


Abstract
Cell-mediated immune response and release of cytokines after the administration of 3,4-methylenedioxymethamphetamine (MDMA, “ecstasy”) alone and in combination with ethanol were assessed in a double blind, randomized, crossover, controlled clinical trial. Six healthy male recreational users of MDMA participated in four different experimental sessions, with a washout interval between sessions of 1 week, in which single oral doses of MDMA (100 mg), ethanol (0.8 g/kg), the combination of both drugs, and placebo were tested. Acute MDMA administration produced a time-dependent immune dysfunction in association with serum concentrations of the drug as well as cortisol stimulation kinetics. Although total leukocyte count remained unchanged, there was a decrease in the CD4 T/CD8 T-cell ratio due to a decrease in both the percentage and absolute number of CD4 T-helper cells and simultaneous increase in natural killer (NK) cells. Ethanol consumption produced a decrease in T-helper cells and B lymphocytes. The combination of MDMA and ethanol caused the highest suppressive effect on CD4 T cells and increasing effect in NK cells. Drugs treatment produced a high increase of immunosuppressive cytokines (transforming growth factor-β and interleukin-10) and a switch from Th1-type cytokines (interleukin-2 and interferon-γ) to Th2-type cytokines (interleukin-4 and interleukin-10). Disregulation in the production of pro- and anti-inflammatory cytokines with an unbalance toward anti-inflammatory response was also observed. The immune function shows a trend toward baseline levels at 24 h after MDMA kinetics. This transient defect in immunological homeostasis, if temporarily repeated, might alter the immune response with a risk for the general health status.
3,4-Methylenedioxymethamphetamine (MDMA, “ecstasy”) is a widely abused psychomotor stimulant with behavioral effects similar to those elicited by amphetamines and hallucinogens (Peroutka et al., 1988; Wright and Pearl, 1995). MDMA is consumed either alone or in combination with other drugs, as ethanol and cannabis (Gamella and Roldan, 1997).
MDMA use has been associated with neurochemical, behavioral, and endocrine alterations similar to those produced by exposure to acute stress (Mas et al., 1999). It has been shown that stress can produce immune dysfunction and alteration of the distribution of immune cells (Breznitz et al., 1998). In vitro exposure to MDMA resulted in a modulation of several immune functional parameters such as T-cell regulatory function, cytotoxic T-lymphocyte activity, natural killer (NK) cell activity, and macrophage function (House et al., 1995). Administration of MDMA in rats caused a rapid and sustained suppression of induced lymphocyte proliferation and a significant decrease in circulating lymphocytes, which were accompanied by a significantly rapid increase in plasma corticosterone concentrations. It was postulated that the result of altered induced proliferation response of lymphocytes could have been due to a combined effect of direct action of MDMA on lymphocytes and to the activation of the hypothalamic-pituitary-adrenal (HPA) axis and/or the sympathetic nervous system via central mechanisms (Connor et al., 1998). In addition, a dose-related differential alteration in T-helper cell function has been reported (Connor et al., 1999). Acute MDMA administration was also found to enhance NK cell activity in mice (Pacifici et al., 1999a).
The immunomodulating properties of MDMA in humans were recently investigated in two studies. In the first study (Pacifici et al., 2000), acute MDMA treatment was shown to produce a time-dependent immune dysfunction associated with MDMA plasma concentrations. Although total leukocyte count remained unchanged, there was a decrease in CD4 T-helper cells and in CD4 T-helper/CD8 T-suppressor cell ratio, whereas the percentage of NK cells significantly increased. A rise in cortisol plasma concentrations similar to that observed in the rat model supported the hypothesis of MDMA-induced release of corticotrophin-releasing factor from the median eminence of the hypothalamus and subsequent HPA axis and sympathetic nervous system activation. In the second study (Pacifici et al., 1999b), cell-mediated immune response was investigated after the administration of MDMA alone and in combination with ethanol in a limited number of subjects. The decrease in CD4 T-cell counts and simultaneous increase in NK cell activity were confirmed and a decrease in the functional responsiveness of lymphocytes to mitogenic stimulation was also observed after MDMA administration. Ethanol consumption produced a decrease of T-helper cells and B lymphocytes and in the phytohemagglutinin (PHA)-induced lymphocyte proliferation. Combined MDMA and ethanol produced the greatest suppressive effect on CD4 T-cell count and PHA-stimulated lymphoproliferation.
Proliferation and differentiation of lymphocytes are generally controlled by cytokines. There are two different subsets of helper T cells defined by their profile of cytokine production and resulting activation of different immune responses (Mosmann and Coffman, 1989). Th1-type cells predominantly produce interleukin-2 (IL-2), interferon-γ (IFNγ), and tumor necrosis factor-β (TNFβ) and are generally associated with inducing cell-mediated immune responses. Conversely, Th2-type cells produce IL-4, IL-5, IL-6, and IL-10, which are generally associated with humoral immune response. In addition, IL-2 and IFNγ share with IL-1β and TNFα proinflammatory properties and are known to play a role in the pathogenesis of infections and autoimmune diseases (Schreurs, 1993). On the other hand, IL-4 and IL-10 together with transforming growth factor-β1 (TGFβ1) are pleiotropic anti-inflammatory cytokines known to produce profound effects on cells involved in the immune response (Whiteside, 1994). Thus, cytokines appear to be required for almost all immunoregulatory activities, representing potential targets for drug-related immunomodulation (Arai et al., 1990). For this reason, measurement of the expression or secretion of various cytokines was proposed as a strategy for early detection of immunotoxicity (Pallardy et al., 1998).
Studies in rats have shown that administration with MDMA increases the secretion of Th1-type cytokine IL-2 without altering the related Th1-type cytokine IFNγ and inhibited lipopolisaccharide-induced TNFα secretion from stimulated blood cultures, suggesting an impairment of macrophage activity (Connor et al., 2000). These data prompted Connor et al. (2000) to consider their impact on susceptibility to bacterial/viral infections.
The present investigation was designed to examine in healthy volunteers acute immunological changes after administration of MDMA alone and in combination with ethanol. Immune parameters were correlated with blood ethanol and plasma MDMA and cortisol concentrations.
Materials and Methods
Subjects.
Male subjects were recruited by “word of mouth”. Eligibility criteria required the recreational use of MDMA on at least five occasions. Exclusion criteria included consumption of more than 20 cigarettes per day and more than 50 g of ethanol/day (6 units/day). Subjects who fulfilled the inclusion criteria were then interviewed by a psychiatrist (structured clinical interview for DSM-IV) to exclude individuals with a history of major psychiatric disorders, including schizophrenia, psychosis, and major affective disorder. Each participant underwent a general physical examination, routine laboratory tests, urinalysis, and a 12-lead electrocardiogram.
A total of six volunteers was included in the study. The participants had a mean age of 23 years (range 19–36), mean weight of 67.0 kg (range 59–76), and mean height of 175.4 cm (range 169–187). All the subjects declared to be MDMA consumers (times of consumption: range 5–100, mean 24). All subjects but one were current smokers with a range of 0 to 20 cigarettes/day. Their average consumption of ethanol was 1.7 units/day (1 unit = 8 g of ethanol), and all of them were habitual users of cannabis with a previous experience with cocaine and methamphetamine consumption. None had history of abuse or drug dependence according to DSM-IV criteria (except for nicotine dependence), nor any medical or psychiatric adverse reaction after MDMA consumption. Subjects were phenotyped for CYP2D6 activity using dextromethorphan as a drug probe. The destromethorphan/dextrorphan ratio was used to classify subjects as poor or extensive metabolizers (Schmid et al., 1985). All participants were extensive metabolizers.
All volunteers gave their written informed consent before inclusion in the study and were economically compensated for inconveniences caused by their participation in the study. The study was conducted in accordance with the Declaration of Helsinki, approved by the Ethical Committee of our institution (CEIC-IMAS), and authorized by the “Dirección General de Farmacia y Productos Sanitarios” (number 98/112) of the Spanish Ministry of Health.
Study Design.
Subjects participated as outpatients in four 10-h experimental sessions, which were separated by a 1-week washout period. The study design was double blind, double dummy, randomized, crossover, and controlled. Treatment conditions were randomly assigned using a balanced 4 × 4 latin-square design. In the four study sessions, subjects arrived at the laboratory at 8:00 AM after an overnight fast and had an indwelling intravenous catheter inserted into a subcutaneous vein in the forearm of the nondominant arm. Thereafter, they remained seated in a quiet room throughout the session. MDMA was administered around 9:30 AM and ethanol around 10:00 AM. Volunteers were allowed to ingest the ethanolic beverage in a time period of 15 min. At 3 and 6 h after drug administration subjects had a light meal. Tobacco smoking was permitted 6 h after drug administration. The room temperature during the trial remained constant between 20 and 21°C. Volunteers were requested to abstain from consumption of any drug of abuse during the study period, and urine drug testing was performed before each experimental session for opioids, cocaine, cannabis, and amphetamines. For all four groups of substances tested, all volunteers were negative before each experimental session. Some variables not included in the present report such as physiological changes as well as subjective effects and psychomotor performance were also measured.
Drugs.
The four drug conditions were as follows: 100 mg ofd,l-MDMA and placebo of ethanol; 0.8 mg/kg ethanol and placebo of MDMA; 100 mg of d,l-MDMA and 0.8 mg/kg ethanol; and placebos of MDMA and ethanol. The dose of MDMA was selected according to a pilot study (Pacifici et al., 1999b)
MDMA was supplied by the Spanish Ministry of Health. MDMA and MDMA-placebo were prepared by the Pharmacy Service of Hospital del Mar (Barcelona, Spain) as identically appearing opaque, white, soft-gelatin capsules, and administered in fasting state with 100 ml of tap water (two capsules each time). Ethanolic beverage was prepared combining vodka (Absolut, Åhus, Sweden) and tonic water (Schweppes, Madrid, Spain) with a total ethanol dose of 0.8 mg/kg. Several drops of aromatic bitters and lemon juice were also added to mask ethanol placebo made with tonic water. Final volume was 350 ml.
Determination of MDMA and Ethanol Plasma Concentration.
Blood samples for determination of MDMA and ethanol concentrations were drawn before treatment and at 1, 2, 4, 6, 8, 10, and 24 h after drug administration. MDMA was determined in plasma by gas chromatography equipped with a nitrogen-phosphorus detector (Ortuño et al., 1999). Blood ethanol concentrations were determined by a headspace injection technique and gas chromatography equipped with a flame ionization detector (Farré et al., 1997).
Determination of Plasma Cortisol.
Plasma cortisol concentrations were determined by fluorescence polarization immunoassay (Abbott Laboratories, Chicago, IL) according to the manufacturer's instructions. The intra-assay coefficients of variation were 2.9 and 2.6 for low (4.0 μg/dl) and high (40.0 μg/dl) controls, respectively. The assay sensitivity is reported to be 0.45 μg/dl.
Blood Cell Preparation for Immunological Tests.
Blood samples for immunological tests were drawn before treatment and at 1, 1.5, 2, 6, and 24 h after drug administration. Peripheral blood was collected in evacuated tubes containing ethylenediaminetetraacetic acid (0.47 M). Complete blood profile and count were obtained for each participant. Peripheral blood mononuclear cells were obtained by centrifuging whole blood on a Ficoll-Hipaque density gradient. Peripheral blood mononuclear cells were rinsed and suspended in tissue culture (RPMI-1640) containing penicillin (100 U/ml), streptomycin (100 mg/l), and 10% fetal bovine serum.
Peripheral Blood Mononuclear Cell Stimulation.
Peripheral blood mononuclear cells (1 × 107 cells/ml) were cultured on 96-well tissue culture plates and stimulated with 2 μg/ml PHA for interleukin induction. After 72-h incubation at 37°C, plates were centrifuged at 800g for 10 min, and supernatants collected and stored at −80°C (Pacifici et al., 1995). All samples of the same subject were assayed in the same analytical batch.
Cytokine Assay in Culture Supernatant.
For quantitative measurement of IL-1β, IL-4, IL-6, IL-10, TNFα, and IFNγ, six specific solid phase sandwich enzyme-linked immunosorbent assay performed on monoclonal antibody-coated microtiter plate were used (Biosource; Celbio, Milan, Italy). For quantitative measurement of IL-2 and TGFβ1 two specific solid phase enzyme-amplified sensitivity immunoassays performed on monoclonal antibody-coated microtiter plate were used (Biosource). Tests were performed according to the manufacturer's instructions. Samples and IL standards were assayed simultaneously and in duplicate. The standard curves (assay sensitivity) were as follows: IL-1β, 0.31 to 20 (0.19) pg/ml; IL-2, 0.9 to 30 (0.1) U/ml; IL-4, 0.39 to 12.5 (0.27) pg/ml; IL-6, 0.16 to 5.0 (0.10) pg/ml; IL-10, 0.78 to 25 (0.21) pg/ml; TNFα, 0.5 to 16 (0.1) pg/ml; IFNγ, 15.6 to 500 (4) pg/ml; and TGFβ1, 16 to 2000 (2) pg/ml. Cell culture samples were appropriately diluted with the cell culture medium to enter calibration curve ranges. Assay performance was tested using two concentrations of cytokines in culture medium throughout the procedure. Mean intra-assay and interassay coefficients of variation were <6%.
Lymphocyte Immunophenotyping.
A total of 100 μl of whole blood was stained using 20 μl of monoclonal antibody reagent. Becton Dickinson (Becton Dickinson Italia S.p.A., Milan, Italy) fluorescence-activated cell-sorting analysis lysing solution was used to lyse red cells after which stained cells were washed once with PBS and fixed with 1% paraformaldehyde. Stained and fixed lymphocytes were analyzed using an Ortho Cytoron Absolute 4 flow cytometer (Ortho Instruments, Ortho-Clinical Diagnostic, Milan, Italy). The LeucoGATE (CD45/CD14) fluorescent information, with forward and side scatter, was used to set an electronic gate around the lymphoid population. This gate included at least 95% lymphocytes and less than 5% nonlymphocytes (granulocytes, monocytes, and debris). Dual-color immunophenotyping was performed using the following Becton Dickinson matched murine monoclonal antibody reagents directly conjugated to phycoerythrin (PE) or fluorescein isothiocyanate (FITC): CD14/PE-CD45/FITC (leucogate reagent for electronic gating), CD8/FITC-HLA-DR/PE (cytotoxic/suppressor cells), CD4/FITC-HLA-DR/PE (helper/inducer cells), CD3/PE-CD16-CD56/FITC (T mature lymphocytes and natural killer cells), and CD3/PE-CD19/FITC (T mature lymphocytes and B lymphocytes). The absolute number of lymphocytes was calculated by multiplying the percentage of each lymphocyte subset in the flow cytometer by the absolute lymphocyte count.
Statistical Analysis.
Values from lymphocyte subsets, plasma cortisol concentrations, and cytokines were transformed to differences from baseline. The peak effect in the first 6 h (maximum absolute change from baseline values) and the 6-h area under the curve (AUC) of effects versus time calculated by the trapezoidal rule were determined for each variable. These transformations were analyzed by a one-way repeated measures ANOVA with treatment conditions as factor. When ANOVA results showed significant differences between treatment conditions, post hoc multiple comparisons were performed using Tukey's test. Additional comparison of time course of the effects was conducted using repeated measures two-way ANOVA with treatment condition (four) and time (0 to 6 h) as factors. When treatment condition or the treatment condition × time interaction was statistically significant, multiple Tukey's post hoc comparisons were performed at each point of time using the mean square error term of the treatment condition × time interaction. Differences associated withp values lower than 0.05 were considered to be statistically significant.
Results
Data on immunological parameters (lymphocyte subsets and cytokine production) before the first drug treatment are shown in Table1. A two-way ANOVA (immunological parameter versus experimental day) analysis did not show differences in basal values along study time in all parameters tested for the six volunteers. Statistical analysis of results for treatment conditions and changes over the time in blood lymphocyte subsets, plasma cortisol concentrations, and cytokine production by stimulated peripheral blood mononuclear cells induced by the study drugs are presented in Tables2 and 3 and in Figs. 1 and2. Both acute MDMA and combined MDMA and ethanol treatments produced an immune dysfunction. Immune dysfunction was time-dependent and paralleled MDMA plasma drug concentrations, MDMA induced cortisol stimulation kinetics (Fig.3), and blood ethanol concentrations (mean and S.D. of Cmax: 1280 ± 170 mg/l for ethanol administration and 1051 ± 59 mg/l for MDMA-ethanol coadministration; mean and S.D. ofTmax: 2.0 ± 0.5 h for ethanol administration and 1.4 ± 0.3 h for MDMA-ethanol coadministration).
Basal lymphocyte subpopulations and cytokine production in healthy volunteers (n = 6)
Statistical results of lymphocyte subsets and blood cortisol level (area under the curve and peak effects from 0 to 6 h,n = 6)
Statistical results of cytokines production (area under the curve and peak effects from 0 to 6 h, n = 6)

Time course (0–24 h) of drug effects on lymphocyte subsets (differences from baseline). Data points represent means from six subjects. ○, 100 mg of MDMA plus 0.8 mg/kg alcohol; ▵, 100 mg of MDMA; ▿, 0.8 mg/kg alcohol; and ■, placebo. At each time filled symbols indicate a significant difference from placebo (p < 0.05). Differences between active conditions are detailed in Table 2.
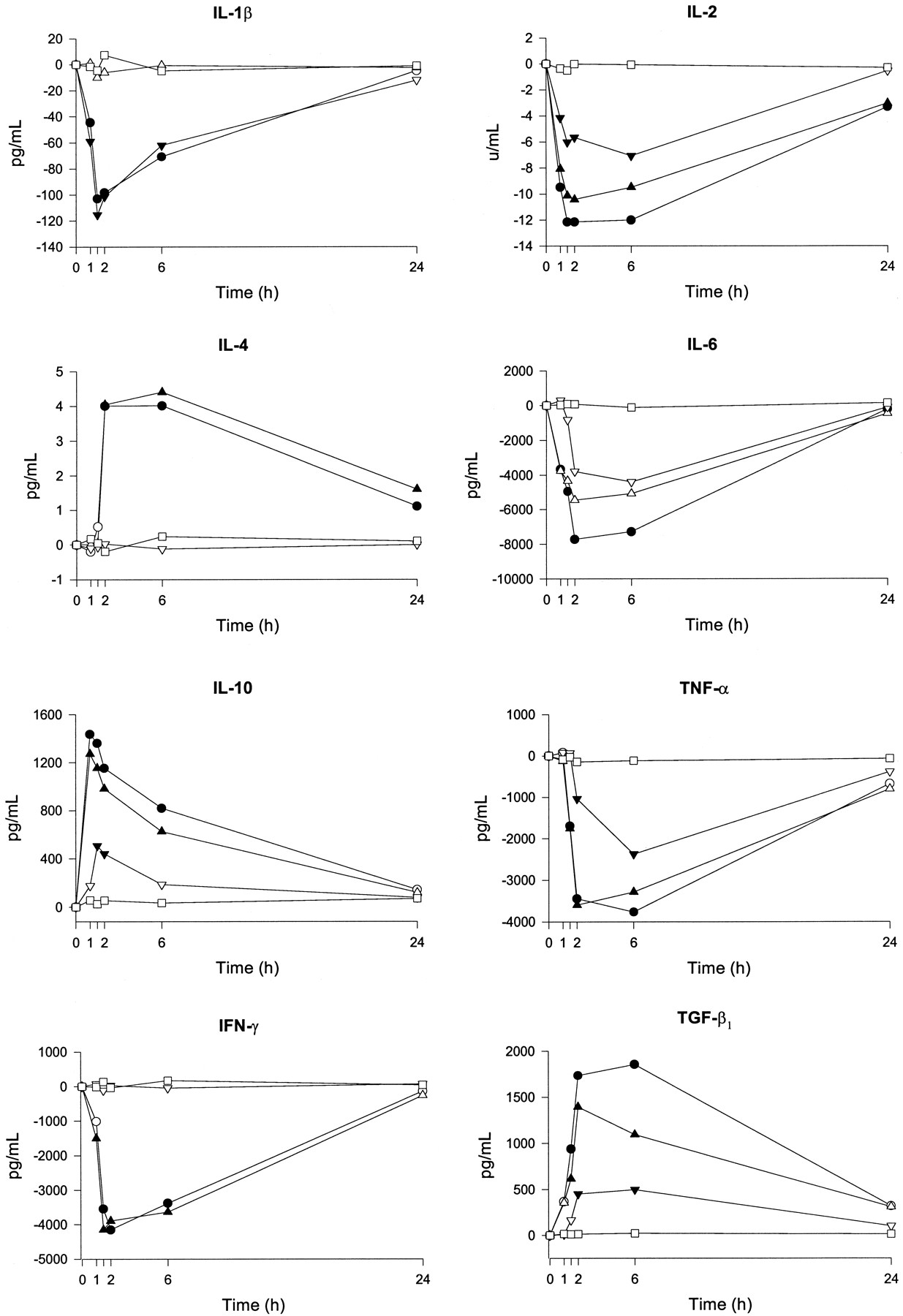
Time course (0–24 h) of drug effects on cytokine production (differences from baseline). Data points represent means from six subjects. ○, 100 mg of MDMA plus 0.8 mg/kg alcohol; ▵, 100 mg of MDMA; ▿, 0.8 mg/kg alcohol; and ■, placebo. At each time filled symbols indicate a significant difference from placebo (p < 0.05). Differences between active conditions are detailed in Table 3.

Plasma concentration-time curve (0–6 h) for MDMA, and time course (0–6 h) of drug effects on plasma cortisol concentrations (differences from baseline) (n = 6). ○, 100 mg of MDMA plus 0.8 mg/kg alcohol; ▵, 100 mg of MDMA; ▿, 0.8 mg/kg alcohol; and ■, placebo. At each time filled symbols indicate a significant difference from placebo (p< 0.05). Differences between active conditions are detailed in Table2.
In case of MDMA administration, all alterations regarding lymphocyte subsets tested peaked between 1 and 2 h from start of the treatment. Particularly, although total leukocyte count remained unchanged, there was a decrease in the CD4 T/CD8 T-cell ratio with a mean peak difference of −0.596 between MDMA and placebo because of decreased proportion of circulating T-helper cells (CD4). No differences were found in the amount of cytotoxic/suppressor lymphocytes (CD8) and of B lymphocytes (CD19). In contrast, there was a high increase of NK cells with a mean peak difference of 311 cells/μl.
Ethanol consumption produced a significant decrease in the proportion of circulating T-helper cells and B lymphocytes (CD19) without altering values of NK cells. Combined MDMA and ethanol produced the greatest suppressive effect on CD4 T-cell count and CD4 T/CD8 T-cell ratio. In fact, the mean peak difference between the combination and the placebo groups was −497 cells/μl for CD4 T-cell count and −0.74 for CD4 T/CD8 T-cell ratio. Conversely, the effect of coadministration of MDMA and ethanol on the reduction of CD19 was less potent than that of ethanol alone. This observation may be related to calculated AUC 0 to 6 h (expressed in mg/l × h−1) of ethanol for both treatment conditions (3769 ± 213 for the combination versus 4082 ± 292 for the ethanol group). As shown in Fig. 3, cortisol plasma concentrations were unchanged in placebo and ethanol treatment conditions, whereas MDMA and combination treatments produced a statistical rise in cortisol concentrations at 2 h after drug administration. Peak drug effects of MDMA versus placebo presented a mean difference of 19.1 μg/dl. When ethanol was coadministered with MDMA, mean peak difference was 15.7 μg/dl.
MDMA treatment was associated with a decrease in the production of the Th1-type cytokines IL-2 and IFNγ (peak effect of −10.53 U/ml and −4214.5 pg/ml, respectively) and an increase in the production of the Th2-type cytokines IL-4 and IL-10 (peak effect of 4.5 pg/ml and 1209.5 pg/ml, respectively). Ethanol treatment produced a decrease of IL-2 secretion with a peak difference with placebo of −7.3 U/ml and a peak IL-10 increase of 518.1 pg/ml.
The equilibrium between proinflammatory (IL-1β, TNFα, and IL-6) and anti-inflammatory (IL-10, IL-4, TGFβ1) cytokines was disrupted by coadministration of MDMA and ethanol. A general trend of decrease in proinflammatory cytokine secretion was observed between ethanol consumption and placebo. Peak differences between treatments were −117.1, −2170, and −5338.8 pg/ml, respectively. In contrast, the anti-inflammatory cytokine TGFβ1 presented a peak increase difference induced by ethanol versus placebo of 477.3 pg/ml. MDMA treatment produced a decrease in TNFα and IL-6 production by stimulated peripheral blood mononuclear cells (peak difference with placebo −3445.2 and −5753.7 pg/ml, respectively) and a peak TGFβ1 increase of 1362.1 pg/ml. When MDMA was combined with ethanol there was an additive phenomenon regarding cytokine production by stimulated peripheral blood mononuclear cells. Immune function was partially restored at 24 h for all treatment conditions.
Discussion
The results show that MDMA administered at doses compatible with its recreational use cause pronounced changes in certain immunological parameters and that these changes occur very rapidly. Indeed, 1 h after the administration, there was a significant reduction of CD4 T-cell count and an increase of NK cell count. These changes in immune function are linked to increased cortisol concentration, which is in turn a consequence of the effects of MDMA administration on central monoaminergic system with the release of corticotrophin-releasing factor and subsequent activation of the HPA axis (Grob et al., 1996). These findings are consistent with results of a pilot study where two different doses of MDMA and 0.8 g/kg ethanol were tested (Pacifici et al., 1999b) and with those of in vitro testing of several 4-substituted amphetamines (Gagnon et al., 1994). Cocaine has been associated with immune alterations in NK cells and helper T cells similar to those observed for MDMA and with activation of the HPA axis, which led to elevation of cortisol levels in humans (Van Dyke et al., 1986; Ruiz et al., 1994; Pellegrino and Bayer, 1998).
Cytofluorimetric analysis revealed that reductions of lymphocyte subset counts were mainly due to T cells and not to B cells. Because evidence suggested that CD4 T-cell function might be suppressed after MDMA treatment, it was investigated whether peripheral blood mononuclear cells exhibited altered Th1-type Th2-type cytokine production, because it is known that Th1/Th2 ratio exerts important effects on the balance of cellular and humoral immunity (Romagniani, 1991). The present data show that MDMA treatment was associated with a decrease in the production of Th1-type cytokines IL-2 and IFNγ and an increase in the production of the Th2-type cytokines IL-4 and IL-10. These results suggested that MDMA might be inducing an unbalanced differentiation of the lymphocytes in peripheral blood as suggested from decrease of CD4 T lymphocytes proportion observed, explaining in part the immunodysfunction caused by MDMA consumption. These findings are consistent with those of Baldwin et al. (1998) and Stanulis et al. (1997) who found disruption of Th1/Th2 balance with a shift toward Th2-type cytokines after the use of cocaine.
Results from the pilot study (Pacifici et al., 1999b) showed that MDMA also impaired lymphoproliferative response to mitogenic stimulation. IL-2, one of key cytokines involved in lymphocyte proliferation, behaves as a hormone-like growth factor that stimulates proliferation of activated T lymphocytes. The decrease of IL-2 secretion, directly correlated with decreasing numbers of CD4 cells in blood of treated subjects, supports IL-2 contribution to the decrease of the lymphoproliferative response. Our results are not in agreement with findings of Connor et al. (2000) in rats, who found that individual Th1-type cytokines IL-2 and IFNγ were differentially affected by the drugs with MDMA increasing IL-2 production and not IFNγ secretion.
Both IFNγ and IL-2 are cytokines produced by Th1 cells and it is coherent that production pattern of the two Th1-type-associated cytokines showed similar kinetic patterns in our study. Decreased release of IL-2 and IFNγ by activated T cells may be another mechanism through which MDMA exerts its immunomodulatory effect. In addition, the equilibrium between proinflammatory (such as IL-1β, TNFα, and IL-6) and anti-inflammatory (IL-10, IL-4, TGFβ1) cytokines, known to play an important role in the inflammatory and immune response (Arai et al., 1990), was altered by MDMA administration. Stimulated peripheral blood mononuclear cells of MDMA-treated subjects released a significantly higher amount of anti-inflammatory cytokines (IL-10, TGFβ1, IL-4) and a significantly lower amount of proinflammatory cytokines (IL-6, TNFα) compared with baseline. In particular, IL-10 concentration, one of the important anti-inflammatory and immunoregulatory cytokines (Mosmann, 1994), was more than 2-fold just after 1 h from MDMA administration. An increase of TGFβ1 quantitatively similar to that of IL-10 but with a more retarded kinetic profile (peak concentration at 2 h) has been also observed, in agreement with what was previously reported for cocaine (Mao et al., 1997). TGFβ1, a pleiotropic immunosuppressive and anti-inflammatory cytokine, has been reported to be involved in negative immunoregulatory functions, such as inhibition of the proliferative response and cytokine production by T-helper cells (Sosroseno and Herminajeng, 1995). TGFβ1 is also known to shift the Th1/Th2 balance toward Th2-type cytokine. Therefore, it may be postulated that the disproportional increase in IL-10 and TGFβ1levels could compromise the T-cell functions by the direct inhibitory effect on T-cell proliferation and by promoting the expansion of T-helper 2-type immune response versus T-helper 1 cell activation.
Acute ethanol treatment produced a significant decrease in the absolute number of T-helper cells and B lymphocytes, and a decrease of proinflammatory cytokines TNFα and IL-1β with a simultaneous increase of inhibitory cytokines TGFβ1 and IL-10. These results are consisted with those of Sacanella et al. (1998) andMandrekar et al. (1996). Ethanol-induced elevation of the IL-10 and TGFβ1 levels contributed to the decreased TNFα production. Down-regulation of inflammatory cytokines TNFα and IL-1β in blood monocytes and alveolar macrophages by acute ethanol treatment has been reported (Verma et al., 1993) and decreased production of TNFα and other inflammatory cytokines by alveolar macrophages in chronically ethanol-fed animals has been implicated as one of the pivotal mechanisms contributing to the increased susceptibility to bacterial pneumonians (Nelson et al., 1989).
Abnormalities of immune function were more pronounced when combining MDMA and ethanol. In combination treatment, more than an interaction between both drugs in synergistic terms, it seems that an additive phenomenon between activities of both drugs on immune function happened. The only exception was represented by the reduction of B lymphocytes, which was more pronounced when ethanol was administered alone. Despite the current belief that nonalcoholic beverages are consumed with MDMA, present findings are clinically relevant becauseGamella and Roldan (1997) have reported ethanol consumption in association with MDMA in up to 76% of the cases.
Moreover, if we consider the use of MDMA, alone and in combination with ethanol as a stressing condition for the central nervous system, rapid and intense effects on the immune response observed in this clinical trial are not surprising. In major stress and in major depression a shift has been observed of the Th1/Th2 balance toward a Th2-type and a Th1-type immune response, respectively. On the other hand, under stressed conditions, the interaction between cells of the immune and nervous system are augmented. It has been recently proposed that there might be a balance between microglia and astrocytes in regulating local immune reaction, including Th1/Th2 responses and secretion of inflammatory mediators (Xiao and Link, 1999).
Acute administration of MDMA alone and in combination with ethanol alters several parameters of immune function in humans. MDMA impaired CD4 T-cell function, responsible for cellular immunity, whereas ethanol consumption produced a decrease in B lymphocytes, known to be responsible for humoral immunity. Hence, in case of MDMA and ethanol coadministration, both cell-mediated and humoral immunity are affected. Changes are transient and immune status shows a trend toward baseline levels at 24 h. Nonetheless, recovery of function at pre-exposure levels could require energetic consumption whose cost on health is difficult to evaluate. Indeed, if we examine basal values of lymphocyte subsets in volunteers of this study, which are recreational healthy consumers of designer drugs, they are within the ranges obtained in a population of blood Spanish donors having same age interval (Larrea et al., 1998). However, it can be noted that volunteers of the study presented values of CD4 (1293.4 ± 221.9 cells/μl) skewed in the upper side of the range of healthy donors (1004.6 ± 443.8 cells/μl) and values of NK (127.3 ± 105.1 cells/μl) with a trend toward low values of blood donors (246.6 ± 180.3 cells/μl). Such observations cannot be linked causally to MDMA consumption because volunteers of the present experiment are concurrent misusers of other substances. There are several reports on the impact of substances such as nicotine, cannabinoids, or cocaine on the immune system, suggesting that in acute conditions they are able to induce profound alterations, whereas in chronic consumers results are more contradictory (Pellegrino and Bayer, 1998). Nevertheless, even a transient defect in immunological homeostasis such as inflammatory cell recruitment and activation may alter immune function with clinical consequences not immediately measurable. Recently, it has been suggested that outcome of different conditions of unknown cause, such as multiple sclerosis, chronic fatigue syndrome, rheumatoid arthritis, or chronic hepatitis, can be related to immune dysregulation (Koziel et al., 1992; Lamanca et al., 1999; Ohshima et al., 1999). In addition, results of this study were obtained in the laboratory setting as opposed to usual MDMA consumption pattern, i.e., in association with other drugs (ethanol, cannabis, cocaine, and tobacco), crowded conditions, stressed physical activity, without control of food intake. This fact could lead to more pronounced immunological changes with enhanced susceptibility to infectious diseases and immunocorrelated pathologies. It has been suggested that cocaine, which displays a common pattern of immune function alteration with MDMA, may be linked of a higher risk of infectious diseases, including AIDS (Pellegrino and Bayer, 1998). The follow-up of a cohort of MDMA consumers and matched controls would be the most powerful approach to assess long-term effects of MDMA on immune system.
Acknowledgments
We thank Esther Menoyo, Isabel Sanchez, Antonella Bacosi, and Nieves Pizarro for assistance in the experimental sessions and laboratory tests, and Dr. Marta Pulido for editing the manuscript.
Footnotes
-
Send reprint requests to: Dr. Rafael de la Torre, Pharmacology Unit, Institut Municipal d'InvestigacióMèdica (IMIM), Doctor Aiguader 80, E-08003 Barcelona, Spain. E-mail: rtorre{at}imim.es
-
This investigation was supported by the Department of Social Affair (Italy) and Fondo de Investigaciones Sanitarias 97/1198, Fondo de Investigaciones Sanitarias 98/0181, Comisio Interdepartamental de Recerca I Innovacio Tecnologica-SGR 99-242, and Plan Nacional Sobre Drogas (Spain).
- Abbreviations:
- MDMA
- 3,4-methylenedioxymethamphetamine
- NK
- natural killer cell
- HPA
- hypothalamic-pituitary-adrenal axis
- PHA
- phytohemagglutinin
- Th1
- T-1 helper lymphocyte (refers to a CD4-subtype cell)
- IL
- interleukin
- IFNγ
- interferon-γ
- TNF
- tumor necrosis factor
- Th2
- T-2 helper lymphocyte (refers to a CD4-subtype cell)
- TGFβ1
- transforming growth factor-β1
- PE
- phycoerythrin
- FITC
- fluorescein isothiocyanate
- AUC
- area under the curve
- Received June 23, 2000.
- Accepted September 19, 2000.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}