


Abstract
Pharmacological responses to aryloxypropanolamines were examined in cells expressing rat or human β1-adrenergic receptors (ARs) using adenylyl cyclase assays. The aryloxypropanolamines CGP 12177 and LY 362884, originally developed as β3-AR agonists, were found to stimulate the β1-AR. Interestingly, both CGP 12177 and LY 362884 exhibited an anomalous biphasic effect on β1-AR. Low concentrations of either CGP 12177 or LY 362884 potently blocked isoproterenol-induced stimulation of β1-AR, whereas higher concentrations of these compounds stimulated the β1-AR. The unusual interaction of these aryloxypropanolamine ligands with the β1-AR was further characterized using β-AR antagonists. Activation of β1-AR by CGP 12177 or LY 362884 was observed to be significantly more resistant to blockade by β-AR antagonists compared with activation by catecholamines. These results suggest that catecholamines and aryloxypropanolamines interact with distinct active conformations of the β1-AR: a state that is responsive to catecholamines and is blocked with high affinity by CGP 12177 and LY 362884, and a novel state that is activated by aryloxypropanolamines but is resistant to blockade by standard β-AR antagonists. Moreover, dependence of antagonist affinity on agonist structure is unprecedented, and its implications on the use of β-AR agonists such as CGP 12177 in receptor classification are discussed.
β-Adrenergic receptors (ARs) are G protein-coupled receptors that are activated by the endogenous catecholamine neurotransmitters epinephrine and norepinephrine. Mutation analysis and computer modeling studies suggest that β-AR activation is primarily determined by the interaction of β-AR ligands with transmembrane amino acid residues (Blin et al., 1993; Granneman et al., 1998). Additional complexities in ligand-receptor interactions also play a role in defining the activity and selectivity of compounds at β-AR. For example, the β2-AR agonist salmeterol interacts with an “exo” site on the β2-AR in addition to its interaction with amino acid residues within the transmembrane region (Green et al., 1996). Furthermore, receptors are capable of existing in an “active” or “inactive” conformation or state, and the ability of a ligand to stabilize a particular conformation of the receptor results in its classification as an agonist, antagonist, or inverse agonist (Samama et al., 1994; Kenakin et al., 1995). Recent studies also suggest that receptors can exist in multiple active conformations (Tuc̆ek, 1997; McLatchie et al., 1998; Muff et al., 1999; Seifert et al., 1999; Zuscik et al., 1998). Thus, differences in the ability of ligands to interact with and stabilize these receptor conformations/states would predict a gradation in the activities of different classes of compounds at β-AR (Kenakin et al., 1995; Ganguli et al., 1998).
The aryloxypropanolamine class of adrenergic ligands displays diverse activities at β-AR, where they function as agonists, inverse agonists, or antagonist. The aryloxypropanolamine CGP 12177 was initially described as a high-affinity antagonist of β1- and β2-ARs (Staehelin and Simons, 1982; Staehelin et al., 1983). It was later demonstrated to be a potent yet partial agonist of the rodent and human β3-AR (Mohell and Dicker, 1989; Granneman et al., 1991, 1993; Granneman and Whitty, 1991). The relatively high potency of CGP 12177 has prompted the development of other β3-AR-selective aryloxypropanolamines with higher efficacies, such as LY 362884 (Arch and Wilson, 1996; Jesudason, 1998). Interestingly, CGP 12177 also increases inotropic and chronotropic responses in the heart, as well as lipolysis in the adipose tissue of mice lacking β3-AR (Kaumann et al., 1998; Preitner et al., 1998). The responses to CGP 12177 in these tissues are resistant to blockade by β-AR antagonists, which is in contrast to the agonist responses induced by catecholamines such as isoproterenol. Based on this unusual pharmacological profile of CGP 12177, the presence of a fourth β-AR subtype was recently proposed (Galitzky et al., 1997; Kaumann and Molenaar, 1997; Malinowska and Schlicker, 1997; Molenaar et al., 1997).
The identity of the receptor mediating the β3-AR-independent effects of CGP 12177 described earlier was clarified in a recent study on brown fat from β-AR knockout mice (Konkar et al., 2000). This study demonstrated that activation of adenylyl cyclase by CGP 12177 in β3-AR knockout mice is mediated by the β1-AR. Significantly, the β1-AR-induced responses to CGP 12177 were resistant to β-AR blockade, implying that they result from a novel interaction of CGP 12177 with β1-AR. In this study, we have further characterized the pharmacological properties of the β1-AR response to CGP 12177 and examined whether other aryloxypropanolamines that have been developed as β3-AR agonists exhibit similar behavior. We confirm here that CGP 12177 stimulates β1-AR and that activation of β1-AR by CGP 12177 is resistant to blockade by β-AR antagonists, in contrast to activation by catecholamines. We also show that a second aryloxypropanolamine, LY 362884, has pharmacological properties similar to CGP 12177. Our results suggest that differences in aryloxypropanolamine versus catecholamine sensitivity to β-blockers are a result of their interactions with distinct active conformations or states of the β1-AR.
Experimental Procedures
Materials.
(−)-Isoproterenol bitartrate, (−)-propranolol hydrochloride, and dobutamine hydrochloride were obtained from Sigma Chemical Co. (St. Louis, MO). (−)-Norepinephrine bitartrate, CGP 12177, and CGP 20712A were obtained from Research Biochemicals International (Natick, MA). LY 362884 was obtained from Eli Lilly (Indianapolis, IN). SB 251023 and SB 226552 were obtained from SmithKline Beecham Pharmaceuticals (King of Prussia, PA). The chemical structures of aryloxypropanolamine ligands used in this study are illustrated in Fig. 1. Materials for adenylyl cyclase assays were obtained from sources previously described (Chaudhry and Granneman, 1991). Ham's F-12 medium was purchased from Irvine Scientific (Santa Anna, CA). Minimum essential medium, penicillin/streptomycin, and geneticin were obtained from Life Technologies (Gaithersburg, MD). All other chemicals were of reagent grade.

Chemical structures of aryloxypropanolamine ligands used in this study.
Cell Culture.
Chinese hamster ovary (CHO) and HEK 293 cells were grown in Ham's F-12 and minimum essential medium, respectively, supplemented with 10% fetal calf serum, l-glutamine (2 mM), penicillin (100,000 units/l), and streptomycin (100 mg/l). CHO and HEK 293 cells were grown at 37°C in a humidified atmosphere of 5% and 10% CO2, respectively. Cells were harvested at approximately 90% confluence. The rat and human β1-ARs were cloned and expressed in either CHO or HEK 293 cells as described previously (Chaudhry et al., 1992; Konkar et al., 2000). The rat β1-AR was expressed at 43 fmol/mg of protein and 3.6 pmol/mg of protein in CHO and HEK 293 cells, respectively. The human β1-AR was expressed in CHO cells at 40 fmol/mg of protein. The human β1-AR cDNA was a gift from Dr. S. Liggett (University of Cincinnati, Cincinnati, OH).
Preparation of Membranes.
Cells were washed twice with PBS (pH 7.4) and then lysed in a hypotonic homogenization buffer containing 25 mM HEPES (pH 8.0), 2 mM MgCl2, 1 mM EDTA, and 10 μg/ml leupeptin. Cell membranes were collected with a rubber policeman and pelleted by centrifugation at 48,000g for 15 min at 4°C. Membranes were stored at −80°C and were resuspended in homogenization buffer immediately before use in experiments.
Adenylyl Cyclase Assay.
Adenylyl cyclase activity was determined according to a modification (Granneman et al., 1991) of the method of Salomon (1979). Briefly, membranes (5–20 μg of protein) were incubated with agonists at 4°C in the presence or in the absence of various β-AR antagonists for 20 min. The adenylyl cyclase reaction was initiated by the addition of the substrate mix and continued for 20 min at 30°C. The contents of the substrate mix and chromatographic separation of cAMP have been described previously (Chaudhry and Granneman, 1994). Any deviations from the methods described here are provided in legends to figures.
Data for antagonism by β-AR antagonists were analyzed using a Schild plot (Arunlakshana and Schild, 1959), and the pA2 value was calculated for each separate experiment together with the gradient of the Schild regression. Prior incubation of membranes with β-AR antagonists for 1 h at 4°C (data not shown) did not alter pA2 values from those reported under Results. Protein was measured according to the Bradford assay using bovine serum albumin as a standard. Kinetic parameters (Kact or EC50 and Vmax) of adenylyl cyclase activity were estimated by nonlinear regression analysis (Prism; GraphPad Software, San Diego, CA).KB values were calculated as follows from the estimates of EC50 values of concentration-response curves obtained in the absence and the presence of a single concentration of antagonist; pKB = −logKB = [antagonist]/(CR − 1); CR = EC50′/EC50, where EC50 is the agonist concentration required to elicit a half-maximal response in the absence of an antagonist and EC50′ is the agonist concentration required to elicit a half-maximal response in the presence of an antagonist. The EC50′ values for catecholamines and aryloxypropanolamines were determined in the presence of 0.1 and 1.0 μM concentration of the β-blocker, respectively.
Statistical Analysis.
Data are presented as mean ± S.E. Student's t test was used to evaluate differences between means, and critical values of P < .05 were judged statistically significant.
Results
Agonist Responses to Aryloxypropanolamines at β1-AR.
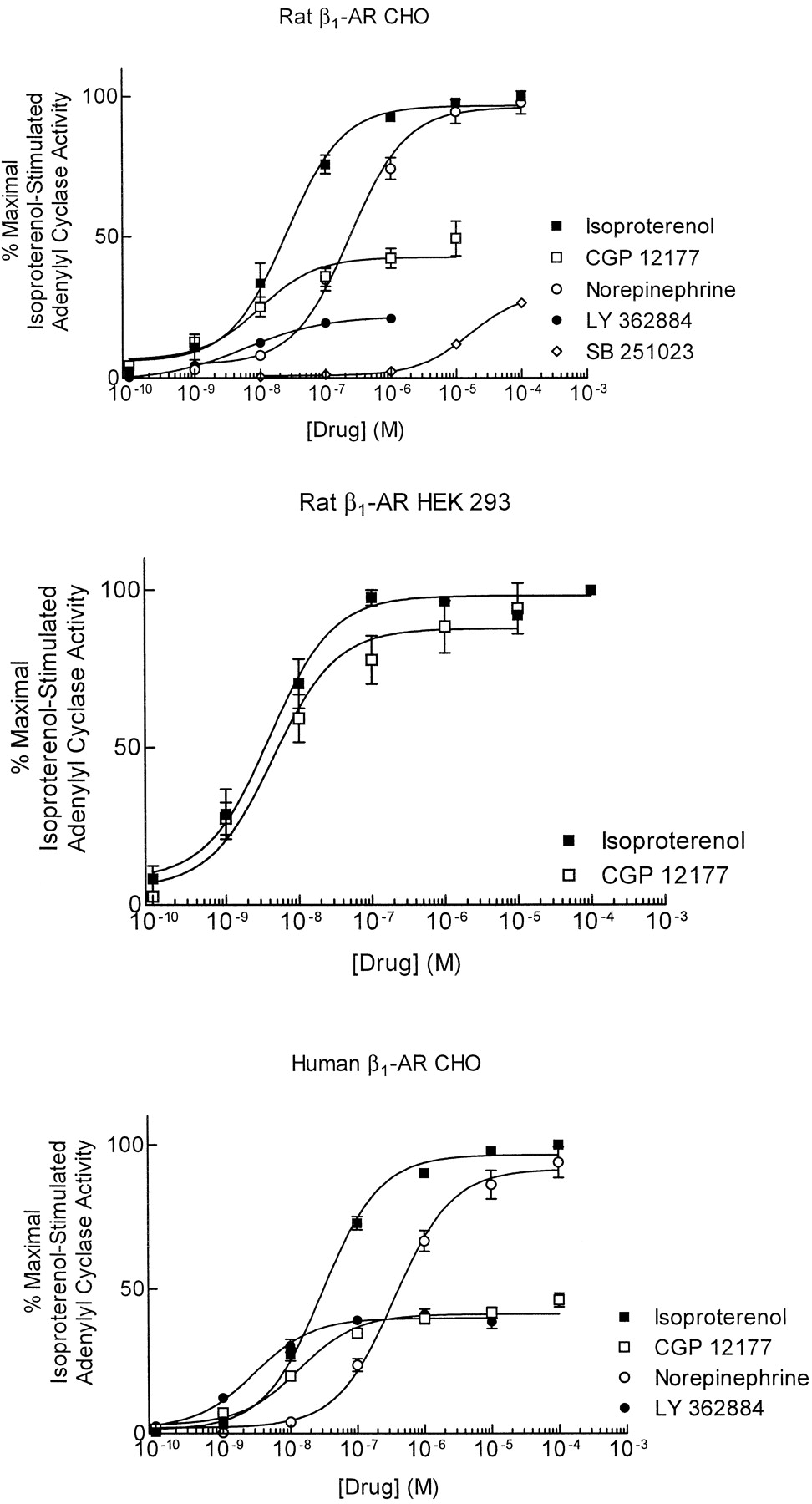
The aryloxypropanolamine CGP 12177 (see Fig.1 for structure) is an agonist of β1-AR (Fig.2), as previously reported (Pak and Fishman, 1996; Konkar et al., 2000). CGP 12177 exhibited potent but partial agonist activity in CHO cells expressing low levels (approximately 40 fmol/mg of protein) of rat or human β1-AR and had agonist potencies (EC50 values) of 14.8 and 12.5 nM, respectively. In HEK 293 cells overexpressing (3.6 pmol/mg of protein) rat β1-AR, CGP 12177 stimulated the rat β1-AR with a potency of 3.3 nM (Fig. 2; Table1). CGP 12177, however, did not stimulate adenylyl cyclase activity in untransfected cells (data not shown). Thus, activation of β1-AR by CGP 12177 was independent of cell type and was observed for both rat and human β1-AR. The agonist response to CGP 12177 suggested that other aryloxypropanolamine ligands developed as β3-AR agonists might also display agonist activity at β1-AR. Consistent with this hypothesis, we observed that the aryloxypropanolamine ligands LY 362884 and SB 251023 (Jesudason et al., 1998; Arch et al., 1999) elicited partial agonist responses at β1-AR expressed in CHO cells. SB 226552 (Sennitt et al., 1998) stimulated β1-AR, but only at the highest concentrations used (100 μM) (Table 1).

Comparison of CGP 12177-, isoproterenol- and norepinephrine-induced activation of rat (top and middle) and human (bottom) β1-ARs expressed in CHO and HEK 293 cells. Adenylyl cyclase activation is used as a measure of β1-AR activation. Data are presented as a percentage of the maximal response to isoproterenol. Values are means ± S.E. of 4 to 16 experiments
Pharmacological parameters of rat and human β1-AR activation by catecholamine and aryloxypropanolamine ligands
Antagonism of Catecholamine-Induced β1-AR Activation by Aryloxypropanolamine Ligands.
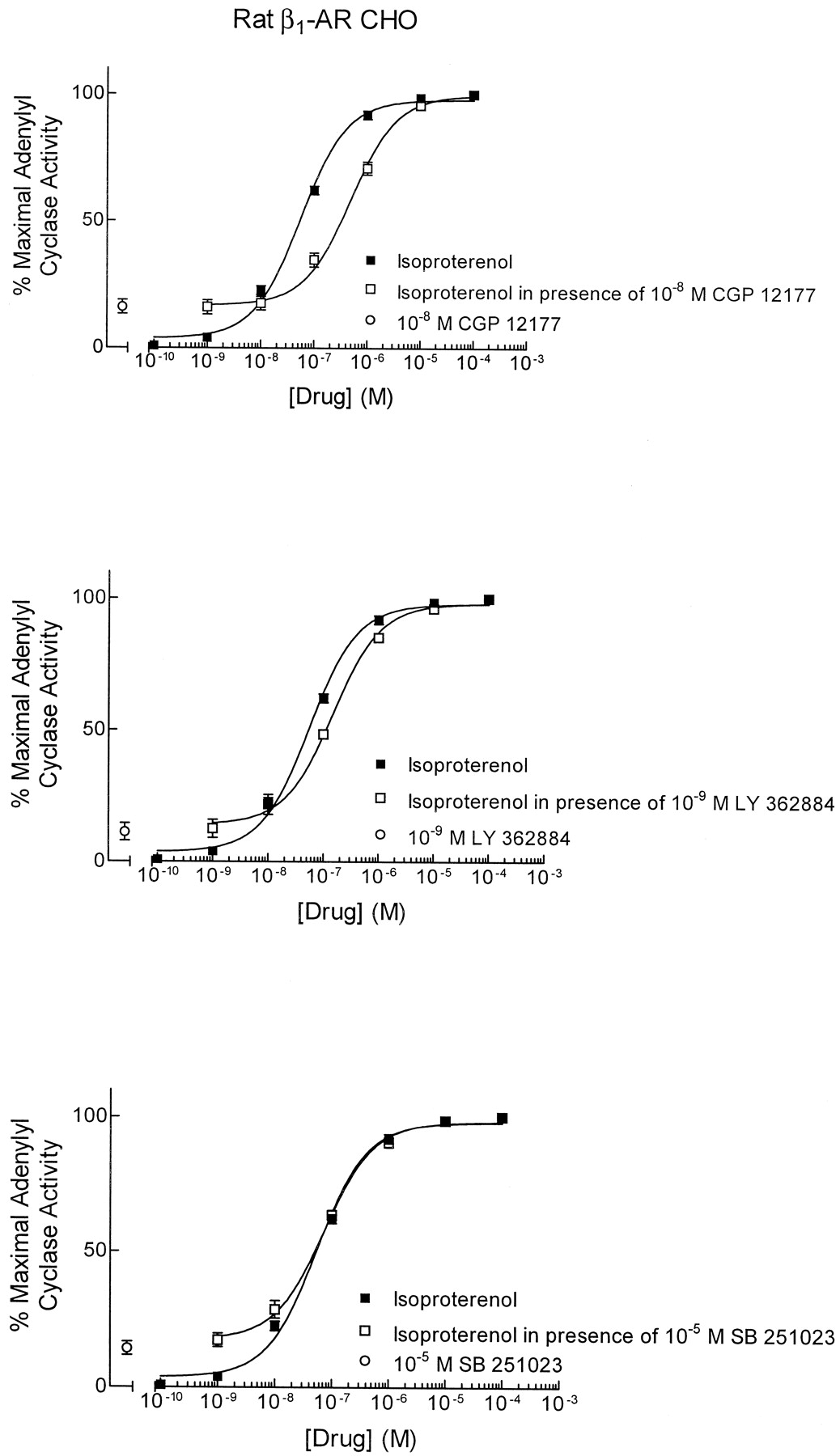
CGP 12177 was initially described as a high-affinity ligand of the β1- and β2-AR that did not stimulate these receptors in tissues except at high concentrations (Staehelin et al., 1983). Our results showed that CGP 12177 stimulated β1-AR with relatively high potency. We therefore examined whether CGP 12177 and LY 362884 behaved as high-affinity antagonists by examining their effects on isoproterenol-induced stimulation of β1-AR. CGP 12177 (10 nM) and LY 362884 (1 nM) shifted the isoproterenol concentration-response curve rightward by 6- and 2-fold, respectively (Fig. 3). The calculated affinity estimates (KBvalues) were 2 and 0.8 nM for CGP 12177 and LY 362884, respectively. Thus, both CGP 12177 and LY 362884 were potent blockers of β1-AR. The aryloxypropanolamine SB 251023 (10 μM), however, failed to block isoproterenol-induced stimulation of β1-AR (Fig. 3).

Antagonism of isoproterenol-induced stimulation of rat β1-adrenergic receptors expressed in CHO cells by 10 nM CGP 12177 (top), 1 nM LY 362884 (middle), and 10 μM SB 251023 (bottom). Adenylyl cyclase activation is used as a measure of β1-AR stimulation. Values are mean ± S.E. of three or four experiments.
Antagonist and Agonist Response to Aryloxypropanolamine Compounds at β1-AR.
The agonist responses to CGP 12177 and LY 362884 at β1-AR occurred at concentrations seven and four times greater, respectively, than those required to block the β1-AR (Figs. 2 and 3), implying that the antagonist and agonist activities of these compounds represent distinct interactions with the β1-AR that can be distinguished by concentration. We therefore constructed concentration-response curves to CGP 12177 and LY 362884 in the presence and in the absence of isoproterenol. A concentration of isoproterenol (10 nM) that is equieffective to the maximal response of either aryloxypropanolamine was used in these experiments. The catecholamine partial agonist dobutamine was used as a reference control.
As shown in Fig. 4, dobutamine exhibited a monotonic inhibition of isoproterenol-stimulated adenylyl cyclase activity, as expected of a simple partial agonist. In contrast, both CGP 12177 and LY 362884 elicited a biphasic response in the presence of an equieffective concentration of isoproterenol. Low concentrations of CGP 12177 or LY 362884 effectively inhibited the response to isoproterenol, whereas higher concentrations of the aryloxypropanolamines stimulated adenylyl cyclase activity (Fig. 4). The agonist response observed in the presence of isoproterenol was identical to the agonist response observed in absence of isoproterenol. These results suggest that CGP 12177 and LY 362884 display both agonist and antagonist activities at the β1-AR that can be distinguished by concentration. We also examined the effects of SB 251023 on the isoproterenol-induced stimulation of β1-AR (Fig. 4). Unlike CGP 12177 and LY 362884, SB 251023 failed to inhibit the agonist response of isoproterenol but instead displayed behavior similar to the catecholamine partial agonist dobutamine.

Effect of CGP 12177, LY 362884, SB 251023, and dobutamine on isoproterenol-stimulated adenylyl cyclase activity at rat β1-ARs expressed in CHO cells. CHO cell membranes expressing the rat β1-AR were stimulated for 30 min at 30°C with increasing concentrations of CGP 12177 (A), LY 362884 (B), SB 251023 (C), or dobutamine (D) in the presence or in the absence of isoproterenol. Values presented are mean ± S.E. of three experiments.
Effect of β-AR Antagonists on Response to CGP 12177 and LY 362884.
The observation that low concentrations of CGP 12177 and LY 362884 block β1-AR, whereas high concentrations stimulate β1-AR suggested that these compounds interact with distinct β1-AR sites or states to elicit antagonist and agonist responses. These results also suggest that the catecholamine-responsive site or state is distinct from the aryloxypropanolamine-responsive site or state. Thus, we hypothesized that β-blockers might differentially block these distinct sites or states of the receptor. In the following experiments, we compared the relative potencies of the β-AR antagonists propranolol and CGP 20712A for blocking activation of β1-AR by catecholamines and aryloxypropanolamines.
CGP 12177-induced activation of human β1-AR was significantly more resistant to blockade by CGP 20712A and propranolol than were isoproterenol- and norepinephrine-induced activations (Figs.5 and 6). Schild plot analysis indicated that a 4-fold greater concentration of CGP 20712A (P < .05) was required to block CGP 12177-stimulated versus isoproterenol-stimulated β1-AR activity (Fig. 5). Similarly, 18- and 15-fold higher concentrations of propranolol (P < .001) were required to block CGP 12177-stimulated versus isoproterenol- and norepinephrine-stimulated β1-AR activity (Fig. 6).

Antagonism of isoproterenol- and CGP 12177-induced activation of human β1-ARs by increasing concentrations of CGP 20712A in CHO cells (top and bottom, respectively). Adenylyl cyclase activation is expressed as a percentage of the maximal response to isoproterenol or CGP 12177. Insets, corresponding Schild plots. Values are presented as mean ± S.E. of four to six experiments.

Antagonism of isoproterenol-, norepinephrine-, and CGP 12177-induced activation of human β1-ARs by increasing concentrations of propranolol in CHO cells (top, middle, and bottom, respectively). Adenylyl cyclase activation is expressed as a percentage of the maximal response to isoproterenol, norepinephrine, or CGP 12177. Insets, corresponding Schild plots. Values are presented as mean ± S.E. of three to seven experiments.
Activation of the rat β1-AR by CGP 12177 was also resistant to β-AR blockade. An 8-fold higher concentration of CGP 20712A (P < .05) was required to block CGP 12177-induced responses at rat β1-AR compared with isoproterenol- and norepinephrine-induced responses (Table 1). Similarly, 24- and 26-fold higher concentrations of propranolol (P < .001) were required to block stimulation of β1-AR by CGP 12177 versus stimulation by isoproterenol and norepinephrine, respectively (Table 1). This unusual resistance to β-AR blockade exhibited by CGP 12177 at the β1-AR expressed in CHO cells was verified in HEK 293 cells, where a 9-fold higher concentration of CGP 20712A (P < .05) was required to block β1-AR stimulation by CGP 12177 versus stimulation by isoproterenol. In contrast, the partial β1-AR agonist dobutamine displayed sensitivity to β-blockade similar to the full catecholamine agonists isoproterenol and norepinephrine (Table 1).
The availability of LY 362884 allowed us to examine whether resistance to β-AR blockade was a unique property of CGP 12177. LY 362884-induced stimulation of the rat β1-AR was 12-fold (P < .05) more resistant to blockade by the β1-selective antagonist CGP 20712A compared with stimulation by isoproterenol. Similarly, a 26-fold higher concentration of propranolol (P < .001) was required to block LY 362884-induced β1-AR activation compared with activation by isoproterenol (Table 1). Stimulation of the human β1-AR by LY 362884 was 35-fold (P < .001) more resistant to blockade by propranolol compared with catecholamines (Table 1).
Discussion
CGP 12177 was initially described as a high-affinity antagonist of β1- and β2-ARs that possessed moderate sympathomimetic properties at higher concentrations (Staehelin et al., 1983). The compound was later observed to be a potent but partial agonist of the rodent and human β3-AR (Mohell and Dicker, 1989; Granneman et al., 1991, 1993) and was used as a prototypical drug for developing highly efficacious β3-AR selective agonists (Arch and Wilson, 1996). In recent studies, CGP 12177 was observed to stimulate cardiovascular and metabolic responses that were resistant to β-AR blockers in wild-type and β3-AR knockout mice (Kaumann et al., 1998; Preitner et al., 1998). These findings suggested the presence of a novel β-AR subtype that has been designated β4-AR (Galitzky et al., 1997;Kaumann and Molenaar, 1997). A more recent study demonstrated that CGP 12177-induced responses attributed to activation of the putative β4-AR in native tissues actually occur through its interaction with β1-AR (Konkar et al., 2000). We have therefore used pharmacological analyses to examine in detail the interaction of CGP 12177 with β1-AR in a cell expression system. We have also examined whether other aryloxypropanolamines, developed as β3-AR agonists (Jesudason, 1998; Sennitt et al., 1998; Arch et al., 1999), exhibit behavior similar to that of CGP 12177 at β1-AR.
In concurrence with earlier observations, low concentrations of CGP 12177 potently blocked isoproterenol-induced activation of β1-AR (Staehelin et al., 1983), whereas at higher concentrations, CGP 12177 behaved as an agonist. Another aryloxypropanolamine, LY 362884, displayed behavior similar to that of CGP 12177. Independent estimates showed that inhibitory potencies (KB values) of CGP 12177 and LY 362884 were 7- and 4-fold higher than their stimulatory potencies (EC50 values), respectively. Thus, in the presence of an equieffective concentration of isoproterenol, both CGP 12177 and LY 362884 exhibited a biphasic response at the β1-AR. At low concentrations, CGP 12177 and LY 362884 blocked β1-AR stimulation by isoproterenol, whereas higher concentrations of these compounds stimulated the β1-AR. The β1-AR stimulation profiles of these compounds in the presence of isoproterenol were identical to the profiles obtained in the absence of isoproterenol. This behavior suggests that although both aryloxypropanolamines block activation of β1-AR by isoproterenol, they also stimulate β1-AR independently of the catecholamine-responsive site or state. The biphasic effect demonstrated by CGP 12177 and LY 362884 is in stark contrast to that observed for the catecholamine dobutamine and the aryloxypropanolamine SB 251023. The latter two compounds exhibit a monotonic inhibition of isoproterenol-stimulated adenylyl cyclase activity, as predicted for simple partial agonists (Ariëns, 1983). Another aryloxypropanolamine (SB 226552) evaluated in this study produced a weak stimulation of the β1-AR and consequently could not be used for detailed pharmacological analyses. However, an earlier study shows that this compound does not interact with the putative β4-AR (Sennitt et al., 1998) and thus would be expected to behave in a manner analogous to dobutamine and SB 251023. These data suggest that the partial agonists dobutamine and SB 251023 interact with the catecholamine-responsive site or state of β1-AR. In contrast, the aryloxypropanolamines CGP 12177 and LY 362884 appear to interact with two different sites or states of the β1-AR.
The presence of an aryloxypropanolamine-responsive β1-AR site or state that is distinct from the catecholamine-responsive state raises the possibility that these two sites or conformations may differ in their interactions with β-AR ligands. We therefore examined the ability of β-AR antagonists to block responses to aryloxypropanolamines and catecholamines to further characterize the aryloxypropanolamine-responsive state of β1-AR. If aryloxypropanolamines and catecholamines interact with the same site or conformation of β1-AR to produce agonist responses, β-AR antagonists should block responses to each compound with equal potency. However, we observe that CGP 12177- and LY 362884-induced responses at β1-AR are consistently and significantly resistant to β-AR blockade compared with the responses to the catecholamines isoproterenol, norepinephrine, and dobutamine. These data provide further evidence indicating that catecholamines and aryloxypropanolamines interact with distinct active conformations of β1-AR.
In this study, we used CGP 12177 in functional assays to define a novel state of β1-AR. It is unclear, however, whether this novel state can be labeled and defined using currently available radioligands. In earlier studies, Pak and Fishman (1996) demonstrated that CGP 12177 binds to a high- and a low-affinity site of β1-AR. The low-affinity CGP 12177 binding site constituted ∼10% of the receptor population and was sensitive to the guanine nucleotide guanosine-5′-O-(3-thio)triphosphate, indicating that it is the agonist-binding site for CGP 12177. In native tissues, however, [3H]CGP 12177 labels a site that is insensitive to propranolol blockade and guanosine-5′-O-(3-thio)triphosphate, and this site was tentatively defined as the putative β4-AR (Kaumann et al., 1998; Sarsero et al., 1998, 1999). Studies using β1- and β3-AR knockout mice show that β1-AR activation completely accounts for the β4-AR-like profile seen in native tissues (Rohrer et al., 1996; Konkar et al., 2000). It is thus likely that the propranolol-resistant [3H]CGP 12177 binding observed in native tissues represents an interaction between CGP 12177 and the novel state of the β1-AR defined here. Our study shows that CGP 12177 activity at β1-AR is resistant to blockade by propranolol. However, propranolol discriminates between the two active states of β1-AR with rather low selectivity. Thus, this antagonist seems unsuitable to accurately define the novel β1-AR state in radioligand binding studies. Further characterization will be possible with the development of highly selective ligands for the aryloxypropanolamine-sensitive state of the β1-AR.
Receptors are thought to exist in an active state or an inactive state, and their ligands are classified as agonists, inverse agonists, or neutral antagonists based on their interactions with these states. Agonists preferentially stabilize the active state, whereas inverse agonists stabilize and promote the inactive state of the receptor. Neutral antagonists, however, lack preference for either the active or the inactive receptor state (Kenakin et al., 1995). These results indicate that CGP 12177 and LY 362884 do not fall into one or the other category but possess both antagonist and agonist properties at β1-AR that are concentration-dependent. Molecular modeling indicates that β-AR ligands can assume either folded or extended conformations, and only the latter conformation allows receptor activation (Blin et al., 1993). The relatively constrained ligand binding pockets of β1- and β2-AR allow interaction with the folded conformations of β-AR ligands, where they behave as antagonists. The less constrained binding pocket of the β3-AR accommodates extended conformations of β-AR ligands and results in receptor activation. Significantly, β3-AR activation by catecholamines and aryloxypropanolamines is poorly blocked by propranolol. Thus, the novel β1-AR state activated by aryloxypropanolamines seems to possess pharmacological properties similar to the β3-AR. The two β1-AR active states described here may allow binding of folded and extended conformations, with the aryloxypropanolamine-responsive state exhibiting β3-AR-like properties of activation by extended conformers and relatively low affinity for propranolol.
Pak and Fishman (1996) found a direct correlation between β1-AR receptor number and CGP 12177 responsiveness in a heterologous expression system. Interestingly, the proportion of the CGP 12177 responsive sites, however, remained constant regardless of the change in expression level of β1-AR. Although these results agree with earlier analysis correlating CGP 12177 responsiveness to receptor expression, overexpression of β1-AR does not appear to be a prerequisite for CGP 12177 response because agonist activity was observed in native tissues (Konkar et al., 2000) and CHO cells expressing rat and human β1-AR at physiological levels. Moreover, expression level does not alter the antagonist potencies because CGP 20712 blocked the rat β1-AR with similar potencies when expressed at physiological levels or at a level approximately 100-fold greater. The molecular basis of the atypical state of the β1-AR is unknown. The CGP 12177-sensitive state might be generated through posttranslational modifications of the receptor or represent homomeric or heteromeric protein interactions. In this regard, we have noted that the activity of CGP 12177 relative to isoproterenol varies among tissues (our unpublished observations), suggesting that the stabilization of the atypical β1-AR state may involve interactions with other proteins (Tang et al., 1999), perhaps in a manner analogous to that described for calcitonin gene-related peptide receptors and receptor activity modifying proteins (McLatchie et al., 1998). Clearly, further work is needed to clarify the molecular basis of aryloxypropanolamine activation of β1-AR and the impact of these ligands on β1-AR signaling.
These results also have implications for the use of antagonist affinity values (pA2 orKB values) in defining and classifying receptors. Antagonists affinity values are used to define receptor subtypes because classic receptor theory maintains that the potency of an antagonist for a given receptor subtype remains constant regardless of the agonist used to elicit the response (Kenakin, 1982; Kenakin et al., 1992). Current receptor theory models also indicate that a given receptor can exist in a conformation that is either active or inactive (Samama et al., 1994). Recent studies suggest that a given receptor can exist in more than one active conformation (Tuc̆ek, 1997;McLatchie et al., 1998; Muff et al., 1999; Seifert et al., 1999; Zuscik et al., 1998). Agonist ligands can thus be predicted to interact with receptor-binding sites differentially based on the conformation presented (Ganguli et al., 1998), and hence antagonist affinities at a given receptor might differ in an agonist-dependent fashion. Consistent with this hypothesis and in contrast to current receptor theory models, we find that antagonist affinities at β1-AR can be agonist-dependent.
In summary, this report presents evidence indicating that the aryloxypropanolamine CGP 12177 is an antagonist of β1-AR at low concentrations and an agonist of β1-AR at high concentrations. In contrast to catecholamines, the agonist response to CGP 12177 is resistant to β-AR blockade. Another aryloxypropanolamine, LY 362884, also displays an unusual pharmacological profile similar to that exhibited by CGP 12177. The blockade of catecholamine-induced stimulation of β1-AR by CGP 12177 and LY 362884 and the resistance of CGP 12177- and LY 362884-induced activation of β1-AR to β-blockers suggest that certain aryloxypropanolamine compounds can interact with at least two distinct active conformations of β1-AR.
Footnotes
-
Send reprint requests to: Dr. James G. Granneman, Parke-Davis Pharmaceutical Research, 2800 Plymouth Rd., Ann Arbor, MI 48201. E-mail: james.granneman{at}wl.com orjgranne{at}med.wayne.edu
-
↵1 This work was supported by United States Public Health Service Grant DK46339.
- Abbreviations:
- AR
- adrenergic receptor
- CHO
- Chinese hamster ovary
- Received February 22, 2000.
- Accepted May 9, 2000.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}