


Abstract
Drug discrimination was used to examine the effects of benzodiazepine (BZ)1 receptor-selective ligands in rhesus monkeys. In diazepam-treated (5.6 mg/kg, p.o.) monkeys discriminating the nonselective BZ antagonist flumazenil (0.32 mg/kg, s.c.), the BZ1-selective antagonist β-carboline-3-carboxylate-t-butyl ester (β-CCt) substituted for flumazenil. The onset of action of β-CCt was delayed with a dose of 5.6 mg/kg β-CCt substituting for flumazenil 2 h after injection. In monkeys discriminating the nonselective BZ agonist midazolam (0.56 mg/kg, s.c.), the BZ1-selective agonists zaleplon (ED50 = 0.78 mg/kg) and zolpidem (ED50 = 1.73 mg/kg) substituted for midazolam. The discriminative stimulus effects of midazolam, zaleplon, and zolpidem were antagonized by β-CCt (1.0–5.6 mg/kg, s.c.), and the effects of zaleplon and zolpidem were also antagonized by flumazenil (0.01–0.32 mg/kg, s.c.). Schild analyses supported the notion of a simple, competitive interaction between β-CCt and midazolam (slope = −1.08; apparent pA2 = 5.41) or zaleplon (slope = −1.57; apparent pA2 = 5.49) and not between β-CCt and zolpidem. Schild analyses also were consistent with a simple, competitive interaction between flumazenil and zaleplon (slope = −1.03; apparent pA2 = 7.45) or zolpidem (slope = −1.11; apparent pA2 = 7.63). These results suggest that the same BZ receptor subtype(s) mediate(s) the effects of midazolam, zolpidem, and zaleplon under these conditions and that selective binding of BZ ligands does not necessarily confer selective effects in vivo.
Drugs facilitating GABAA-mediated chloride flux (e.g., agonists; positive GABAA modulators) at benzodiazepine (BZ) receptors elicit sedative-hypnotic, anxiolytic, muscle relaxant, and anticonvulsant effects with an improved margin of safety compared with other positive GABAAmodulators (e.g., barbiturates; for review, see Woods et al., 1992). Different BZ receptors comprise at least one of six different protein subunits designated α1–6 and are divided into two subtypes: BZ1 receptors containing α1-subunits and BZ2receptors containing α2-, α3-, or α5-subunits (Sanger et al., 1994 for review). A more recent nomenclature adopted by the International Union of Pharmacology proposes an alternative designation for BZ receptors (Barnard et al., 1998). It has been postulated that the various effects elicited by BZ site ligands are mediated by different BZ receptor subtypes. This notion has been supported by transgenic mouse studies showing that mutations targeted at α2 subunits decrease the anxiolytic effects of BZs, whereas mutations targeted at α1subunits decrease the sedative-hypnotic effects of BZs (Rudolph et al., 1999; McKernan et al., 2000).
Most BZ receptor ligands have relatively little selectivity for particular BZ receptor subtypes, making it difficult to differentiate the functional significance of BZ receptor heterogeneity. BZs such as diazepam and midazolam are nonselective (Huang et al., 2000), whereas non-BZ compounds such as zaleplon and zolpidem are approximately 10-fold selective for BZ1 receptors compared with other GABAA receptors in binding assays in vitro (Dämgen and Lüddens, 1999; Huang et al., 2000). Selectivity at BZ1 receptors might be responsible for zaleplon and zolpidem having behavioral effects that differ from those of nonselective BZs. For example, in rodents, zaleplon and zolpidem preferentially elicit sedative-hypnotic effects and do not share discriminative stimulus effects with other positive GABAA modulators (Depoortere et al., 1986; Sanger and Zivkovic, 1986; Sanger et al., 1987, 1996; Vanover and Barrett, 1994; Ator and Kautz, 2000 for exception). However, in primate species, zaleplon and zolpidem share discriminative stimulus and subject-rated effects with other positive GABAA modulators (Griffiths et al., 1992; Rush and Griffiths, 1996; Rowlett and Woolverton, 1997; Rush et al., 1997, 1999; Rowlett et al., 1999, 2000for exceptions; Ator, 2000). Among antagonists (e.g., neutral GABAA modulators) at BZ receptors, β-carboline-3-carboxylate-t-butyl ester (β-CCt) is approximately 20-fold selective for BZ1 receptors (Huang et al., 2000). Receptor selectivity could be responsible for the ability of β-CCt to antagonize some (e.g., anxiolytic, sedative, anticonvulsant, and discriminative stimulus effects) but not other (e.g., muscle relaxation) effects of nonselective BZ agonists (Shannon et al., 1984, 1988; Griebel et al., 1999; Rowlett et al., 2001).
It is not clear to what extent selectivity for BZ1 receptors differentiates zaleplon, zolpidem, and β-CCt from nonselective BZ receptor ligands. To date, the literature has provided only limited evidence to support the selectivity of certain compounds for BZ receptor subtypes. The general goal of this study was to use a well established, quantitative pharmacologic procedure (Schild analysis) to specifically test whether selective binding that occurs in vitro confers selective effects in vivo. Specifically, two different drug discrimination procedures were used to compare zaleplon, zolpidem, and β-CCt with nonselective BZs in rhesus monkeys. In one procedure, β-CCt was administered to diazepam-treated (5.6 mg/kg/day, p.o.) monkeys discriminating the nonselective BZ antagonist flumazenil (0.32 mg/kg, s.c.). In a second procedure, monkeys discriminating midazolam (0.56 mg/kg s.c.) received β-CCt in combination with midazolam, zaleplon, or zolpidem or flumazenil in combination with zaleplon or zolpidem. Schild analyses were used to confirm whether interactions were simple and competitive and to determine whether antagonist potency was the same or different among the different agonists, i.e., whether agonists were acting at the same or different BZ receptors.
Materials and Methods
Subjects.
Four adult female (midazolam discrimination) and three adult male (flumazenil discrimination) rhesus monkeys (Macaca mulatta) were housed individually on a 14-h light/10-h dark schedule, maintained at 95% free-feeding weight (range = 3.8–11.5 kg) with a diet comprising primate chow (High Protein Monkey Diet; Harlan Teklad, Madison, WI), fresh fruit, and peanuts, and provided water in the home cage. Monkeys were trained previously to discriminate midazolam or flumazenil (Gerak and France, 1999; Lelas et al., 2000; McMahon et al., 2001). The animals used in these studies were maintained in accordance with the Institutional Animal Care and Use Committee, The University of Texas Health Science Center at San Antonio, and with the 1996 Guide for the Care and Use of Laboratory Animals (Institute of Laboratory Animal Resources on Life Sciences, National Research Council, National Academy of Sciences).
Apparatus.
During experimental sessions, monkeys were seated in chairs (model R001; Primate Products, Miami, FL) that provided restraint at the neck and placed in ventilated, sound-attenuating chambers equipped with two response levers, stimulus lights, and a food cup to which pellets (Bio-Serv, Frenchtown, NJ) could be delivered from a dispenser. For monkeys discriminating midazolam under a schedule of stimulus-shock termination, feet were placed in shoes containing brass electrodes through which a brief electric stimulus (3 mA, 250 ms) could be delivered from an a.c. generator. An interface (MedAssociates, St. Albans, VT) connected the chambers to a computer that controlled and recorded experimental events.
Flumazenil Discrimination Procedure.
Diazepam was given 3 h prior to experimental sessions. Experimental sessions consisted of multiple 15-min cycles each comprising a 10-min timeout period, during which responses had no programmed consequence, followed by a 5-min response period, during which green stimulus lights were illuminated and a fixed ratio (FR)5 schedule of food presentation was in effect. A maximum of 10 food pellets was available during a cycle; when the maximum number of food pellets was obtained in less than 5 min, the remainder of the response period was a timeout. The selection of vehicle- and flumazenil-appropriate levers varied among monkeys and remained the same for an individual throughout the study. Responding on the incorrect lever reset the response requirement on the correct lever.
Vehicle training comprised administration of vehicle or sham injections during the first minute of each cycle, of no more than eight cycles. Flumazenil training sessions comprised administration of flumazenil (0.32 mg/kg, s.c.) during the first minute of a cycle, followed by a vehicle or sham injection during the first minute of a second cycle. Test sessions were conducted following training sessions in which ≥80% of the total responses occurred on the lever designated correct by the injection administered during the first minute of the cycle and fewer than five responses occurred on the incorrect lever prior to completion of the FR response requirement on the correct lever. Prior to each test, these criteria had to be satisfied for training sessions during which both flumazenil and vehicle or sham injections were administered. The type of training session preceding test sessions varied nonsystematically. Test sessions were identical to training sessions except that five consecutive responses on either lever resulted in food delivery. Cumulative flumazenil dose-effect tests were conducted by injecting the appropriate vehicle solution during the first minute of the first cycle, followed by increasing doses of flumazenil during the first minute of subsequent cycles, with the cumulative dose increasing by 0.25 or 0.5 log unit per cycle. Test sessions ended when ≥80% of the total responses occurred on the flumazenil-appropriate lever or when response rate decreased to less than 20% of control response rate. Single doses of β-CCt (3.2–10 mg/kg) were administered on separate occasions at the beginning or 2 h prior to eight sham injection cycles. The largest dose of β-CCt (10 mg/kg) was not administered at the 2-h pretreatment interval due to a limited supply of this compound.
Midazolam Discrimination Procedure.
Multiple cycle procedures were similar to those described above except that the 5-min response period comprised an FR5 schedule of stimulus-shock termination. Illumination of red stimulus lights located above each lever signaled the beginning of the response period in which shock was to be delivered every 15 s. Five consecutive responses on the lever designated correct by the injection administered during the first minute of the cycle extinguished the stimulus lights and postponed the shock schedule for 30 s. Responding on the incorrect lever reset the response requirement on the correct lever.
Test sessions were conducted when animals satisfied the criteria specified above for monkeys discriminating flumazenil. Cumulative midazolam, zaleplon, or zolpidem dose-effect tests were conducted by injecting the appropriate vehicle solution during the first minute of the first cycle followed by increasing doses of the respective test compound during the first minute of subsequent cycles, with the cumulative dose increasing by 0.25 or 0.5 log unit per cycle. Test sessions ended when ≥80% of the total responses occurred on the midazolam-appropriate lever or when response rate decreased sufficiently to result in the delivery of more than two shocks. On separate occasions, a dose of β-CCt (1–5.6 mg/kg) or flumazenil (0.01–0.32 mg/kg) was administered followed by cumulative doses of a test compound. Flumazenil was administered during the first cycle, and β-CCt was administered 2 h before administration of the agonists, the time at which 5.6 mg/kg β-CCt substituted for flumazenil in diazepam-treated monkeys.
Drugs.
The vehicle for oral administration of diazepam was fruit punch combined with suspending agent K (Bio-Serv) in a concentration of 1 g of suspending agent per liter of fruit punch. Tablets containing 10 mg of diazepam (Zenith Laboratories, Inc., Northvale, NJ) were dissolved in vehicle, mixed in a blender, and administered using a 12-G drinking needle attached to a 60-cc syringe. To obtain a dose of 5.6 mg/kg diazepam, a standard concentration of diazepam was given in a volume adjusted to individual body weights. The diazepam mixture was prepared immediately before administration.
The following drugs were administered s.c. in a volume of 0.01–0.1 ml/kg b.wt. expressed in terms of the forms listed below: β-CCt (synthesized by J.M.C.; Cox et al., 1995), flumazenil (F. Hoffmann LaRoche Ltd., Basel, Switzerland), midazolam hydrochloride (Roche Pharma Inc., Manati, Puerto Rico), zolpidem (Synthelabo Recherché, Bagneux Cedex, France), and zaleplon (Wyeth-Ayerst Research, St. Davids, PA). Midazolam was purchased as a commercially prepared solution in a concentration of 5 mg/ml and diluted with saline. β-CCt and zaleplon were dissolved in a vehicle comprising 50% ethanol and 50% Emulphor. Flumazenil was dissolved in a vehicle comprising 40% propylene glycol (Sigma Chemical, St. Louis, MO), 50% saline, and 10% ethanol. Zolpidem was suspended in 5% Tween 80 (Sigma) in sterile water.
Data Analyses.
Drug discrimination data are expressed as the percentage of total responses occurring on the drug-appropriate lever averaged among monkeys (±S.E.M.) and plotted as a function of dose. Substitution for the training drug was defined as ≥80% responding on the drug-appropriate lever. When a test with a given compound was conducted more than once, the determinations were averaged for an individual subject for further analyses. Doses of a compound required to produce 50% drug-appropriate responding (ED50) and the 95% confidence limits (CLs) were estimated using linear regression by using more than two appropriate data points, otherwise by interpolation. ED50values for midazolam, zaleplon, and zolpidem following administration of β-CCt or flumazenil (zaleplon and zolpidem only) were compared with the average of control ED50 values for the respective test compounds. The magnitude of rightward shift elicited by β-CCt or flumazenil was determined first for individual monkeys and then averaged among all monkeys. For β-CCt or flumazenil antagonism of BZ receptor ligands, pA2 analyses were carried out with the Pharm/PCS Pharmacologic Calculation System (version 4.2) based on Tallarida and Murray (1987). Slopes of Schild plots were considered to conform to unity when 95% confidence limits included −1 and did not include 0 (e.g., Paronis and Bergman, 1999).
Control response rate represents the average of the five vehicle training sessions before the test. Response rate was calculated as a percentage of control rate for individual animals, then averaged among subjects (±S.E.M.) and plotted as a function of dose.
Results
Substitution of β-CCt for Flumazenil in Diazepam-Treated Monkeys.
Administration of vehicle occasioned primarily vehicle-lever responding for 3 h and 45 min after injection (data not shown). A dose of 3.2 mg/kg β-CCt occasioned primarily vehicle-appropriate responding for 3 h and 45 min after injection in three monkeys (Fig. 1, top panel). A larger dose (5.6 mg/kg) of β-CCt occasioned a time-related increase to 70 to 90% flumazenil-appropriate responding beginning 2 h after injection of β-CCt in three monkeys. A still larger dose (10 mg/kg) of β-CCt occasioned a time-related increase to ≥80% flumazenil-lever responding beginning 30 min after injection in two monkeys. The β-CCt dose-effect function (Fig.2, top panel) was constructed from the percentage of flumazenil-lever responding observed 1 h after injection. β-CCt substituted for flumazenil with an ED50 (95% CL) of 6.15 (5.00, 7.41) mg/kg. Administration of the flumazenil vehicle solution occasioned predominantly vehicle-appropriate responding, whereas cumulative doses of flumazenil dose-dependently increased responding on the flumazenil-appropriate lever with a cumulative dose of 0.1 producing ≥80% flumazenil-lever responding (Fig. 2, top panel). The ED50 (95% CL) for the flumazenil-discriminative stimulus was 0.05 (0.03, 0.07) mg/kg.

Time course of discriminative stimulus and rate effects of β-CCt in diazepam-treated monkeys discriminating flumazenil. Abscissae: time in min. Ordinates: mean (±S.E.M.) percentage of responding on the drug-appropriate lever (%DR = drug responding, top) and mean response rate expressed as percentage of control (vehicle training days) rate [rate (% of control), bottom]. S.E.M. values for mean response rate are omitted for clarity. Data represent average values from three monkeys for 5.6 mg/kg (■) and two monkeys for 3.2 (○) and 10 (⋄) mg/kg.
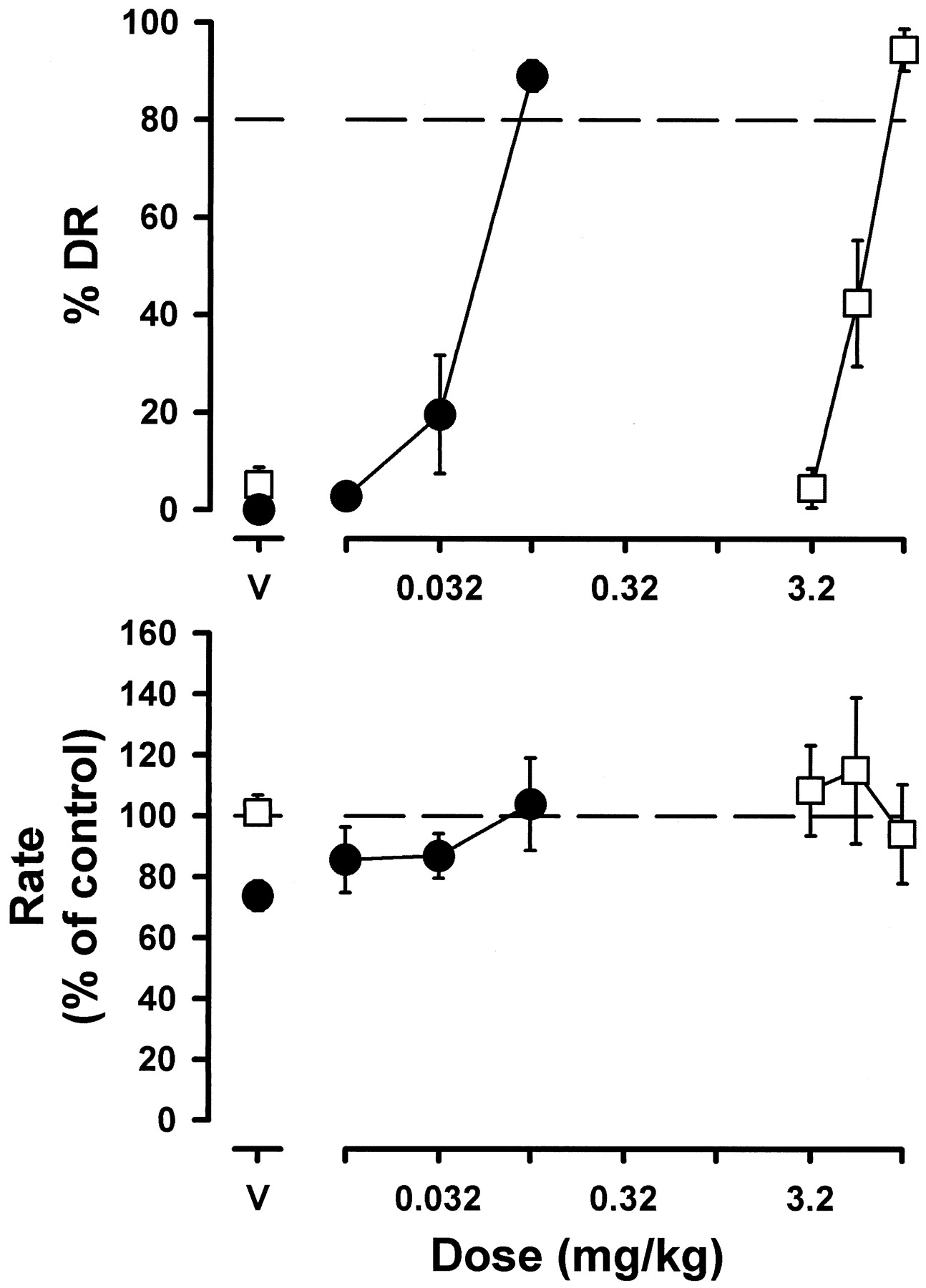
Discriminative stimulus and rate effects of β-CCt (■) and flumazenil (•) in diazepam-treated monkeys discriminating flumazenil. Abscissae: dose in milligrams per kilogram of body weight; V, vehicle. Ordinates: mean (±S.E.M.) percentage of responding on the drug-appropriate lever (%DR = drug responding, top) and mean response rate expressed as percentage of control (vehicle training days) rate [rate (% of control), bottom]. Data for β-CCt are redrawn from 1 h in Fig. 1. Vehicle and flumazenil data represent average values from three monkeys.
Under control conditions, the mean response rate (±S.E.M.) was 1.10 ± 0.04 responses per second. β-CCt (3.2–10 mg/kg), flumazenil (0.01–0.32 mg/kg), and their respective vehicle solutions did not substantially alter response rate (Figs. 1 and 2, bottom panels).
Substitution of Zaleplon and Zolpidem for Midazolam: Antagonism with β-CCt or Flumazenil.
Cumulative doses of midazolam increased midazolam-appropriate responding in a dose-related manner; similarly, zaleplon and zolpidem substituted (≥80% midazolam-lever responding) for midazolam (Fig. 3through 7, top panels; ●). The order of potency [ED50 values (95% CLs)] for midazolam-lever responding was midazolam [0.20 (0.14, 0.26) mg/kg] > zaleplon [0.78 (0.39, 1.09) mg/kg] ≥ zolpidem [1.73 (0.83, 2.83) mg/kg]. The vehicle solutions for each positive GABAAmodulator occasioned predominantly saline-appropriate responding (Figs.3-7, top panels).

Discriminative stimulus and rate effects of midazolam alone and in combination with β-CCt in monkeys discriminating midazolam. β-CCt (1, 3.2, or 5.6 mg/kg) was administered 1 h 45 min prior to vehicle (V) and 2 h prior to the administration of the first dose of midazolam. Data represent average values from three monkeys. See Fig. 2 for other details.

Discriminative stimulus and rate effects of zolpidem alone and in combination with flumazenil in monkeys discriminating midazolam. Data represent average values from three monkeys. See Fig. 2for other details.
β-CCt dose-dependently antagonized the midazolam discriminative stimulus with doses of 1, 3.2, and 5.6 mg/kg β-CCt shifting the midazolam dose-effect function 2-, 4-, and 7-fold to the right, respectively (Fig. 3, top panel). β-CCt dose-dependently antagonized the midazolam-like discriminative stimulus effects of zaleplon with doses of 1, 3.2, and 5.6 mg/kg β-CCt shifting the zaleplon dose-effect function 2-, 8-, and 21-fold to the right (Fig.4, top panel). β-CCt was comparatively less effective in antagonizing the midazolam-like discriminative stimulus effects of zolpidem (Fig. 5, top panel); doses of 1, 3.2, and 5.6 mg/kg β-CCt shifted the zolpidem dose-effect function 2-, 2-, and 3-fold to the right.

Discriminative stimulus and rate effects of zaleplon alone and in combination with β-CCt in monkeys discriminating midazolam. Data represent average values from three monkeys. See Fig. 2for other details.

Discriminative stimulus and rate effects of zolpidem alone and in combination with β-CCt in monkeys discriminating midazolam. Data represent average values from three monkeys. See Fig. 2for other details.
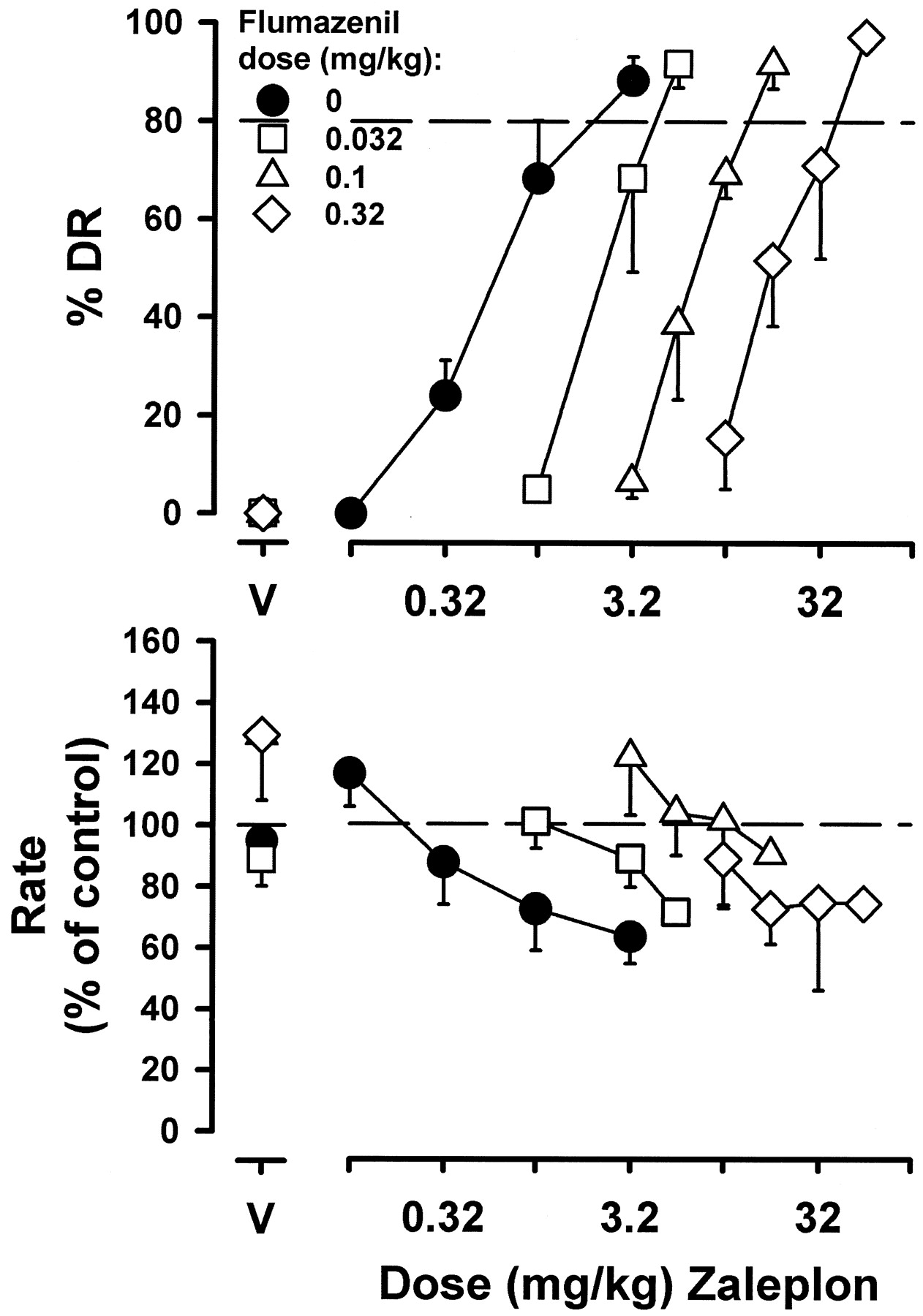
Flumazenil dose-dependently antagonized the midazolam-like discriminative stimulus effects of zaleplon with doses of 0.032, 0.1, and 0.32 mg/kg flumazenil shifting the zaleplon dose-effect function 4-, 11-, and 33-fold to the right (Fig.6, top panel). Flumazenil also antagonized the midazolam-like discriminative stimulus effects of zolpidem with doses of 0.01, 0.032, and 0.1 mg/kg flumazenil shifting the zolpidem dose-effect function 2-, 6-, and 20-fold to the right (Fig. 7, top panel).

Discriminative stimulus and rate effects of zaleplon alone and in combination with flumazenil in monkeys discriminating midazolam. Flumazenil (0.032, 0.1, or 0.32 mg/kg) was administered during the first cycle 15 min prior to the first dose of zaleplon. Data represent average values from three monkeys. See Fig. 2 for other details.
Figure 8 depicts a Schild plot for β-CCt antagonism of midazolam, zaleplon, and zolpidem and for flumazenil antagonism of zaleplon and zolpidem; Table1 depicts the coefficient of determination (r2), slope (95% CL), and, when appropriate, the apparent pA2 value (±95% CL) for each antagonist-agonist combination. The slopes for β-CCt in combination with midazolam or zaleplon were not significantly different from unity, yielding apparent pA2 values of 5.41 and 5.49, respectively. In contrast, the slope for β-CCt in combination with zolpidem was significantly different from unity, thereby precluding determination of an apparent pA2 value (Table 1). The slopes for flumazenil in combination with zaleplon or zolpidem were not significantly different from unity, yielding apparent pA2 values of 7.45 and 7.63, respectively (Table1).

Schild plots constructed from the mean data shown in Figs. 3-7. Abscissae: negative logarithm of the dose of antagonist in moles per kilogram. Ordinates: logarithm of the dose ratio −1.
Results of Schild analyses for combinations of BZ receptor antagonists and agonists in rhesus monkeys (n = 3)
Under control conditions, the mean response rate (±S.E.M.) was 1.81 ± 0.23 responses per second. Midazolam, zaleplon, and zolpidem decreased rate of responding in a dose-related manner (Figs.3-7, bottom panels; ●). β-CCt and flumazenil appeared to antagonize the rate-decreasing effects of zaleplon and zolpidem. ED50 values were not determined for rate-decreasing effects because at the doses studied, each agonist failed to decrease response rate to below 50% of the control response rate in all monkeys.
Discussion
The agonists zaleplon and zolpidem and the antagonist β-CCt bind selectively to BZ1 receptors, although the functional consequences of this selective binding are not fully established (Huang et al., 2000; Rowlett et al., 2000). The current study examined the selectivity of these ligands in one group of monkeys discriminating midazolam (0.56 mg/kg) and in another group of diazepam-treated (5.6 mg/kg/day) monkeys discriminating flumazenil (0.32 mg/kg). The putative BZ1 subtype-selective antagonist β-CCt substituted for the nonselective BZ antagonist flumazenil in diazepam-treated monkeys. The putative BZ1 subtype-selective agonists zaleplon and zolpidem substituted for midazolam in untreated monkeys. In midazolam-discriminating monkeys, β-CCt shifted the midazolam, zaleplon, and zolpidem dose-effect curves to the right. Schild analysis supported the notion of a simple, competitive interaction between β-CCt and midazolam and zaleplon and not between β-CCt and zolpidem. Similarly, flumazenil shifted the zaleplon and zolpidem dose-effect curves to the right and Schild analysis confirmed the notion of a simple, competitive interaction between flumazenil and each of the BZ1 subtype-selective agonists. These results fail to provide support for the notion that zaleplon, zolpidem, and β-CCt act selectively at BZ1 receptors.
Various studies in transgenic mice suggest that different receptor subtypes selectively mediate anxiolytic and sedative-hypnotic effects of ligands acting at GABAA BZ receptors, although the interpretation of these data is not without controversy (Rudolph et al., 1999; McKernan et al., 2000). Clear evidence for functional differences among BZ receptor subtypes could dramatically impact the use of BZ receptor-selective ligands in medicine by providing a framework within which drugs could be developed toward specific receptors and specific therapeutic targets. However, today most BZ receptor ligands have relatively little selectivity for particular BZ receptor subtypes and they often produce a host of both preferred and nonpreferred effects.
Substitution of β-CCt for the flumazenil-discriminative stimulus suggests that these antagonists have qualitatively similar effects in diazepam-treated monkeys. The flumazenil-discriminative stimulus in diazepam-treated monkeys appears to be related to antagonism of chronic diazepam (e.g., precipitated withdrawal) and is comparable to spontaneous withdrawal insofar as temporary suspension of diazepam treatment elicits signs of withdrawal accompanied by responding on the flumazenil-appropriate lever (Gerak and France, 1999). In addition, substitution for flumazenil occurs with other drugs that antagonize diazepam at BZ receptors [e.g., Ro 14-4513] or that functionally antagonize diazepam at other sites on the GABAAreceptor complex (e.g., pentylenetetrazol; Gerak and France, 1999). The similar although not identical binding profile of β-CCt and flumazenil at BZ receptors suggests that β-CCt antagonizes chronic diazepam at BZ receptors to precipitate withdrawal (Huang et al., 2000). β-CCt is approximately 100-fold less potent than flumazenil in diazepam-treated monkeys, a difference comparable with that observed in untreated monkeys (see below; Paronis et al., 2001). β-CCt also had a slower onset of action in diazepam-treated monkeys than the onset of action previously reported for flumazenil (Gerak and France, 1999).
The BZ1 receptor-selective ligands zaleplon and zolpidem substituted for midazolam. These data corroborate the general finding that BZ1 receptor-selective ligands substitute for other positive GABAA modulators in primates including BZs and barbiturates (Griffiths et al., 1992; Rush and Griffiths, 1996; Rowlett and Woolverton, 1997; Rush et al., 1997,1999; Rowlett et al., 1999, 2000 for exceptions; Ator, 2000). Similarities among zaleplon, zolpidem, and nonselective BZs in primates contrast with differences among these ligands in rodents. For example, zaleplon and zolpidem do not consistently cross-substitute with nonselective BZs and other positive GABAAmodulators (Depoortere et al., 1986; Sanger and Zivkovic, 1986; Vanover and Barrett, 1994; Sannerud and Ator, 1995; Ator and Kautz, 2000 for exception). Moreover, zaleplon and zolpidem are more potent in behavioral assays predictive of sedative activity as compared with other types of assays (Depoortere et al., 1986; Sanger and Zivkovic, 1988; Sanger et al., 1996). The apparently unique behavioral profile of zaleplon and zolpidem in rodents has been attributed to the selectivity of these drugs for BZ1 receptors.
The receptor selectivity of these drugs was examined by comparing the ability of β-CCt or flumazenil to antagonize zaleplon, zolpidem, or midazolam. Schild analysis was used to characterize whether drug interactions were simple and competitive and to provide in vivo apparent pA2 values, or estimates of theKB of an antagonist, when appropriate. β-CCt antagonism of midazolam was orderly and dose-related, yielding a high regression coefficient. Schild analysis supported the notion of a simple, competitive interaction between β-CCt and midazolam, yielding an apparent pA2 value of 5.41. This result differs somewhat from previous studies demonstrating β-CCt antagonism of midazolam and diazepam (Shannon et al., 1988; Paronis and Bergman, 1999). One important variable that might account for differences between the present and previous studies is the pretreatment interval used for β-CCt administration. β-CCt was administered 2 h prior to tests because the doses of β-CCt that were studied in combination with midazolam, zaleplon, and zolpidem did not substitute for flumazenil until 2 h after injection. One assumption underlying the use of Schild analysis is that measurements be taken at time of peak effect (Arunlakshana and Schild, 1959). Thus, different pretreatment times could account for differences in the magnitude of antagonism obtained between this and previous studies.
β-CCt also antagonized zaleplon and zolpidem, although the antagonism of zolpidem was not as uniform as that of zaleplon. β-CCt antagonism of zaleplon was orderly and dose-related, yielding a high regression coefficient. Schild analysis supported the notion of a simple, competitive interaction between β-CCt and zaleplon, yielding an apparent pA2 value of 5.49. In contrast, β-CCt antagonism of zolpidem was less orderly and dose-related, yielding a relatively low regression coefficient. Schild analysis of these data revealed a slope that deviated from unity (i.e., the upper and lower limits of the 95% CLs included positive and negative values), suggesting that effects of β-CCt and zolpidem were not the result of a simple, competitive interaction. The similarity in pA2 values for β-CCt in combination with midazolam or zaleplon indicates that the effects of these agonists are mediated by the same BZ receptors. The deviation of unity in the regression line for β-CCt in combination with zolpidem might suggest that β-CCt and zolpidem interact with more than one BZ receptor subtype.
The results with β-CCt suggest that the mechanism of action for zolpidem might not be identical to the mechanism of action for zaleplon and midazolam. However, results with flumazenil in combination with the different agonists suggest otherwise. Flumazenil antagonism of zolpidem was orderly and dose-related, yielding a high regression coefficient. Schild analysis supported the notion of a simple, competitive interaction between flumazenil and zolpidem, yielding an apparent pA2 value of 7.63. Schild analysis also supported the notion of a simple, competitive interaction between flumazenil and zaleplon, yielding an apparent pA2 value of 7.45. These pA2 values are similar to those previously reported for flumazenil in combination with midazolam using identical procedures (pA2 = 7.83; Lelas et al., 2000), thereby indicating that the effects of midazolam, zaleplon, and zolpidem are mediated by the same BZ receptors. The difference in potency between flumazenil and β-CCt in antagonizing the discriminative stimulus effects of agonists in untreated monkeys that discriminate midazolam was the same (100-fold) as the difference in potency between these antagonists in producing discriminative stimulus effects in diazepam-treated monkeys.
Drug discrimination has been used to examine flumazenil antagonism of midazolam, zaleplon, or zolpidem in other studies (Spealman, 1985;Rowlett et al., 1999; Ator, 2000; Lelas et al., 2000), whereas β-CCt has been studied less extensively in drug discrimination assays (e.g.,Shannon et al., 1988). The present study systematically compared BZ1 receptor-selective ligands to nonselective BZ receptor ligands under the same procedures. Other behavioral paradigms comparing the effects of these and other BZ receptor ligands suggest that receptor selectivity is responsible for differences among these ligands (cf., Paronis et al., 2001; Rowlett et al., 2001). Substitution of BZ1 receptor-selective ligands for nonselective BZs in the present study could be interpreted as evidence that BZ1 receptors play an important role in the discriminative stimulus effects of nonselective BZs. However, the selectivity of zaleplon, zolpidem, and β-CCt for BZ1 receptors is less than 30-fold in binding assays (Dämgen and Lüddens, 1999; Huang et al., 2000), and the results of Schild analyses are not consistent with different BZ receptor subtypes mediating the effects of midazolam, zaleplon, and zolpidem. Therefore, despite marginal binding selectivity for BZ1 receptors in vitro, the behavioral effects of zaleplon, zolpidem, and β-CCt under these conditions do not appear to result from selective actions at BZ1 receptors. Other ligands with greater selectivity for BZ1receptors will be required to determine whether BZ1 receptors selectively elicit some (e.g., sedation) and not other BZ effects (e.g., anxiolysis; Rudolph et al., 1999; McKernan et al., 2000).
Acknowledgments
We thank B. Engelhardt, A. Gaylor, and S. Tucker for providing technical assistance.
Footnotes
-
This research was supported by National Institute on Drug Abuse Grant DA09157. C.P.F. is the recipient of a Research Scientist Development Award (DA00211).
- Abbreviations:
- GABAA
- γ-aminobutyic acidA
- BZ
- benzodiazepine
- β-CCt
- β-carboline-3-carboxylate-t-butyl ester
- CL
- confidence limit
- FR
- fixed ratio
- Received August 7, 2001.
- Accepted October 19, 2001.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}