


Abstract
Therapeutic molecules conjugated with antibodies to the platelet-endothelial cell adhesion molecule-1 (PECAM-1) accumulate in the pulmonary endothelium after i.v. injection in mice. In this study, we characterized PECAM-directed targeting to the lung and heart after local versus systemic intravascular administration in a large animal model, pigs. Radiolabel tracing showed that 1 h post-i.v. injection, 35% of anti-PECAM versus 2.5% of control IgG had accumulated in the lungs. Infusion of anti-PECAM via a catheter placed in the right pulmonary artery (RPA) resulted in a 3-fold elevation of the uptake in the right lower lobe and 2-fold reduction of uptake in the left lobes in the lung. Cardiac uptake of anti-PECAM was negligible after i.v. and RPA infusion. In contrast, delivery with a catheter placed in the right coronary artery (RCA) resulted in a 4-fold elevation of cardiac uptake of anti-PECAM, but not IgG, compared with i.v. injection. To estimate the targeting of an active reporter enzyme, streptavidin-conjugated β-galactosidase (β-Gal) was coupled to anti-PECAM or IgG (anti-PECAM/β-Gal and IgG/β-Gal) and injected into the RCA. β-Gal activity was markedly elevated in the heart and lungs (5- and 25-fold increased, respectively) after injection of anti-PECAM/β-Gal, but not IgG/β-Gal. Image analysis confirmed endothelial targeting of anti-PECAM/β-Gal in the heart and lung. In summary, anti-PECAM antibody conjugates deliver agents to the pulmonary endothelium regardless of injection route, whereas local arterial infusion permits targeting to the cardiac vasculature. This paradigm may be useful for drug targeting to endothelium in lungs, heart, and possibly other organs.
The endothelium represents a large, accessible, and important target for the treatment of a spectrum of cardiovascular and pulmonary diseases. However, most drugs and drug/gene vehicles (e.g., liposomes) have no significant specific affinity to the endothelium and undergo fast elimination by the bloodstream, even after local infusion via vascular catheters. Therefore, effective delivery requires administration of high doses and prolonged contact with endothelium (Schulick et al., 1995; Rekhter et al., 1998). Accordingly, specific devices, which allow transient cessation of blood flow in the site of catheter placement, have been designed to attain more effective prolonged interaction of the infused material with vascular targets (Varenne et al., 1998). However, this may lead to tissue ischemia and vascular injury (Channon et al., 1998; Wright et al., 1998). Therefore, the lack of effective, rapid, and safe targeting of therapeutic molecules to endothelium represents an important biomedical problem.
Vascular immunotargeting, a strategy based on conjugation of a diagnostic or therapeutic molecule with antibodies directed against specific endothelial surface determinants, may help to solve the problem (Muzykantov, 1998). A promising target determinant is the CD31 antigen (platelet-endothelial cell adhesion molecule-1, PECAM), which is abundantly, stably, and ubiquitously expressed on the surface of endothelial cells (Newman, 1997). Active reporter enzymes and genetic materials coupled to PECAM antibodies (anti-PECAM conjugates) bind selectively to endothelial cells in culture and in the pulmonary vessels in mice after intravenous administration (Muzykantov et al., 1999; Christofidou et al., 2000; Li et al., 2000; Scherpereel et al., 2001; Wiewrodt et al., 2002).
The lungs receive the first pass of the entire cardiac output of venous blood and have a high capillary density. In fact, roughly 30% of the total endothelium in the body belongs to the pulmonary vasculature (Davis and Hagen, 1993). Thus, even antibodies that do not discriminate between pulmonary and systemic endothelium (e.g., anti-PECAM) accumulate in the lungs after i.v. injection (Muzykantov et al., 1999;Danilov et al., 2001).
However, because PECAM-1 is highly expressed on the surface of endothelial cells in other vascular beds, we hypothesized that the potential utility of anti-PECAM targeting is not limited to the pulmonary vasculature and that local vascular administration of anti-PECAM conjugates may facilitate uptake in endothelial cells in other organs. To test this hypothesis and to further extend the anti-PECAM immunotargeting paradigm, we studied the targeting of anti-PECAM conjugates injected in the vasculature regionally versus systemically. To achieve this goal with conventional catheterization techniques, we chose to study a large animal model, the pig. The results demonstrate, for the first time, that local intravascular delivery of anti-PECAM conjugates markedly enhances the uptake in the cardiac vasculature, thus permitting organ-selective delivery of an active reporter molecule to endothelial cells in the heart.
Materials and Methods
Antibodies and Enzymes.
mAb 62 is a monoclonal mouse IgG2 antibody directed against human platelet-endothelial adhesion molecule-1 with cross-reactivity with PECAM isoforms of other species, as previously described (Muzykantov et al., 1999; Nakada et al., 2000). A murine monoclonal antibody mAb T412 directed against an irrelevant human antigen that showed no reactivity with pig tissues and cells (provided by Centocor, Malvern, PA) was used as an isotype-matched control mouse IgG2. β-Galactosidase covalently coupled to streptavidin (SA-β-Gal, with a specific β-Gal activity 400 units/mg) was purchased from Sigma Chemical (St. Louis, MO).
Immunohistochemical Detection of PECAM in Pig Tissues.
Immunohistochemical analysis was done on 6-μm-thick frozen sections from optimal cutting temperature compound (VWR Scientific Products, Irving, TX)-embedded tissues. Tissue sections were incubated with anti-PECAM mAb 62 or with isotype-matched, control mouse IgG (20 μg/ml), and processed further for immunostaining by using the Vectastain kit (Vector Laboratories, Burlingame, CA) according to the manufacturer's protocol. Briefly, antibody-coated sections were incubated with a biotinylated, affinity-purified anti-mouse IgG secondary antibody, washed, and incubated further with a conjugate of avidin/biotinylated horseradish peroxidase (Vectastain ABC Elite Reagent). The tissue sections were then incubated with the peroxidase substrate solution and the chromogenic reaction product (red/brown) evaluated using an Olympus II photomicroscope.
Biotinylation and Radiolabeling of Proteins.
Proteins were biotinylated with 6-biotinylaminohexanoic acidN-hydroxysuccinimide ester (Pierce Chemical, Rockford, IL) and designated as b-IgG or b-anti-PECAM. Streptavidin and control murine IgG were labeled with 125I or131I (PerkinElmer Life Sciences, Boston, MA) by using Iodogen (Pierce Chemical) according to the standard manufacturer's instructions. The radiolabeled125I-SA was coupled with b-mAb 62 at equimolar ratio, as described previously, to form an anti-PECAM/125I-SA conjugate (Muzykantov et al., 1999). The conjugate and control 131I-IgG are designated in the following text as125I-anti-PECAM and131I-IgG.
Conjugation of Proteins.
SA-β-Gal was coupled to biotinylated anti-PECAM or IgG as described previously (Scherpereel et al., 2001). Before conjugation with biotinylated proteins, SA-β-Gal was dissolved at 0.5 mg/ml in endotoxin-free PBS, pH 7.4. Either b-anti-PECAM or b-IgG was added dropwise to SA-β-Gal in a vortex mixer at room temperature. The size of the resulting anti-PECAM/SA-β-Gal and IgG/SA-β-Gal conjugates was determined using a Dynamic light scattering (Brookhaven Instruments, Brookhaven, NY), with a 90o angle, as described (Scherpereel et al., 2001). The size of the conjugates depends on the molar ratio of the b-IgG or b-anti-PECAM and SA-β-Gal or streptavidin. For example, the molar ratio SA-β-Gal to b-anti-PECAM or b-IgG providing nonaggregated, stable conjugates was established to be from 1.0 to 1.5. Dynamic light scattering analysis revealed that the conjugates formed at these conditions have diameters of 100 to 200 nm, whereas the size of the individual components was approximately 15 and 30 nm for immunoglobulins and SA-β-Gal, respectively. Thus, streptavidin cross-linking of biotinylated immunoglobulins forms high-molecular-weight aggregates. The resulting conjugates are designated hereafter in the text as anti-PECAM/β-Gal and IgG/β-Gal. At optimal molar ratios, both SA-β-Gal and streptavidin form conjugates of similar size with radiolabeled and nonlabeled b-anti-PECAM or b-IgG. The size of the conjugates is stable for several weeks. A full discussion of the importance of anti-PECAM/streptavidin conjugate size is included in our recent article (Wiewrodt et al., 2002).
Animal Study.
All animal procedures were approved by the Institutional Animal Care and Use Committee of the Children's Hospital of Philadelphia. Twenty-five pigs aged 7 to 25 days and weighing 3.1 ± 0.2 kg were used for the study. After intramuscular administration of ketamine (20 mg/kg) and acepromazine (1 mg/kg), intravenous access was obtained via femoral vein. All animals received 100 units/kg heparin. Femoral arterial and venous catheters were placed percutaneously in all animals. In pigs designated for right coronary artery catheterization, a 4 French sheath (Cordis, Inc., Miami, FL) was placed into right carotid artery via cut down to facilitate coronary catheterization. Fluoroscopic guidance was used to selectively catheterize the right pulmonary artery (n = 5) or right coronary artery (n = 13). Contrast injections were performed to document catheter position in these animals.
In Vivo Administration of 125I-Anti-PECAM and131I-IgG.
Ten micrograms of125I-anti-PECAM mixed with same amount of131I-IgG was infused as a mixture in 0.5 ml of saline buffer via the indicated route over approximately 1 min. In most of the previous immunotargeting studies with radioisotope tracing in mice and rats, uptake of antibodies in organs was determined 1-h postinjection (Muzykantov et al., 1999; Christofidou et al., 2000;Danilov et al., 2001; Scherpereel et al., 2001). To collect data comparable with other animal species, 1 h after injection pigs were hemodiluted just before sacrifice. Hemodilution was performed by withdrawing of blood from the femoral venous catheter while simultaneously infusing lactated Ringers solution through the femoral arterial catheter to maintain isovolemia. The internal organs were dissected, washed with saline, blotted dry, and weighed. Tissue radioactivity in a 2- to 3-g piece of organ tissue and 5-ml samples of blood was determined in a gamma counter (PerkinElmer-Wallac, Gaithersburg, MD). Simultaneous injection of a mixture of both anti-PECAM and control IgG labeled with different isotopes permitted a direct comparison of both conjugates in the same animal, thus allowing an accurate estimation of the specificity of anti-PECAM targeting, while reducing the number of animals needed for the studies.
The results of 125Iodine and131Iodine measurements in the organs were used to calculate three parameters characterizing related, yet different aspects of the anti-PECAM biodistribution and targeting (see Danilov et al., 2001, for detailed explanations). First, the percentage of injected dose in an organ (%ID) measures the total amount of antibody uptake in an organ, showing biodistribution and effectiveness of antibody uptake. Second, the percentage of injected dose per gram of tissue (%ID/g) permits comparisons of the antibody targeting to different organs, evaluating tissue selectivity of an antibody uptake. These two parameters (%ID and %ID/g) provide different absolute values in many organs (including the lung, which weighs approximately 50 g in pigs). Third, the ratio between %ID/g in an organ of interest and that in blood gives the localization ratio (LR). This parameter compensates for the difference in the circulating levels of radiolabeled antibody (e.g., due to different rate of uptake by clearing or target organs) and allows a more objective comparison of targeting between different carriers, with different rates of blood clearance. By dividing the LR of the anti-PECAM conjugate uptake in an organ by that of the IgG, one can calculate an immunospecificity index (ISI = LRmAb/LRIgG, i.e., the ratio between the tissue uptake of immune and nonimmune counterparts normalized to their blood level). ISI is the most objective parameter of the targeting specificity.
Measurement of β-Gal Activity in Vivo.
β-Galactosidase activity in the organ homogenates was determined using a β-Gal enzyme activity assay kit (Promega, Madison, WI) as described previously (Scherpereel et al., 2001). Organs were homogenized in 3 ml of 1× reporter lysis buffer from the kit, containing protease inhibitor cocktail (10 μl/ml; Sigma Chemical) and centrifuged at 4°C, 4000 rpm (3000g) for 45 min. Enzymatic β-Gal activity was determined in the supernatants at various dilutions. The BCA Protein Assay kit (Pierce Chemical) was used to measure the protein concentration in the samples.
Biodistribution and Tissue Localization of β-Gal Conjugates in Pigs.
The tissue localization of enzymatically active β-Gal in mice was studied both with β-Gal activity assay in tissue homogenates and histologically by X-Gal staining as described (Scherpereel et al., 2001). Anesthetized pigs underwent right coronary artery injection of 1 mg of SA-β-Gal, conjugated either to biotinylated anti-PECAM (n = 3) or biotinylated control IgG (n= 3). Three pigs injected with saline were used as controls. Our recent study in mice showed-β-Gal enzymatic activity reaches a transient peak in the lung 15 to 30 min post-i.v. injection (Scherpereel et al., 2001). Thirty minutes after injection of the conjugate, the pigs were bled and perfused through a femoral artery catheter with about 1 liter of sterile saline, the organs harvested, and flash frozen either in reporter lysis buffer for the β-Gal activity assay performed as described above, or in ornithine carbamyl transferase for the X-Gal staining. Frozen sections of tissues were rinsed in PBS, fixed in 0.5% glutaraldehyde, and incubated in X-Gal solution for 4 h at room temperature.
Cellular Localization of Conjugates in Lung and Heart.
The cardiopulmonary block was removed from pigs 30 min postinjection of anti-PECAM/SA-β-Gal, IgG/SA-β-Gal, or saline and perfused with 300 ml of sterile saline then 200 ml of 2% formaldehyde in PBS, pH 7.4, for 15 to 20 min. Visualization of the tissues was performed as described in the previous study (Tzeng et al., 1996; Villanueva et al., 1998). Briefly, small samples were cut and put in 2% formaldehyde in PBS on ice for 1 h then in 10× sucrose in PBS. Frozen sections were incubated in 5% normal goat serum and then washed three times for 5 min by using 0.1 M PBS containing 5% bovine serum albumin and incubated in a cocktail of Alexa 488 conjugated to streptavidin (Molecular Probes, Eugene, OR) and Cy3-conjugated to phalloidin in the same buffer for 1 h. The sections were then washed five times in PBS and mounted without further washing in Gelvatol (Monsanto, St. Louis, MO). To examine the precise distribution of delivered β-galactosidase in cells, the sections were scanned with a 40× plan apochromat objective at 1024 × 1024 pixel resolution in a Leica TCS-NT confocal microscope, with a magnification factor of 2× at the scan head. To maximize z-axis resolution, scans were performed with a small pinhole such that the resolution from a measured point spread function in the z-axis was less than twice the X-Y resolution (0.35 μm). Each scan is the Kalman average of four sequential scans through the midplane of the cells.
Statistics.
Analysis of statistically significant differences (p < 0.05) between groups was performed using a t test or a one-way analysis of variance (SigmaStat 2.0, Tandel Corp., San Rafael, CA). Post hoc testing was performed using the Fischer Least Square difference test. If not otherwise mentioned, all data are expressed as mean ± S.E.M.
Results
Cross-Reactivity of mAb 62 with Swine PECAM-1.
In the first set of experiments, we visualized PECAM in swine tissues sections by using mAb 62, a murine monoclonal generated against purified human PECAM. This antibody has shown broad cross-reactivity with a number of other species (Muzykantov et al., 1999; Nakada et al., 2000). Immunostaining revealed a strong positive reaction in the luminal layer of blood vessels in all organs examined. Figure1 shows that mAb 62, but not control IgG, shows intense staining of vascular endothelium in blood vessels of diverse caliber in porcine lungs (Fig. 1A) and heart (Fig. 1C).

PECAM immunostaining in the porcine cardiopulmonary vasculature. Porcine lung (A and B) and heart (C and D) were incubated with a murine monoclonal antibody directed against human PECAM-1 (anti-PECAM mAb 62; A and C) or control murine IgG (B and D). Original magnifications, 200× (B–D) and 500× (A). Note strong positive reaction in the pulmonary alveolar capillaries, as well as in blood vessels of diverse caliber in the lungs and heart.
Biodistribution of Radiolabeled Anti-PECAM after Intravenous Injection in Pigs.
We characterized the distribution of131I-IgG and125I-anti-PECAM 1 h after i.v. injection in pigs. At this time point, the blood level of control131I-IgG was close to 0.2% ID/g (Fig.2A). Because the blood volume in a 3- to 4-kg pig is approximately 300 ml, roughly 60% of the injected IgG is in the blood. The blood level of 125I-anti-PECAM was significantly lower than that of control131I-IgG (p < 0.01) and did not exceed 0.05% ID/g. This result likely reflects anti-PECAM binding to endothelium (see below) with partial depletion of the circulating pool to approximately 15% of injected dose.
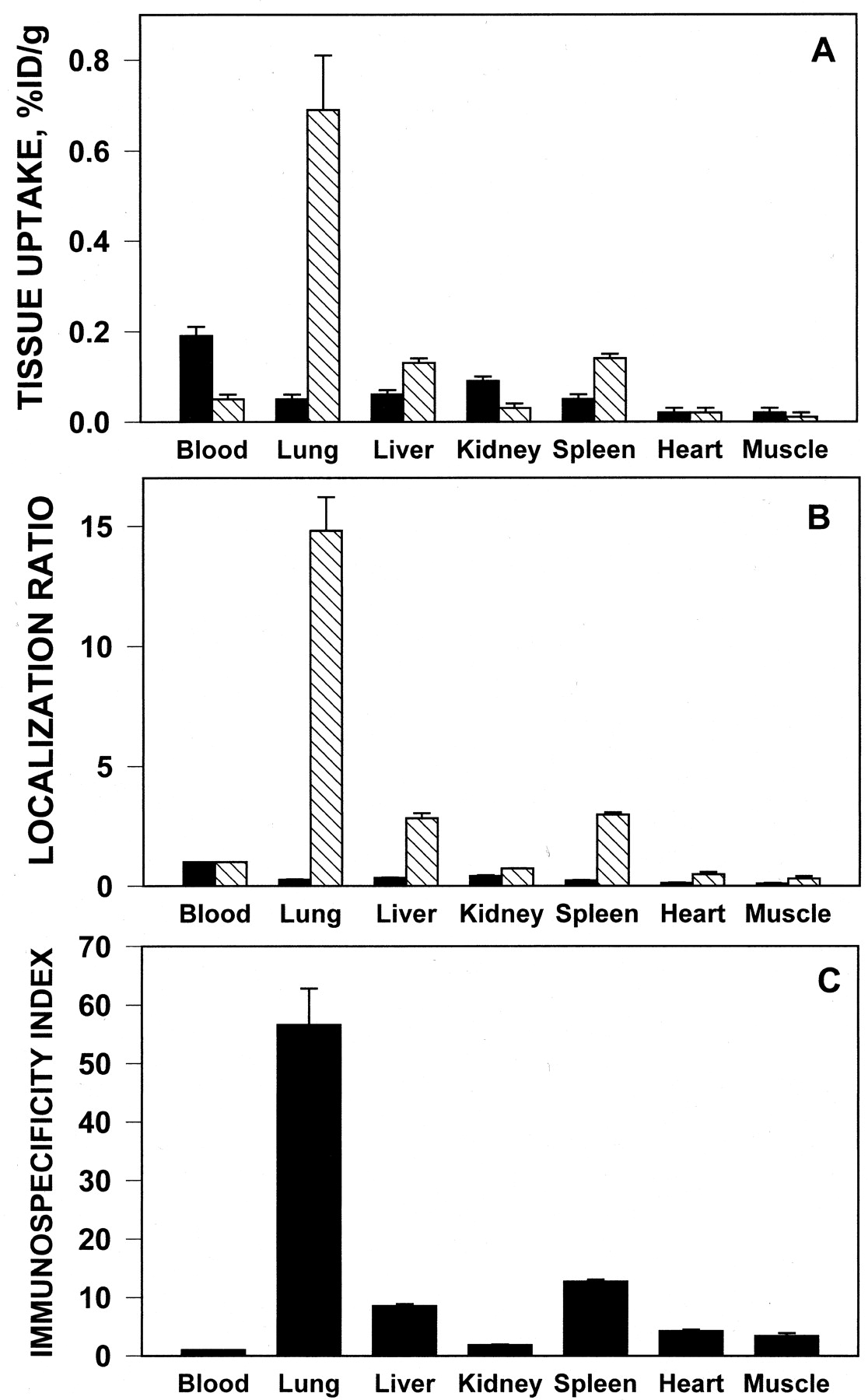
Distribution of 131I-IgG (▪) and anti-PECAM/125I-streptavidin (▧) in the organs 1-h postsystemic intravenous injection in pigs. A, absolute value of the isotopes uptake normalized per weight. B, ratio of radioactivity per gram of tissue to that in blood (localization ratio). C, immunospecificity index of anti-PECAM uptake in the organs calculated as ratio of LR for 125I and 131I. The data are shown as mean ± S.D., n = 3.
The value of 131I-IgG uptake did not exceed 0.1% ID/g in any tissue (Fig. 2A) and the tissue-to-blood ratio (LR) was below 0.5 in all organs (Fig. 2B). The pulmonary uptake of131I-IgG did not exceed 0.05% ID/g and the LR was close to 0.3.
In contrast, the pulmonary uptake of125I-anti-PECAM was close to 0.7% ID/g (Fig.2A), with the localization ratio close to 15 (Fig. 1B). The immunospecificity index of 125I-anti-PECAM pulmonary targeting (ratio between LR of 125I and131I in the lungs) exceeded 50 (Fig. 2C). Thus, the anti-PECAM carrier displayed preferential pulmonary accumulation after i.v. injection in pigs, similar to that reported in rodents (Muzykantov et al., 1999; Scherpereel et al., 2001).
To allow a comparison between large and small organs, the data in Fig.2A are expressed as %ID/g. The total uptake of125I-anti-PECAM, determined as %ID/g multiplied by the organ weight, was moderate (5–7%) in liver and spleen (Table1). As seen with tracing of radiolabeled proteins in vivo in smaller animals, the balance of injected tracers in major organs and blood does not usually add up to 100%. The main reason is that it is difficult to accurately determine uptake in skin and other nonmeasured parts of body. However, the pulmonary uptake of125I-anti-PECAM attains 20% of injected dose post-i.v. injection (an average lung is 25 g in 3-kg pigs). Thus, the lungs display the highest total uptake of125I-anti-PECAM after i.v. injection.
Uptake of radiolabeled anti-PECAM in the organs 1-h postintravenous injection in anesthetized pigs
Effect of Local versus Systemic Route of Intravascular Administration on 125I-Anti-PECAM Uptake in Lung and Heart.
To test the hypothesis that local administration of the conjugate through vascular catheters may facilitate delivery to the endothelium in an organ supplied by that vessel, we compared the uptake of 125I-anti-PECAM in the organs 1 h post-i.v. injection with that postinfusion via a catheter inserted into the right pulmonary artery (RPA) or the right coronary artery (RCA).
Figure 3 shows the levels of125I in the right and left lower lobes of the pig lungs after infusion of 125I-anti-PECAM via i.v., RPA, and RCA routes. Systemic i.v. injection provided an equal uptake of 125I-anti-PECAM in right and left lobes. A similar result was seen with a catheter placed in the RCA. This was not unexpected because blood returning to the heart via the coronary sinus enters the pulmonary artery and encounters the vasculature in all lung lobes during the same passage, hence homogeneously distributing125I-anti-PECAM equally in the lobes. In contrast, local infusion via the right pulmonary artery resulted in heterogeneous 125I-anti-PECAM uptake in the lung lobes: uptake in the right lower lobe (the first-pass vasculature) was 3 times higher than after i.v. delivery (1.98 versus 0.68% ID/g;p < 0.01). In comparison, uptake in the lobes of the left lung was 2-fold lower than after i.v. injection (0.25–0.3 versus 0.7% ID/g). Uptake of 125I-anti-PECAM in the lingular lobe was more variable between animals, varying from 0.25 and 0.51 to 4.9 and 2.2% ID/g. We attribute this to variability of catheter placement relative to the origin of the branch supplying this lobe (which arises as a proximal branch of the right pulmonary artery).

Comparison of anti-PECAM/125I-streptavidin distribution in the lung lobes after the conjugate infusion via intravenous (▪), right pulmonary artery (■), and right coronary artery (▨) routes of administration. The data are shown as mean ± S.D., n = 5.
Further support for a local PECAM-directed targeting paradigm comes from the results of infusion via the coronary artery showing markedly enhanced the cardiac uptake of125I-anti-PECAM. Thus,125I-anti-PECAM uptake in the heart was augmented 4-fold, to 0.08 versus 0.02% ID/g post-i.v. or RPA routes (Fig.4). 131I-IgG infused by any route did not accumulate in the heart and thus the ISI after RCA infusion was close to 10, indicating highly specific, antigen-targeted uptake. The uptake of125I-anti-PECAM in the right atrium after RCA infusion was also high (0.08 ± 0.03% ID/g), whereas that in the left atrium was low (0.03 ± 0.01% ID/g; not significantly different from i.v. or RPA administration).

Cardiac uptake of 131I-IgG (■) and anti-PECAM/125I-streptavidin (▪) 1-h postinfusion via systemic i.v., RPA, and RCA routes of administration. The data for the left ventricle are shown as mean ± S.D., n = 3 (i.v.), 5 (RPA), and 7 (RCA). ∗, difference with other routes and with nonimmune control statistically significant atp < 0.01.
The data shown in Table 2 allow comparison between distribution of 131I-IgG and125I-anti-PECAM in kidney, liver, and spleen after i.v., RPA, and RCA infusion in pigs. Hepatic and splenic uptake of 125I-anti-PECAM was modest using all routes, yet it was lower after infusion via RPA (50% of i.v. level). This result likely reflects more effective depletion of the circulating pool by pulmonary vasculature after local administration of the conjugate. However, the lung was the major target for125I-anti-PECAM by using all routes. Regardless on the pathway of injection, IgG did not accumulate in any organ.
Distribution of radiolabeled IgG and anti-PECAM in pigs 1-h postinfusion via i.v., RPA, or RCA
Delivery of Reporter Enzyme Conjugated with Anti-PECAM Carrier.
To characterize the targeting of a functionally active enzyme to the cardiopulmonary endothelium, we infused streptavidin-β-galactosidase coupled with either IgG or anti-PECAM (IgG/β-Gal and anti-PECAM/β-Gal, 200 μg/kg) in intact pigs. Because the radioisotope data showed that RCA infusion permitted uptake of anti-PECAM in both pulmonary and cardiac vasculature, we used this route in all subsequent experiments.
Table 3 shows β-Gal activity in the organs 30 min after administration of IgG/β-Gal or anti-PECAM/β-Gal. This time point was chosen based on previous studies in mice showing maximal activity near this time point (Scherpereel et al., 2001). Similar to the isotope tracing data, β-Gal activity in liver and spleen was markedly lower than in lungs after injection of anti-PECAM/β-Gal. However, the basal level of hepatic and splenic β-Gal activity (measured in the tissue homogenates harvested from control pigs infused with saline) was markedly higher than in other organs. Therefore, to compensate for this basal level, it has been subtracted from the data for both conjugates. The baseline-corrected ratio between immune and nonimmune counterparts objectively shows a specific elevation of β-Gal activity. Thus, the β-Gal activity in liver and spleen was significantly higher 30 min postinjection of anti-PECAM/β-Gal than that of nonimmune counterpart (4- and 2-fold, respectively). There was no specific augmentation of β-Gal activity in the kidney; both conjugates induced approximately 3-fold elevation of renal β-Gal activity. Most likely, kidneys represent a pathway for the enzyme excretion.
Distribution of β-galactosidase activity in the organs of newborn pigs 30 min after injection of the conjugates into the right coronary artery
IgG/β-Gal infusion led to 2-fold increase in β-Gal activity over the basal level in the lung, probably due to nonspecific uptake in the pulmonary vasculature. In contrast, infusion of the same dose of anti-PECAM/β-Gal provided a 20-fold increase in β-Gal activity in the lungs. Thus, the baseline-corrected ratio of pulmonary β-Gal activity after injection of anti-PECAM/β-Gal versus IgG/β-Gal exceeded 25.
Importantly, RCA administration of anti-PECAM/β-Gal resulted in a 5-fold elevation of the enzyme activity in the heart, whereas IgG/β-Gal induced only 2-fold increase. Therefore, the baseline-corrected ratio of cardiac β-Gal activity after injection of anti-PECAM/β-Gal versus IgG/β-Gal exceeded 5.
Tissue Localization of Anti-PECAM/β-Gal in Cardiopulmonary Vasculature.
To visualize active conjugates in the tissues, organ sections were stained with X-Gal (Fig.5). Intense X-GAL staining was detected in pulmonary vessels after infusion of anti-PECAM/β-Gal, but not IgG/β-Gal. Staining was restricted to the vascular intima: no β-Gal activity was detected in other tissue compartments, including interstitium, airways, and alveolar space. Staining was most frequently associated with alveolar capillaries, yet in addition, staining was often located along the luminal surfaces of arteries and veins of diverse caliber. There was a diffuse faint X-GAL staining in the liver after anti-PECAM/β-Gal infusion; however, it was equivalent to that detected after infusion of control IgG/β-Gal. We also observed positive X-GAL staining in the splenic follicles after injection of either conjugate, suggesting that in this compartment, the conjugates are taken up nonspecifically at a low level. There was some specific X-Gal staining in the renal glomeruli after injection of anti-PECAM/β-Gal, but total staining in the kidney tissue was fairly weak. Importantly, we observed specific X-Gal staining in the cardiac vasculature after RCA infusion of anti-PECAM/β-Gal, but not IgG/β-Gal conjugate.
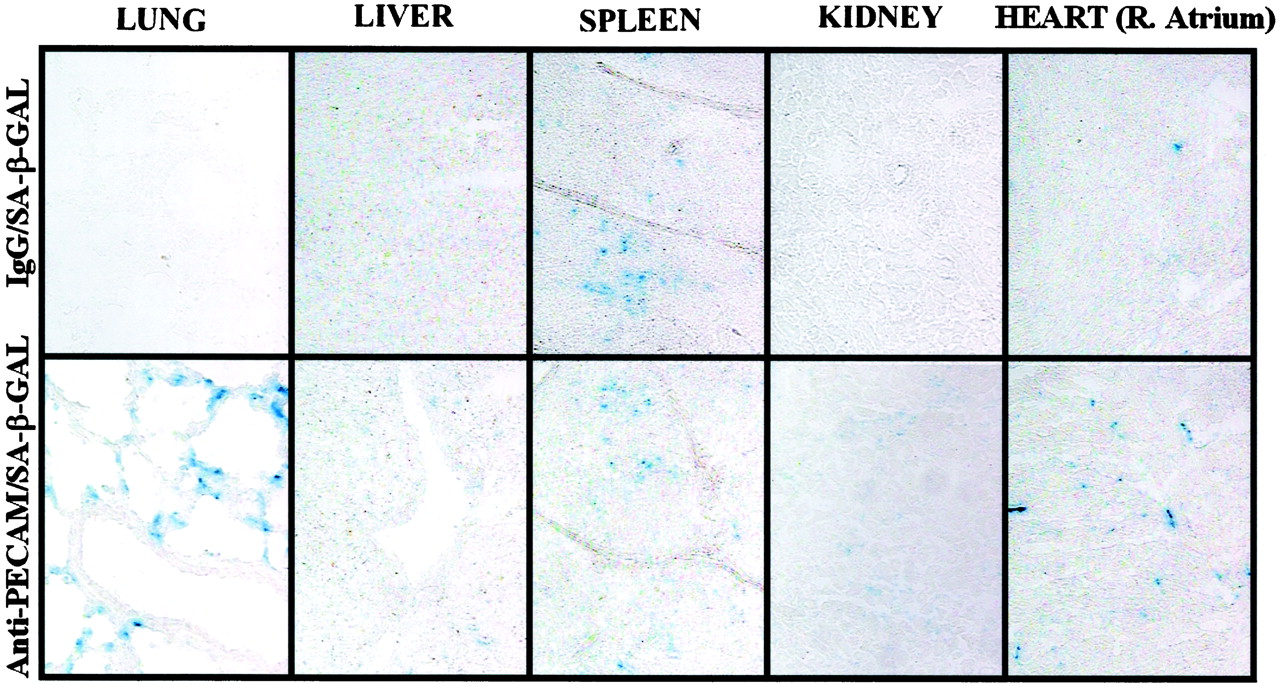
Visualization of β-Gal enzymatic activity in pig organs. Organs were harvested from pigs 30 min after infusion of IgG/β-Gal (left column) or anti-PECAM/β-Gal (right column) conjugates in pigs via the right coronary artery. Right atrium shown for the heart. Tissue sections were exposed to X-GAL and inspected at 200× magnification (except 400× for the lungs).
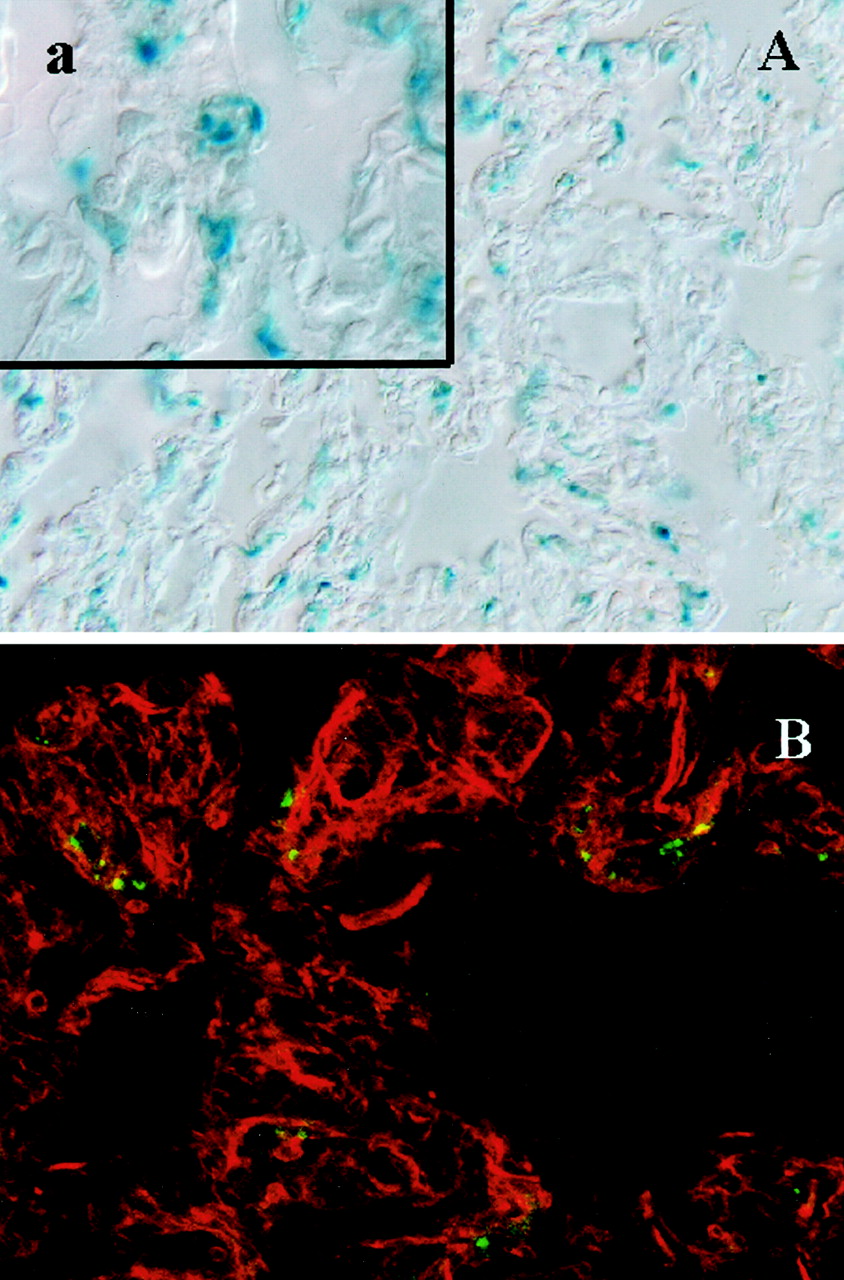
To obtain higher resolution, confocal microscopy was used to visualize anti-PECAM/β-Gal conjugates in formaldehyde-fixed tissue sections by using fluorescent immunostaining with anti-β-gal antibodies. Figure6A shows X-Gal staining and anti-β-Gal-immunostaining (Fig. 6B) in the lung tissue post-RCA infusion of anti-PECAM/β-Gal in pigs. Vascular fluorescence was evident after injection of the conjugate (Fig. 6B). In control animals, no significant FITC fluorescence could be detected (data not shown). Similarly, both X-Gal staining and β-Gal-immunostaining localized the enzyme to small vessel endothelium in the heart post-RCA infusion of anti-PECAM/β-Gal (Fig. 7).

Localization of β-Gal enzyme in the lung tissue 30 min postinfusion of anti-PECAM/β-Gal via the right coronary artery (in anesthetized pigs). A, X-GAL staining in the formaldehyde-fixed lung tissue section (the inset shows higher magnification). B, immunostaining with antibody directed against β-Gal and secondary FITC-labeled antibody inspected in a confocal microscope. Original magnification, 100× (a, 200×). Red color in B shows counterstaining for actin.

Localization of β-Gal enzyme in the heart tissue (right atrium) 30 min postinfusion of anti-PECAM/β-Gal via the right coronary artery (in anesthetized pigs). A, X-GAL staining in the formaldehyde-fixed heart tissue section (the inset shows higher magnification). B, immunostaining with antibody directed against β-Gal and secondary FITC-labeled antibody inspected in a confocal microscope. Magnification, 100× (a, 200×). Red color in B shows counterstaining for actin.
Discussion
Several strategies have been developed to facilitate effective, safe, and specific targeting of therapeutic molecules to endothelial cells. The pulmonary vasculature represents a privileged target for drug delivery either via systemic intravenous or pulmonary artery catheter administration (Muller et al., 1994; Schachtner et al., 1995;Rodman et al., 1997; Danilov et al., 2001). Cationic liposomes and viral particles tend to accumulate in the lungs due to venous blood filtration, thus providing transfection of pulmonary vascular cells, although the specificity and safety of this delivery remain to be rigorously characterized (Channon et al., 1998; Li et al., 1999a,b). Vascular immunotargeting may further enhance drug delivery to the endothelium. This approach permits preferential targeting of enzymes, genes, and viruses to pulmonary endothelium, after systemic intravascular administration in mice and rats (Muzykantov, 1998;Muzykantov et al., 1999; Christofidou et al., 2000; Li et al., 2000;Reynolds et al., 2000; Danilov et al., 2001; Scherpereel et al., 2001). This study validates the utility of this approach in an additional and larger species, the pig.
Delivery of drugs and genes to endothelial cells in extrapulmonary vessels represents even more challenging goal. Several laboratories have evaluated this area by using ligands that recognize specific vascular areas (Huang et al., 1997; Lee et al., 2000). However, unless specific endothelial determinants in an organ of interest are quite specific, uptake of the immunoconjugates in the lung may deplete the circulating pool and thus compromise targeting to the cardiac, renal, and, other regional endothelium. For example, although PECAM is expressed in most vascular beds, intravenous injection leads primarily to pulmonary uptake.
Theoretically, administration of antibody-conjugated drugs via a catheter placed in the vessel of interest could augment local or regional delivery to endothelium in this organ. Results of previous studies, from this and other laboratories, indicate that PECAM-1, a surface adhesion molecule constitutively expressed on endothelial cells in diverse vessels at very high density, is a good candidate target for delivery of enzymes and genes to endothelium (for review, seeMuzykantov, 2001). Thus, an important goal of this study was to test our ability to deliver PECAM conjugates in a larger animal species (pigs) where catheter-based techniques could be used. We therefore compared specific immunotargeting of anti-PECAM conjugates after systemic versus local vascular infusion via catheters placed in various vascular areas.
Our data show that anti-PECAM provides an effective carrier for vascular immunotargeting in diverse animal species ranging from mice to large mammals. The pulmonary vasculature is a primary target for anti-PECAM carriers in all species tested to date, regardless of the route of administration. Effectiveness and specificity of anti-PECAM pulmonary targeting in pigs were equivalent to the best results reported so far with other candidate carriers in rodents. For example, the localization ratio and immunospecificity index achieved in the present study (15 and 55, respectively), match results obtained with another premier antiendothelial carrier, anti-angiotensin-converting enzyme, in rats (Danilov et al., 2001). The dynamic light scattering measurements indicated that both anti-PECAM and IgG conjugates had diameters ranging from 100 to 200 nm (i.e., 50–100 times less than minimal size required for mechanical uptake in the capillaries). However, only anti-PECAM/β-Gal accumulated in the lungs, whereas IgG/β-Gal failed to target the lung or any other organ regardless on the route of administration. Taken together, the data shown in this article indicate that PECAM-directed targeting is due to specific binding of the antibody to endothelial surface.
In the present study, we evaluated the biodistribution of anti-PECAM conjugates in pigs at a relatively short time after intravascular injection by using radiolabeled reagents and enzymatic tracers. In previous studies, we analyzed the stability of radiolabeled immunoconjugates postintravascular injection and found that within at least several hours postinjection, the radiolabeled antibodies and immunoconjugates directed against another endothelial antigen, angiotensin-converting enzyme, circulate as integral proteins and retain their antigen-binding capacity (Muzykantov et al., 1989). Also, the data on the enzymatic activity of anti-PECAM/β-Gal in the tissues (Figs. 5-7) imply that within the observed time interval, the immunoconjugates retain their integrity.
We could not detect specific cardiac uptake of the anti-PECAM conjugate after systemic and right pulmonary artery infusions, probably due to a relatively low binding capacity of cardiac vasculature. According to estimates in diverse animal species, the surface vascular area in the heart is 20 to 50 times less than that in the lung (Crone, 1963; Davis and Hagen, 1993; Panes et al., 1995). After i.v. injection, the pulmonary vasculature represents the endothelial bed encountered first by the injected conjugates. Because swine lung is a large organ with an extensive capillary network, we postulate that the circulating pool of the conjugate is depleted with the lungs, and other organs show lesser uptake (localization ratio was below 1 in all tissues except liver and spleen, where it was close to 2).
However, i.v., RPA, and RCA routes provide markedly different patterns of anti-PECAM distribution within the cardiopulmonary vasculature. PECAM-directed targeting was greatly facilitated when the anti-PECAM conjugate was administered into the arterial supply of the target tissue. Infusion of 125I-anti-PECAM in the coronary artery provided a severalfold enhancement of its cardiac uptake. Although not specifically examined in this study, it is likely that the volume and rate of conjugate infusion may affect its uptake in the target organ. It is tempting to speculate that in the case of regional arterial administration, a slow infusion may provide an additional augmentation of the conjugate uptake in the respective target organ(s).
Both pulmonary and coronary endothelial cells represent important targets for drug delivery. Imaging analysis by using both regular and confocal microscopy indicates that anti-PECAM targeting was directed to vascular endothelial cells in the heart and lungs in pigs. No β-Gal activity was detected in the airway and interstitial compartments. Cell selectivity of anti-PECAM targeting contrasts with results obtained with other strategies such as systemic or RPA infusion of nontargeted liposomal and viral vehicles, which have been reported to deliver their cargoes to extravascular cells (Muller et al., 1994; Schachtner et al., 1995; Rekhter et al., 1998).
Certain therapeutic agents, such as anticoagulants or fibrinolytics, must be retained on the luminal surface of the cells (Muzykantov et al., 1996), whereas therapies with other drugs (e.g., inhibitors of nuclear factors, antioxidants enzymes, prostacyclin synthase, and nitric-oxide synthase) would benefit from intracellular delivery (Muzykantov, 1998, 2001). Our recent studies documented that anti-PECAM conjugates ranging from 100 to 300 nm in diameter enter endothelium in cell culture and in intact mice (Wiewrodt et al., 2002). Investigation of the internalization and intracellular trafficking of the conjugates in pigs would require large quantities of the conjugates and kinetic studies, which were beyond the scope and resources of the present work. However, our data support the possibility of intracellular delivery of therapeutic cargo materials to these important target cells. It is likely that a conjugated agent will affect the rate of internalization and subsequent metabolism of the conjugates. Characterization of the subcellular destination of the conjugated drugs deserves further studies involving electron microscopy.
Visualization of the cardiopulmonary vasculature reveals that not all endothelial cells in the lung and heart show anti-PECAM conjugate uptake, even after right coronary artery administration in pigs (Figs.6 and 7). Experiments in mice revealed that injection of as much as 250 μg of anti-PECAM (10 mg/kg) does not saturate all vascular binding sites (Christofidou et al., 2000; Scherpereel et al., 2001). Approximation of these data to large animals, such as pig, indicates that the dose used in the present study (200 μg/kg) was far below the saturation level. However, an order of magnitude higher dose could be used for functional interventions, experimental or therapeutic.
Such interventions may have a role in the treatment of restenosis or reperfusion injury after coronary interventions through delivery of genes and proteins such as nitric-oxide synthase, vascular endothelial growth factor, antioxidant enzymes (Losordo et al., 1998; Shears et al., 1997; Tzeng et al., 1996; Muzykantov, 2001). Although a variety of animal studies have demonstrated protection against reperfusion injury, the clinical application of such strategies is limited by potential systemic toxicity. Targeted delivery of interleukin-10, adenosine receptor agonists, or other proteins could be an effective means of limiting infarct size in conjunction with coronary interventions (Louttit et al., 1999; McVey et al., 1999; Yang et al., 2000). In ongoing studies, the applicability of this system for the targeting of genetic material is being investigated. Several studies have suggested efficacy of therapeutic angiogenesis with vascular endothelial growth factor and fibroblast growth factor in chronic ischemia (Isner et al., 1996; Laham et al., 2000). Recent work suggests similar effects in myocardial ischemia; however, this has necessitated direct myocardial injection (Losordo et al., 1998; Schwarz et al., 2000). PECAM-directed immunotargeting might allow selective delivery of genes promoting angiogenesis via less invasive means. In ongoing studies, the applicability of this system for the targeting genetic material is being investigated. It is also tempting to hypothesize that this strategy may be applicable to other regional vascular beds, such as the cerebral circulation.
In summary, the results presented in this report demonstrate for the first time that anti-PECAM antibodies can serve as carriers that effectively target active protein to vascular endothelial cells in large animals. The pulmonary endothelium represents a prime target. Selective catheter placement in the pulmonary vasculature augmented the conjugate uptake in target lung lobes, whereas coronary artery infusion resulted in selective uptake in downstream cardiac endothelial cells. This system offers a versatile delivery strategy that may be useful for intracellular targeting of active protein or genes into the vascular endothelial cells. Theoretically, diverse antibody carriers directed against endothelial antigens, including surface adhesion molecules such as intercellular adhesion molecule-1, PECAM-1, and selectins can be used to provide therapeutic cargo with affinity to endothelium (Spragg et al., 1997; Muzykantov, 1998; Villanueva et al., 1998; Harari et al., 1999; Muzykantov et al., 1999; Li et al., 2000).
Footnotes
-
This work was supported by the National American Heart Association (Established Investigator Grant 9640204 to V.R.M. and Initial Investigator Grant SDG 00301920 to M.C.S.), Specialized Center of Research National Institutes of Health (Specialized Center for Research in Atherosclerosis in Acute Lung Injury from National Heart, Lung, and Blood Institute HL 60290, Project 4 to V.R.M. and S.M.A.). A.S. was supported by a grant from French Academy of Medicine (Mitjavile Prize 2000). R.W. is a postdoctoral fellow of the Mildred Scheel Stiftung für Krebsforschung der Deutschen Krebshilfe e. V. (D/98/02288). J.-.C.M. is a fellow from the Spanish Government Research Fellowship. This study was presented as a poster at the American Heart Association Scientific Conference in November 2000.
- Abbreviations:
- PECAM
- platelet-endothelial cell adhesion molecule-1
- mAb
- monoclonal antibody
- SA-β-Gal
- streptavidin-β-galactosidase
- PBS
- phosphate-buffered saline
- ID
- injected dose
- ID/g
- injected dose per gram
- LR
- localization ratio
- ISI
- immunospecificity index
- RPA
- right pulmonary artery
- RCA
- right coronary artery
- FITC
- fluorescein isothiocyanate
- Received October 3, 2001.
- Accepted December 5, 2001.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}