nitrocellulose membranes. Antibodies against PEPT1 and PEPT2 were generated previously by immunization of rabbits with keyhole limpet hemocyanin-conjugated synthetic peptides (Shen et al., 1999). After incubation with 6% nonfat dry milk in Tris-buffered saline/Tween 20 (20 mM Tris-HCl, pH 7.5, 150 mM NaCl, 0.1% Tween 20) for 2 h at room temperature, the membranes were incubated with polyclonal antibody (1:1000 dilution in blocking buffer) for 1.5 h at room temperature. The membranes were then washed and incubated with the second antibody (horseradish peroxidase-conjugated goat anti-rabbit IgG, 1:5000; Vector Laboratories, Burlingame, CA), and PEPT1 or PEPT2 protein was detected on X-ray film by an enhanced chemiluminescence system (ECL Plus; Amersham Biosciences).
Confocal Immunofluorescence.
Epithelial cells of rat choroid plexus cultured on Anopore culture inserts were washed in phosphate-buffered saline, then fixed in 2% paraformaldehyde for 30 min at 4°C. Preparations were permeabilized and blocked in solution A (0.2% saponin and 0.1% bovine serum albumin in phosphate-buffered saline) for 30 min, washed in solution A, and then incubated overnight with affinity-purified rat PEPT1 or PEPT2 antisera (1:15 dilution). The secondary antibody, fluorescein isothiocyanate-conjugated goat anti-rabbit IgG (Vector Laboratories) was applied at a 1:100 dilution. The cells were stained with propidium iodide (1 μg/ml) for 1 min and covered by a glass coverslip with ProLong antifade mounting medium (Molecular Probes, Inc., Eugene, OR). Sections were then examined, in planes encompassing the coverslip to filter insert, with an OZ laser scanning confocal fluorescence imaging system coupled to a Nikon microscope (Nikon, Melville, NY) and a computer. The specificity of immunoblot and immunolocalization experiments was assured by preincubation of the antisera with an appropriate immunizing peptide, as reported previously (Shen et al., 1999).
GlySar Stability.
Choroid plexus cells were incubated apically or basolaterally with 1 mM [14C]GlySar for 15 min or 1 h. At the end of incubation, the media from both compartments were aspirated and saved for analysis. The cell monolayers were washed four times with the ice-cold uptake buffer, and 1 ml of ice-cold Milli-Q water (Millipore Corp.) was then added. Cells were scraped off the support and sonicated for 10 min. Cell lysate was treated with acetonitrile, vortexed for 5 s, sonicated for 5 min, and centrifuged for 5 min at 4°C. The supernatant was analyzed by high-performance liquid chromatography, and the concentrations of radiolabeled GlySar and glycine were determined. The stability of GlySar was determined by its recovery and the appearance of glycine following incubation. Results were evaluated from three separate experiments.
Analytical Method.
GlySar and glycine were detected using a high-performance liquid chromatography system consisting of a pump (model 510; Waters, Milford, MA), a reversed-phase column (3-μm, Ultracarbon C-18, 4.6 × 150 mm; Phenomenex, Torrance, CA), and a radiochromatography detector (FLO-ONE 500TR; Packard BioScience, Meriden, CT). The mobile phase was composed of 0.01 M phosphate buffer (pH 2.0) and 0.1% heptafluorobutyric acid, and isocratically pumped at 1 ml/min. Retention times for glycine and GlySar were 2.9 and 5.9 min, respectively, under ambient conditions.
Data Analysis.
The apparent permeability coefficient (Papp) of mannitol was calculated according to the following equation:Papp = Vr · dC/(A · Co · dt), where Vr is the volume of the receiver chamber, A is the surface area of the filter, Co is the initial mannitol concentration in the donor chamber, and dC/dt is the change in mannitol concentration in the receiver chamber per unit of time. A correction was made for filter and aqueous boundary layer effects by determining mannitol transport in the absence of a cell monolayer and applying the equation (Yu and Sinko, 1997): 1/Pmono = 1/Papp − 1/Pcorr, wherePmono is the permeability of the cell monolayer and Pcorr is the measured permeability using blank filters (i.e., 1/Pcorr = 1/Paqueous + 1/Pfilter).
For kinetics studies, the concentration-dependent uptake of GlySar (V) was best fit to the following relationship: V =Vmax · C/(Km + C) +Kd · C, whereVmax is the maximal rate of uptake,Km is the Michaelis constant,Kd is the rate constant for nonsaturable transport, and C is the substrate (GlySar) concentration.
Data are reported as mean ± S.E. of three or four independent experiments, with data from each experiment being determined in duplicate or triplicate. Statistical differences among treatment groups were evaluated by analysis of variance, and pairwise comparisons were made using Tukey's test (α = 0.05). All statistical computations were performed using SYSTAT (version 8.0; Systat, Inc., Evanston, IL). Nonlinear as well as linear regression analyses were conducted using Scientist (version 2.01; MicroMath Scientific Software, Salt Lake City, UT) and a weighting factor of unity. The quality of the fit was determined by evaluating the coefficient of determination (r2) and the standard error of parameter estimates, and by visual inspection of the residuals.
Results
Formation of Polarized Monolayers with Functional Barrier Properties.
Cell attachment on laminin-coated filters was complete by 48 h, at which time the media were changed, and optical confluence was achieved 2 to 3 days thereafter. As observed in Fig.1, the cells grew as small, densely packed, polygonal shapes and produced a monolayer displaying a typical cobblestone appearance. Transmission electron microscopy was also performed on cultured choroid plexus cells to confirm the presence of a differentiated monolayer of epithelial cells (data not shown), as reported previously (Strazielle and Ghersi-Egea, 1999).
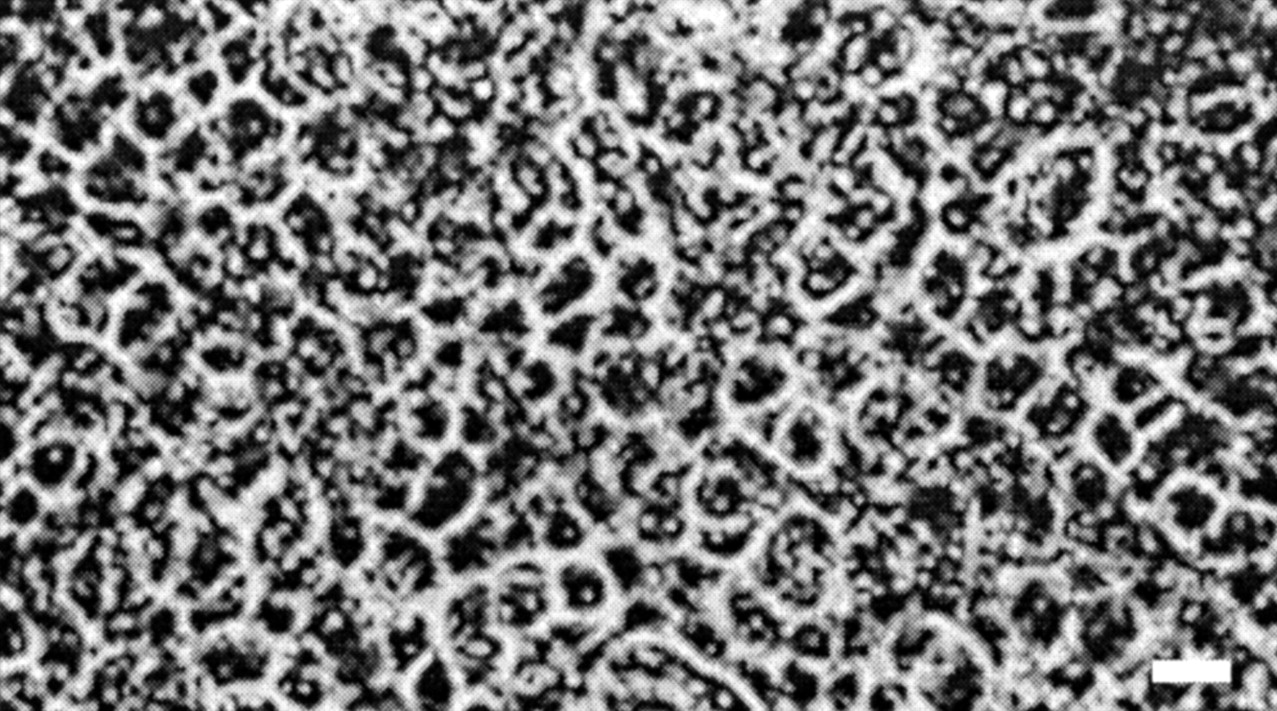
Phase-contrast micrograph of a confluent monolayer of rat choroid plexus epithelial cells on laminin-coated Transwell filters (culture time 10 days). Scale bar, 10 μm.
The in vitro establishment of barrier properties was demonstrated by monitoring the TEER of cells cultured on semipermeable supports along with the paracellular transport of a hydrophilic marker, mannitol. As shown in Fig. 2A, TEER values increased rapidly after day 2 and reached a plateau around day 10. The TEER value of 155 Ω · cm2 at day 10 is similar to the TEER measured in isolated bullfrog choroid plexus mounted in an Ussing chamber (Saito and Wright, 1983). The maximal TEER was maintained until day 16, at which time these values steadily fell. Permeability to mannitol was inversely related to the electrical resistance, and a Pmono value of 2.1 × 10−4 cm/min was observed at day 10 (Fig. 2B); this value is similar to that reported by Strazielle and Ghersi-Egea (1999). Moreover, the transfer of mannitol was several times lower across inserts with cells (14-fold at 15 min) compared with control inserts (i.e., filters without cells), indicating that the monolayer acts as a significant barrier to paracellular transport (Fig. 2C).

Development of a permeability barrier in rat choroid plexus monolayers, as demonstrated by TEER versus culture time (panel A), mannitol permeability (2 μM) of the cell monolayer versus culture time (panel B), and mannitol transport (2 μM) versus time (panel C). In panel A, data were corrected for the mean electrical resistance of laminin-coated filters (without cells). In panel B, data were corrected for filter and aqueous boundary layer effects. Studies were performed in Tris/MES buffer (pH 7.4/7.4 for donor/receiver compartments), and data were expressed as mean ± S.E. from four separate experiments. Error bars are smaller than the symbols when not evident.
pH Dependence of GlySar Accumulation.
Since peptide-mediated transport is proton-coupled, the effect of pH on GlySar accumulation was examined at both the apical and basolateral sides of choroid plexus cells. This was achieved by varying the buffer pH in the donor chamber from 5.5 to 8.0 while maintaining the pH of the receiver chamber at 7.4. As shown in Fig. 3, the GlySar uptake from the apical side was markedly influenced by the pH of the apical medium, with a maximal uptake at pH 6.0. In contrast, GlySar uptake from the basolateral side appeared to be pH-insensitive. These findings suggest that a peptide transporter (or transporters) may be located at the apical membrane of choroidal epithelium. Nevertheless, for all subsequent experiments, neutral uptake buffer (pH 7.4) was used in both the apical and basal chambers to provide a more physiological situation.
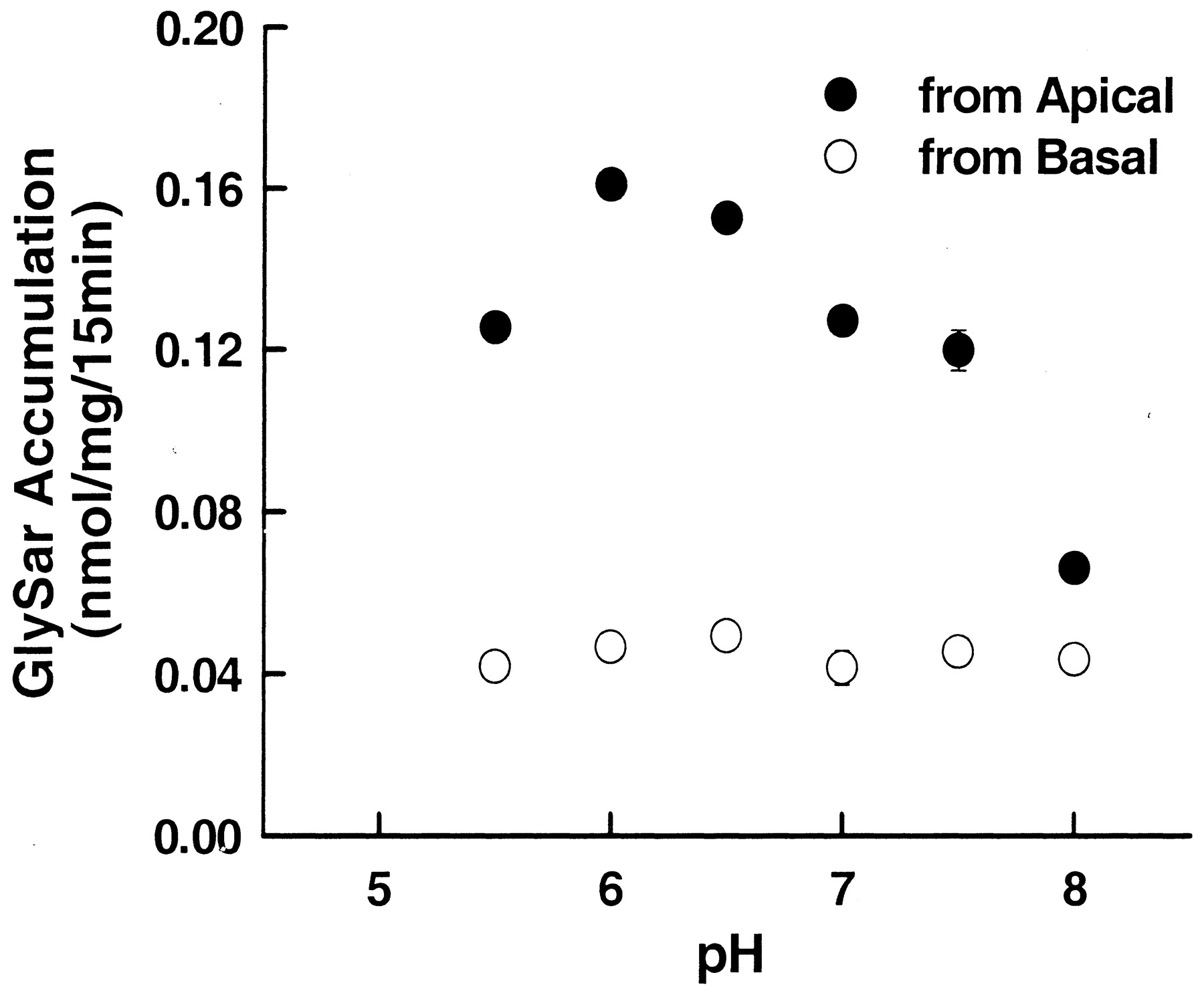
pH-dependent accumulation of GlySar in choroid plexus epithelial cells. The 15-min uptake of [14C]GlySar (2 μM) was measured in monolayers grown in Transwell chambers containing low-sodium Tris/MES buffer (pH 5.5–8.0 versus pH 7.4 for donor versus receiver compartments, respectively) at 37°C. All data were mannitol-corrected and were expressed as mean ± S.E. from four separate experiments.
Intracellular Accumulation and Transepithelial Transport of GlySar.
The intracellular accumulation and transepithelial transport of radiolabeled GlySar were evaluated in choroid plexus epithelial cells using pH 7.4/7.4 for donor/receiver chambers. As observed in Fig. 4A, GlySar accumulation was far more rapid when introduced from the apical as opposed to the basolateral surface of the membrane (i.e., 3-fold difference). Similarly, the apical-to-basolateral transport of GlySar was substantially faster than that of GlySar in the reverse direction (Fig.4B). These findings suggest a unidirectional transcellular transport that corresponds to peptide efflux from the CSF to blood.

Intracellular accumulation (panel A) and transepithelial transport (panel B) of GlySar in choroid plexus epithelial cells. Monolayers grown in Transwell chambers were preincubated apically (0.4 ml) and basolaterally (1.2 ml) with low-sodium Tris/MES buffer (pH 7.4) for 10 min at 37°C. Buffer was removed and appropriate volumes of fresh uptake buffer, containing [14C]GlySar and [3H]mannitol (2 μM of each), were added to the apical or the basolateral side (pH 7.4/7.4 for donor/receiver compartments); control buffer (i.e., no GlySar or mannitol) was added to the opposite side. Incubation proceeded for the specified period of time at 37°C. All data were mannitol-corrected and were expressed as mean ± S.E. from four separate experiments.
GlySar Efflux from Choroid Plexus Cell Monolayers.
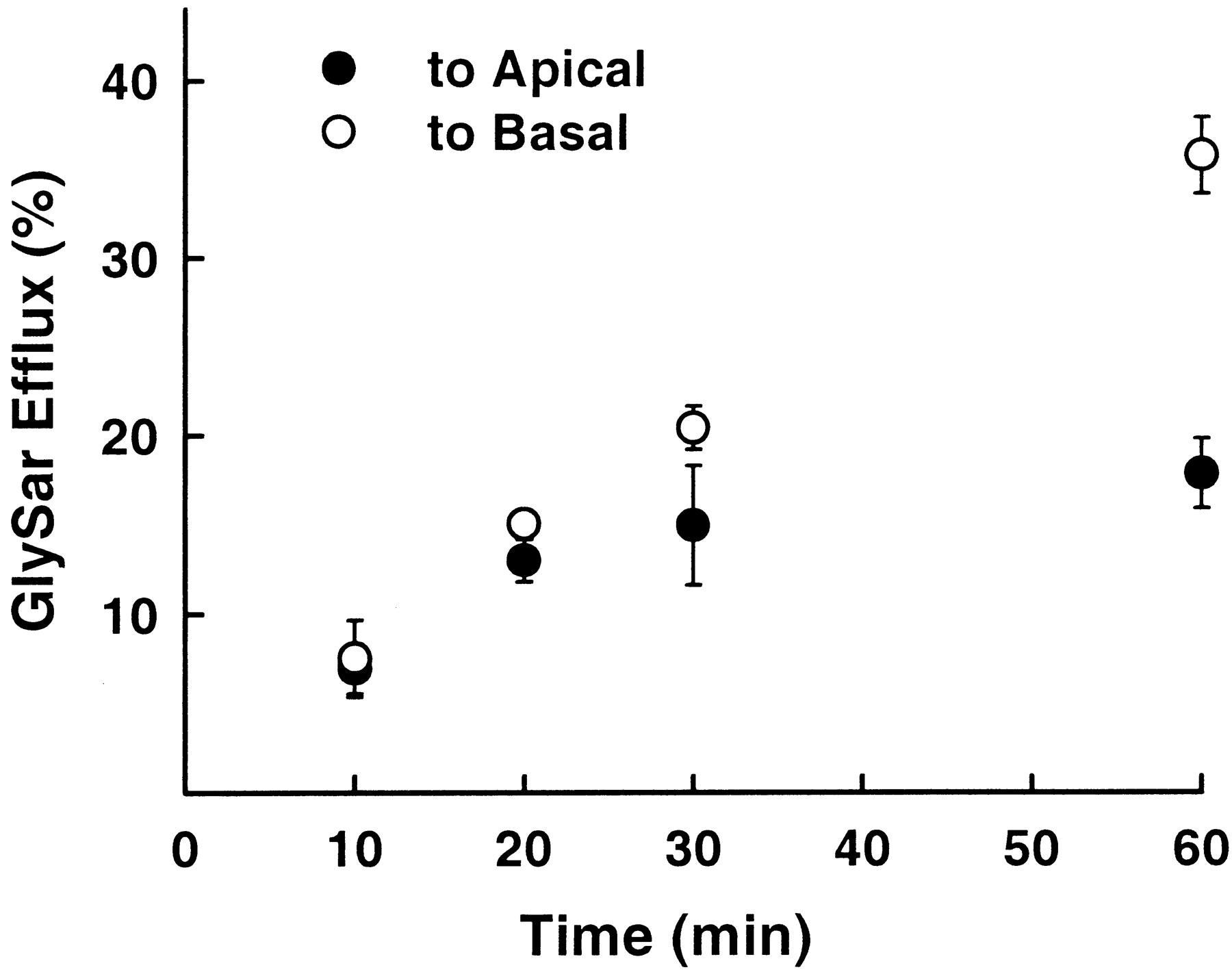
To probe the fate of small peptides or mimetics once in the cell, GlySar efflux was examined after preloading. As shown in Fig.5, little difference was noted in the time course of GlySar appearance in the apical or basal chambers over the first 20 min. However, at the 30- and 60-min time points, GlySar was accumulated to a greater extent on the basal side compared with the apical side and reached an apparent plateau, suggesting equilibration between the apical and cell compartments. This finding is consistent with a peptide transporter being present on the apical surface, with GlySar being directed back into the cell. It also demonstrates that GlySar is preferentially transported across the basolateral membrane (equivalent to the blood side) once in the cell.

Efflux of GlySar from choroid plexus epithelial cells. Monolayers grown in Transwell chambers were preincubated apically (0.4 ml) and basolaterally (1.2 ml) with low-sodium Tris/MES buffer (pH 7.4) for 10 min at 37°C. Buffer was removed and an appropriate volume of fresh uptake buffer, containing [14C]GlySar (1 mM), was added to the apical side (pH 7.4) for 60 min; control buffer (i.e., no GlySar) was added to the basolateral side (pH 7.4). After several washes, monolayers were incubated at 37°C with control buffer (no GlySar) in both the apical and basolateral compartments. Efflux was expressed as a percentage of the initial GlySar concentration in the cells after loading for 60 min and represents the mean ± S.E. from four separate experiments.
Concentration-Dependent Uptake of GlySar.
To compare the affinity of GlySar for peptide transport systems in polarized membranes, the concentration dependence of GlySar uptake was evaluated from either the apical or the basolateral side of choroid plexus cells. The results are displayed in Fig. 6 (A and B) in which Km values were 59.6 ± 0.9 μM and 1.4 ± 0.2 mM when GlySar uptake was measured from the apical and basal membranes, respectively.Vmax values were 0.46 ± 0.01 and 1.77 ± 0.13 nmol/mg/15 min, respectively, for apical and basal membranes. Moreover, Kd values were small (0.00042 ml/mg/15 min, apical; 0.00012 μl/mg/15 min, basal) and, as a result, under linear conditions, carrier-mediated transport accounted for ≥95% of the total transport apically or basally. GlySar interacted with a single specific transporter at each membrane surface under physiologic pH conditions, as demonstrated by the linear relationship of the transformed data (insets for Fig. 6). These findings suggest that distinct transport systems may exist at the apical and basolateral membranes and that the kinetic parameters of the apical membrane are consistent with the high-affinity properties of PEPT2.

Concentration-dependent uptake of GlySar in choroid plexus epithelial cells. The 15-min uptake of [14C]GlySar (2–5000 μM) was measured in monolayers grown in Transwell chambers incubated apically (panel A) or basolaterally (panel B) with low-sodium Tris/MES buffer (pH 7.4/7.4 for donor/receiver compartments) at 37°C. All data were corrected for nonsaturable uptake and expressed as mean ± S.E. from four separate experiments. Lines were generated using fitted mean parameters (see text), as determined by nonlinear regression analysis. Insets are Woolf-Augustinsson-Hofstee plots of the transformed data [uptake (v), nmol/mg/15 min versus uptake per concentration (v/S), μl/mg/15 min].
Inhibitor Analysis.
Specificity of peptide-mediated transport was examined using a wide range of potential inhibitors of [14C]GlySar uptake from choroid plexus epithelial cells (pH 7.4/7.4 for donor/receiver chambers). When probed from the apical membrane (Fig. 7A), GlySar uptake was reduced about 80 to 90% by di- and tripeptides (i.e., glycylproline, GlySar, glycylglycylhistidine, and carnosine) but was unaffected by amino acids (i.e., glycine, sarcosine, andl-histidine). Although the aminocephalosporins cefadroxil and cephalexin inhibited about 85% of GlySar uptake, cephalosporins without an α-amino group (i.e., cephaloridine, cephalothin) had no effect. These results are consistent with the fact that aminocephalosporins and dipeptides share a common transport system. In contrast, prototypical organic anions (i.e., SITS, PAH) and organic cations (i.e., TEA, NMN) showed no effect on GlySar uptake. Interestingly, a similar pattern of inhibition was observed when GlySar uptake was probed from the basal membrane (Fig. 7B). However, the inhibitory effects were markedly reduced, being on the order of about 35 to 75%.

Effect of potential inhibitors on the uptake rate of GlySar from apical (panel A) and basolateral (panel B) membranes. Monolayers grown in Transwell chambers were preincubated apically (0.4 ml) and basolaterally (1.2 ml) with low-sodium Tris/MES buffer (pH 7.4) for 10 min at 37°C. Buffer was removed and appropriate volumes of fresh uptake buffer, containing [14C]GlySar and [3H]mannitol (2 μM each) with and without 1 mM inhibitors, were added to the apical or the basolateral side (pH 7.4/7.4 for donor/receiver compartments); control buffer (i.e., no GlySar or mannitol) was added to the opposite side. Incubation proceeded for the specified period of time at 37°C. All data were mannitol-corrected and expressed as mean ± S.E. from four separate experiments. ★★, P < 0.01, ★★★,P < 0.001 compared with control values.
Stability Study.
The stability of GlySar was determined in the donor, receiver, and intracellular compartments of choroid plexus epithelial cells. Regardless of whether the compound was introduced at the apical or basolateral membrane surface, significant hydrolysis of GlySar was not evident in any of the samples being tested. Following incubation for up to 1 h, more than 99% of GlySar was recovered intact (i.e., <1% hydrolysis to glycine). This finding clearly demonstrates that GlySar was intact during the accumulation, transport, and efflux studies in choroidal cell cultures.
Evidence for PEPT2 Protein.
Since the present study suggested an apical location for the peptide transporter, immunoblot analyses were performed using apical membrane vesicles prepared from choroid plexus cell cultures. Protein was also extracted from renal and intestinal brush border membrane vesicles for use as positive controls for PEPT2 and PEPT1, respectively. As shown in Fig.8A, a primary hybridization band of about 85 kDa was detectable in neonatal choroid plexus cells using PEPT2 antisera, as was a broad band of similar mass for rat kidney. In contrast, PEPT1 antisera failed to detect a brush border antigen in neonatal choroid plexus epithelial cells; however, a strong signal was observed at about 90 kDa for rat intestine. In a similar manner, PEPT2 but not PEPT1 protein was observed in apical membrane vesicles prepared from adult choroid plexus whole tissue (Fig. 8B).

Immunoblot analyses of PEPT2 and PEPT1 in apical membrane vesicles prepared from choroid plexus cell cultures of neonatal rats (panel A) and choroid plexus whole tissue of adult rats (panel B). As positive controls, apical membrane vesicles were prepared from the kidney and small intestine of adult rats (panels A and B). PEPT2 is observed as a band at 85 kDa and PEPT1 as a band at 90 kDa. BBMV, brush border membrane vesicles.
Confocal Microscopy of Choroid Plexus Epithelial Cells.
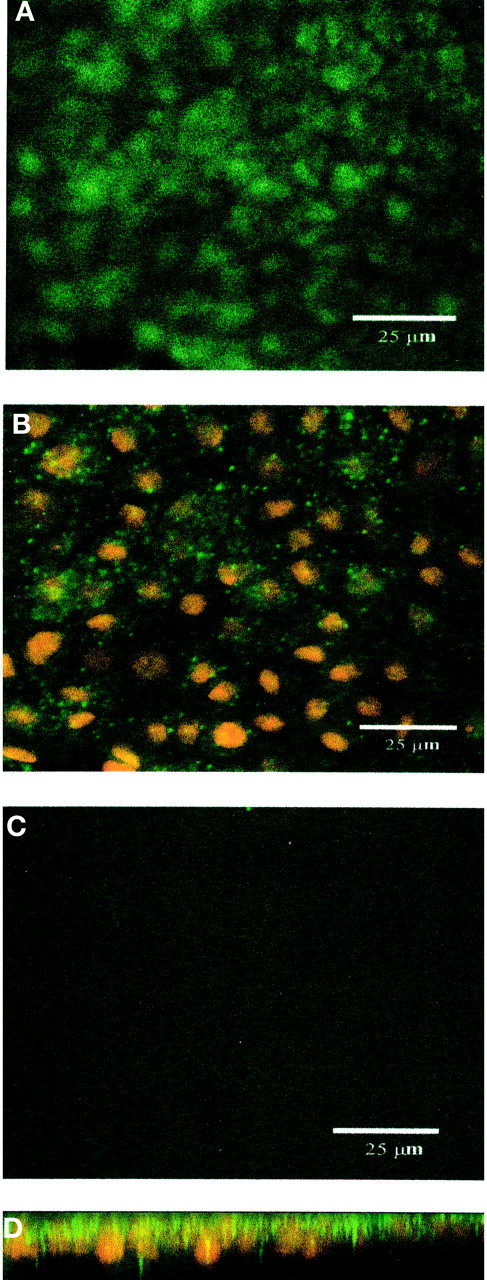
To further characterize the cellular expression of PEPT2, primary cultures of choroid plexus epithelial cells obtained from neonatal rat were established and grown on Anopore membrane filters. Formation of confluent cell monolayers with tight junctions and apical microvilli, features characteristic of native choroid plexus epithelia in vivo, was confirmed by electron microscopy. Cells were grown under conditions identical to those used for functional studies, and monolayer integrity was maintained, as demonstrated by high TEER values. In 10-day-old cultures, indirect immunofluorescence with confocal microscopy detected PEPT2 with a granular staining pattern at the apical surface of the cell (Fig. 9A). PEPT2 was also observed subapically, although the staining was less dense (Fig. 9B). Optical sectioning of the z-series perpendicular to the plane of the cell monolayer confirmed a predominantly apical (and subapical) distribution for PEPT2 (Fig. 9D). In contrast, no specific fluorescence was observed for PEPT2 at the basolateral membrane (Fig. 9C). Consistent with our previous data from Western blot analysis (Novotny et al., 2000), PEPT1 staining was not evident in choroid plexus epithelial cells (Fig.10).

Immunolocalization of PEPT2 expression in rat choroid plexus epithelial cells in primary culture by confocal microscopy (green signal). Nuclei were detected with propidium iodide (red signal). Top-down views of apical cell layer (panel A), cell layer across the nuclear plane (panel B), and basal cell layer (panel C); optical sectioning of the z-series perpendicular to the plane of the cell layer (panel D). PEPT2 localizes predominantly toward apical surfaces. Scale bar, 25 μm.
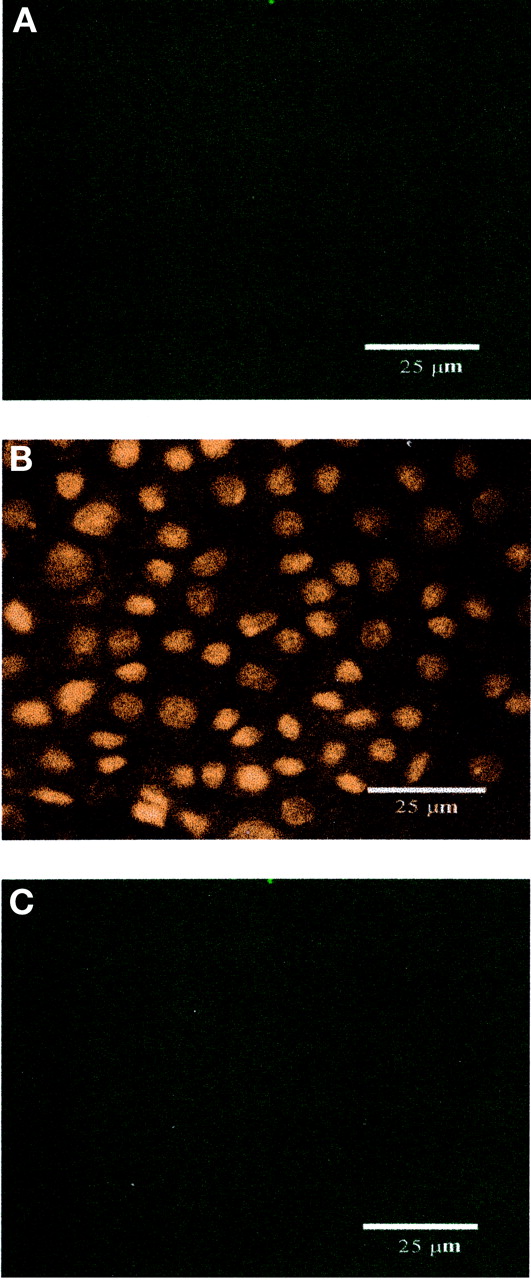
Immunolocalization of PEPT1 expression in rat choroid plexus epithelial cells in primary culture by confocal microscopy (green signal). Nuclei were detected with propidium iodide (red signal). Top-down views of apical cell layer (panel A), cell layer across the nuclear plane (panel B), and basal cell layer (panel C). No PEPT1 expression was observed in rat choroid plexus epithelium. Scale bar, 25 μm.
Discussion
PEPT2 has been cloned from rat brain (Wang et al., 1998), along with PHT1 (Yamashita et al., 1997) and PHT2 (Sakata et al., 2001), and PEPT2 mRNA transcripts have been found in several regions of the central nervous system, including the choroid plexus (Berger and Hediger, 1999). More recently, the functional activity of GlySar was demonstrated in whole tissue rat choroid plexus (Teuscher et al., 2000), as was PEPT2 protein (Novotny et al., 2000). The presence of peptide transporters in the brain and, in particular, choroid plexus has generated substantial interest as to their physiologic role and pharmacologic implications. Along these lines, PEPT2 mediated the uptake of a diverse group of neuropeptides in isolated rat choroid plexus (Teuscher et al., 2001), suggesting a role for PEPT2 in the regulation of neuropeptides, peptide fragments, and peptidomimetics in cerebrospinal fluid. Although these studies have indicated a presence for the high-affinity peptide transporter PEPT2 in choroid plexus whole tissue, they were not able to delineate the precise membrane location, subcellular distribution, or directionality of PEPT2.
In the present study, several pieces of evidence point to the apical expression of PEPT2 in choroid plexus epithelial cells. First, GlySar uptake by the apical peptide transporter was influenced by medium pH, whereas pH had little effect on the basolateral transporter. Second, transporter affinity for GlySar from apical membranes was consistent with the high-affinity properties of PEPT2 (i.e.,Km value of 60 μM). Third, immunoblots established that PEPT2 was present on the apical surface. Finally, immunofluorescent confocal microscopy demonstrated that PEPT2 was localized to apical (and subapical) regions but was not present in basolateral membranes. This study also points to the PEPT2-mediated translocation of dipeptide from CSF to cell. Hence, apical uptake and apical-to-basal transepithelial transport of GlySar were much greater in choroid plexus cell monolayers than the reverse processes. Once inside the cell, GlySar was preferentially effluxed to the basal compartment. These results are consistent with the study by Huang (1982) in which Tyr-d-Ala-Gly was preferentially cleared out of the CSF from the apical CSF-facing membrane of rat choroid plexus. It should also be appreciated that PEPT2 expression was maintained in apical membranes of rat choroid plexus from neonatal to adult ages. Apical PEPT2 expression in adult rat choroid plexus is further supported by the similarity in GlySar uptake in whole tissue (i.e., Km value of 129 μM; Teuscher et al., 2000).
These findings do not rule out the possibility of other membrane proteins participating in the apical or basolateral transport of GlySar in choroid plexus. However, it is unlikely that PEPT1 is involved since previous immunoblots failed to detect this protein in whole tissue (Novotny et al., 2000) and, in this study, PEPT1 staining was not visible anywhere in the choroid plexus cells during immunolocalization experiments. PHT1 and PHT2 can also be ruled out as candidate proteins since GlySar uptake was not inhibited by saturating concentrations ofl-histidine when present at either the apical or the basal surface of the cell. At this point, the role of peptide/histidine transporters in choroid plexus is uncertain. However, they may have a role in the intracellular trafficking of small peptides, as suggested by the lysosomal expression of rat PHT2 in transfected baby hamster kidney and human embryonic kidney-293T cells (Sakata et al., 2001).
The coordinated expression of PEPT2 in choroid plexus apical and subapical compartments serves to underscore the complexity of neuropeptide homeostasis in the CNS. It has been proposed that termination of neuropeptide activity is not the result of reuptake systems alone but may also be affected by diffusive and enzymatic processes (Konkoy and Davis, 1996). Neuropeptides are broken down to their constitutive amino acids by a variety of peptidases, being both membrane-bound and cytosolic. For example, the tripeptidyl-peptidases (TPP) I and II have been detected in choroid plexus epithelium (Facchinetti et al., 1998; Kida et al., 2001). TPP II has a neutral pH optimum and is located in the cytoplasm, whereas TPP I has an acidic pH optimum, suggesting a lysosomal localization (Page et al., 1993; Vines and Warburton, 1998; Ezaki et al., 1999). The lysosomal peptidases cleave neuropeptides into tripeptides that can be further degraded into dipeptides and amino acids, and then exported to the cytoplasm. It is reasonable to suggest that peptide fragments could be removed from lysosomes by specific peptide transporters. Although the responsible protein has not been identified, the subapical expression of PEPT2 in choroid plexus epithelial cells, along with its proton-coupled symport, make it a candidate transporter for peptide efflux from the lysosomes. However, further studies will be needed to confirm this hypothesis.
The preference of GlySar for apical-to-basal transport and cellular efflux to the basal compartment suggests that small peptides are translocated from the CSF to blood, rather than the reverse. Having distinct peptide transporters at each membrane surface of the choroid plexus may aid in this process. In the present investigation, a high-affinity protein, PEPT2, was found at the apical membrane. Although a low-affinity uptake (i.e., millimolar value forKm) was observed for GlySar at the basolateral membrane, the molecular properties of this protein have not been delineated. This finding agrees with our general lack of knowledge concerning the cellular efflux mechanism of peptides/mimetics across the basolateral membrane of other epithelia (e.g., intestine and kidney). Notwithstanding this uncertainty, it has been shown in this study and others (Thwaites et al., 1993a,b; Terada et al., 1999, 2000;Shu et al., 2001) that dipeptides, tripeptides, and peptidomimetics interact with basolateral transport systems that are distinct from PEPT1 and PEPT2. Ultimately, cloning should help to elucidate the precise nature of the basolateral peptide transporter in choroid plexus, as well as in other tissues.
The PEPT2-mediated uptake of peptides is energized by the electrochemical gradient of protons across the intestinal or renal brush border membrane in the lumen-to-cytoplasm direction (Nussberger and Hediger, 1995). This gradient is even more dramatic when comparing intracellular pH with the unstirred water layer lining the surface of the apical membrane (Lucas, 1983). To our knowledge, there is no information available on the magnitude of proton gradients across the plasma membrane of any of the cell types in the nervous system (Wang et al., 1998). For this reason and because of physiological considerations, choroid plexus epithelial cells in primary culture were studied at pH 7.4 in both the donor and receiver chambers (except for pH-dependent studies). Although we did not measure the pH of choroid plexus epithelial cells, it is known that the normal intracellular pH of choroid plexus is about 7.0 (Johanson, 1978) and that it is regulated less effectively in vitro than in vivo. Considering the physiological bulk fluid of CSF (pH of 7.3), it is unlikely that an inwardly directed pH gradient exists between the CSF and choroidal cells. However, it is possible that an acid microclimate (an area of low pH adjacent to the apical membrane) may exist at the choroidal apical membrane, such as that observed in the intestine and kidney. Alternatively, in the absence of a pH microclimate, GlySar accumulation may be driven by the electrochemical gradient of membrane potential that exists in choroid plexus epithelium (Saito and Wright, 1983). Thus, PEPT2-mediated uptake may be energized by the pH and/or membrane potential differences that develop at the microclimate of apical villi (Boyd and Ward, 1982; Ganapathy and Leibach, 1983; Thwaites et al., 1993c).
In conclusion, this study is unique in demonstrating the preferential uptake and transepithelial transport of a model dipeptide (GlySar) from the apical surface of choroid plexus epithelial cells. Although immunoblot and confocal immunofluorescence studies placed PEPT2 at the apical (and subapical) regions of the choroidal cell, immunolocalization studies could not detect PEPT1 or functional studies, PHT1 and PHT2. Taken as a whole, these findings suggest that PEPT2 may have a role as an efflux pump for the effective removal of peptides from CSF into the blood. Furthermore, the pharmacokinetics of peptidomimetic drugs in the CNS (e.g., aminocephalosporins and penicillins) may be impacted by PEPT2 activity at the blood-CSF barrier. Thus, maximizing the CNS penetration of therapeutic peptides and/or mimetics may require the concurrent blockade of PEPT2.
Acknowledgments
We thank Thomas Komorowski of the Michigan Diabetes Research and Training Center Morphology and Image Analysis Core for help with the confocal immunofluorescence experiments.
Footnotes
-
This work was supported in part by Grants R01 GM035498 (to D.E.S.), R01 NS034709 and P01 HL018575 (to R.F.K.), P60 DK020572 (Michigan Diabetes Research and Training Center Core) from the National Institutes of Health, and by the Vahlteich Research Award from the University of Michigan College of Pharmacy. N.S.T. was supported by an American Foundation for Pharmaceutical Education Fellowship, a Rackham Predoctoral Fellowship, and the Pharmacological Sciences Training Program of the National Institutes of Health (Grant T32 GM007767).
- Abbreviations:
- CSF
- cerebrospinal fluid
- GlySar
- glycylsarcosine
- TEER
- transepithelial electrical resistance
- MES
- 2-(N-morpholino)ethanesulfonic acid
- HPLC
- high-performance liquid chromatography
- CNS
- central nervous system
- TPP
- tripeptidyl-peptidases
- SITS
- 4-acetamido-4′-isothiocyanatostilbene-2,2′-disulfonic acid
- PAH
- p-aminohippuric acid
- NMN
- N1-methylnicotinamide
- Received December 6, 2001.
- Accepted February 20, 2002.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}