


Abstract
Phosphorothioate oligodeoxynucleotides produce splenomegaly and mononuclear cell infiltrates in multiple organs in mice after repeated i.v. administration. Several phosphorothioate oligodeoxynucleotides were studied to better understand the basis of immunostimulatory properties of these molecules in mice and to study the effects of chemically modified oligonucleotides. Chemical modifications examined included 5-methyl cytosine and 2′-methoxyethoxy substituents. Male mice (six per group) were treated with oligonucleotide concentrations of 0, 2, 10, or 50 mg/kg by i.v. injection every other day for 14 days. Immune stimulation was assessed 24 h after the last dose by measuring spleen weight, or histologic and immunohistochemical examination of liver and kidney. Immune stimulation was dose-dependent for the phosphorothioate oligodeoxynucleotides studied, but potency varied as a function of sequence. Results from this study reveal that there is a close correlation between the extent of splenomegaly and other evidence of immune stimulation, such as the severity of cell infiltrates in liver and kidney in mice. Immunohistochemical analysis indicated that cell infiltrates in liver and kidney were primarily mononuclear cells associated with increased expression of the endothelial-leukocyte cellular adhesion molecule intracellular adhesion molecule-1 and the cytokine interleukin-6. Immune stimulation was markedly decreased with oligonucleotides containing the 5-methyl cytosine and further decreased by 2′-methoxyethoxy modifications. Administration of these modified oligonucleotides to mice did not produce splenomegaly even at the 50-mg/kg dose, and only produced minimal cell infiltrates despite the presence of comparable or greater tissue oligonucleotide concentrations. Thus, chemical modifications appeared to increase the tolerability profile for these compounds that are representative of the second generation of antisense oligonucleotides.
With the progression of antisense oligonucleotides toward use as human therapeutics, there has been extensive characterization of these molecules in animal models of pharmacology, as well as toxicology (Bennett et al., 1996; Bennett, 1998; Levin et al., 1998). The promise of antisense therapeutics is to use the specificity afforded by Watson-Crick hybridization to inhibit the expression of a single disease-causing protein, such as the case of selective inhibition of the protein kinase C-α isozyme by ISIS 3521 (Dean and McKay, 1994;Dean et al., 1994; McKay et al., 1996). Although there is ample evidence of sequence-dependent activity on intended molecular targets, there are also a number of hybridization-independent effects that have been characterized for phosphorothioate oligodeoxynucleotides (Bennett, 1998; Levin et al., 1998). An understanding of the relative potency and mechanism of hybridization-independent effects is important because these effects can complicate the interpretation of pharmacology experiments or may produce undesired toxicity.
Common examples of hybridization-independent effects of unmodified phosphorothioate oligodeoxynucleotides following systemic administration in mice include dose-dependent splenomegaly, lymphoid hyperplasia, and mixed mononuclear cell infiltrate in numerous tissues (Sarmiento et al., 1994; Branda et al., 1996; Henry et al., 1997b,c). These effects appear to be interrelated and are collectively regarded as a form of immune stimulation. Furthermore, these effects have been observed in rodents for most unmodified phosphorothioate oligodeoxynucleotides studied to date (Monteith et al., 1997).
Evidence from in vitro experiments suggests that this immune stimulation is a result of direct cellular activation. Exposure of isolated B lymphocytes or peripheral blood leukocytes to phosphorothioate oligodeoxynucleotides produces a mitogenic response with stimulation of growth, Ig expression, and cytokine release (Pisetsky and Reich, 1994; Krieg et al., 1995; Klinman et al., 1996;Liang et al., 1996; Boggs et al., 1997; Zhao et al., 1997). Other cell types, such as human keratinocytes, also have been shown to be activated and produce cytokine release [interleukin (IL)-1α] when exposed to oligonucleotides (Crooke et al., 1996). Although immune stimulation is a common property of phosphorothioate oligodeoxynucleotides with nonmethylated cytosine, there are sequence motifs that appear to increase the potency of immune stimulation. A palindromic motif was identified in oligonucleotides that potentiated immune stimulation (Yamamoto et al., 1994). Another laboratory reported similar findings in which oligodeoxynucleotides containing a cytosine-guanine (CpG) dinucleotide motif with two purines on the 5′ end and two pyrmidines on the 3′ end provided optimal mitogenic activation of splenocytes (Krieg et al., 1995, 1996). Oligonucleotides with these sequence motifs have more pronounced immunostimulatory effects relative to other phosphorothioate oligodeoxynucleotides.
Phosphorothioate oligodeoxynucleotides designed for antisense applications typically do not contain the optimal sequence motif for immune stimulation, and the potency of this side effect is generally low relative to intended antisense pharmacology (Bennett et al., 1997). Furthermore, the potency of immune stimulation is species specific and monkeys have been shown to be less sensitive to the immunostimulatory effects of phosphorothioate oligodeoxynucleotides compared with mice in vivo (Henry et al., 1997b). However, to minimize the potential for these side effects, it is desirable to identify future generations of antisense oligonucleotides with reduced potential for immune stimulation. Chemical modifications to the typical phosphorothioate oligodeoxynucleotide can modulate the potency of immune stimulation. For example, methylation of cytosine, especially those in CpG motifs, decreases the potency of immune stimulation in vitro (Krieg et al., 1996). Modification of the 2′ position of ribose sugar backbone with a methoxy moiety also has been reported to decrease immune stimulation in vivo (Zhao et al., 1995). In addition, 2′-modified oligonucleotides have equal or greater antisense activity relative to unmodified phosphorothioate oligodeoxynucleotides (Monia et al., 1993, 1996;Altmann et al., 1996; Baker et al., 1997).
In the present study, the multiorgan cellular infiltrate has been characterized as a component of the immune stimulation resulting from systemic administration of phosphorothioate oligodeoxynucleotides in mice. The time course of cell infiltrates has been studied and the dose-response severity correlated to the extent of splenomegaly. Immunohistochemical techniques were used to identify infiltrating cell types, adhesion molecule expression, and local cytokine production. In addition, comparison was made between several phosphorothioate oligodeoxynucleotides and chemically modified compounds to examined the relative effects on immune stimulation in rodents. In particular, oligonucleotides containing 5-methyl cytosine (5-methyl C) nucleotides, as opposed to cytosine, in the presence or absence of 2′-methoxyethyl (2′-MOE) modifications have been investigated. General toxicity parameters for phosphorothioate oligodeoxynucleotide and modified oligonucleotides also were investigated. The results from these analyses demonstrate that both 5-methyl C substitution and 2′-MOE modifications reduce immune stimulation in rodents.
Materials and Methods
Oligonucleotides.
The oligonucleotides used in this study contain various sugar and nucleobase modifications as described in Table 1. Oligonucleotides were synthesized at Isis Pharmaceuticals, Inc., with a Milligen model 8800 DNA synthesizer with solid-phase triester chemistry as described inBeaucage and Iyer (1992). 2′-Deoxy phosphoramidites were purchased from commercial sources (Pharmacia); the 2′-O-MOE phosphoramidites were manufactured specifically for Isis Pharmaceuticals under contract. The phosphorothioate internucleotide linkage was generated by use of Beaucage reagent as the P(III) 224 P(V) oxidant. The test compounds were purified by reversed phase HPLC to a final purity of >92% full-length oligomer (determined by capillary electrophoresis and strong anion exchange HPLC). There were no detectable levels of endotoxin in the final lyophilized product as determined by Limulus Amebocyte Lysate analysis (Associates of Cape Cod, Woods Hole, MA). Lyophilized bulk drug substance was formulated in phosphate-buffered saline (pH 7.4) at the concentrations used in this study (0.4, 2, and 10 mg/ml).
Oligonucleotide sequence and modifications
Animal Studies.
Male CD-1 mice were obtained from Charles River Breeding Laboratories (Wilmington, MA), C3H and C57BL/6 mice were obtained from Harlan Breeders (Indianapolis, IN). Mice were randomly chosen and assigned to dose groups based on body weight. The mice were ∼6 to 8 weeks old (20–30 g b.wt.) at the initiation of the study. The mice were housed individually in metal cages with suspended wire-mesh floors, and were maintained in an environmentally controlled room (12-h light/dark cycle; 72 ± 6°C; 55 ± 15% relative humidity) with ad libitum access to standard rodent feed (Agway rodent diet; Agway, Syracuse, NY) and water. All animal husbandry procedures were performed in full compliance with American Association of the Accreditation of Laboratory Animal Care guidelines.
For the comparison of strain sensitivity to immune stimulation, mice (three per strain per time point) received 1, 3, or 7 doses of 50 mg/kg ISIS 1082 by i.v. injection. Doses were administered on a daily schedule. Twenty-four hours after the last dose, kidney, liver, spleen, and lung were processed for routine histopathologic examination to evaluate the extent of mononuclear cell infiltrate.
In experiments designed to compare different oligonucleotides, mice (six per group) were injected on alternate days for 14 days (seven total doses) with oligonucleotide (2, 10, and 50 mg/kg/injection), or vehicle (phosphate-buffered saline, pH 7.4). Mice were injected via the tail vein with a constant volume of 10 ml/kg, and a rate of ∼2 ml/min. During the treatment period, all animals were observed twice daily for viability and once daily for signs of toxicity. Additional antemortem observations included body weight and food consumption measurements obtained at weekly intervals throughout treatment period.
Necropsies were performed the day after the last injection (i.e., day 15) after the start of the study for the recovery group animals. Before necropsy, all animals were fasted overnight and blood was collected from the retro-orbital sinus for evaluation of hematology and serum chemistry parameters. Water remained available ad libitum. Immediately before blood collection, all animals were lightly anesthetized with ether. The animals were sacrificed by exsanguination while under ether anesthesia. Serum chemistry parameters included aspartate aminotransferase, alanine aminotransferase (ALT), alkaline phosphatase, creatine kinase, total bilirubin, albumin, globulin, total protein, blood urea nitrogen, creatinine, cholesterol, triglycerides, sodium, potassium, chloride, calcium, phosphate, and glucose. Hematologic parameters included both total and differential leukocyte counts, erythrocyte counts, hemoglobin concentration, hematocrit, mean corpuscular volume, mean corpuscular hemoglobin, mean corpuscular hemoglobin concentration, platelet counts, and reticulocyte counts.
At necropsy, a complete examination of all body cavities was conducted. Selected organs (i.e., kidney, liver, spleen, lung, heart, brain, adrenals, and testes/ovaries) were excised, trimmed of fat and connective tissue, and weighed. These organs, as well as a number of other organs and tissues, were preserved in 10% neutral buffered formalin, embedded in paraffin, and stained with H&E for histopathologic evaluation.
Immunohistochemistry Staining.
Four-micrometer-thick cryostat sections were fixed in ice-cold acetone and stained with rat monoclonal antibodies directed against mouse antigens to detect a variety of antigens. The list of antigens examined and source of antibodies used are summarized in Table2. Primary antibodies were detected with horseradish peroxidase-conjugated donkey anti-rat IgG F(ab′)2 (Jackson ImmunoResearch, West Grove, PA). All slides were stained on a Dako Autostainer (Dako Corp., Carpenteria, CA). Diaminobenzidine was used as a substrate and sections were counterstained with hematoxylin, dehydrated, and mounted with permanent mounting medium.
Antibodies used for immunostaining
Results
Kinetics of Mononuclear Cell Infiltrates.
Several mouse strains were examined to study strain differences in sensitivity to immune stimulation by phosphorothioate oligodeoxynucleotides and the time course of cell infiltrates following single or multiple doses. ISIS 1082, a 21-base phosphorothioate oligodeoxynucleotide targeting UL13 gene product of human herpes simplex virus-1, has previously been reported to produce a mononuclear cell infiltrate in rat liver, spleen, kidney, and lung (Henry et al., 1997a). In the present experiment, CD-1, C3H, and C57BL/6 mice were treated with 50 mg/kg ISIS 1082 by i.v. injection for one, three, and seven daily doses. Kidney, liver, lung, and spleen were analyzed 24 h after the last dose by routine histology to evaluate the extent of mononuclear cell infiltrate. A single dose of ISIS 1082 produced a focal mononuclear cell infiltrate in the liver for one out of three CD-1 mice (Table3). The remaining tissues appeared normal after a single administration. No mononuclear cell infiltrate was observed in any tissue examined for C3H or C57BL/6 mice after a single dose of this oligonucleotide. Three daily doses of ISIS 1082 produced a mild mononuclear cell infiltrate in all tissues examined for two out of three C3H mice. None of the other mice strains exhibited a mononuclear cell infiltrate after three daily doses of ISIS 1082 (Table 3). Seven daily doses of ISIS 1082 produced a mononuclear cell infiltrate in all three mouse strains with C3H mice exhibiting the most pronounced effects to ISIS 1082, followed by CD-1 mice, and then C57BL/6 mice. Seven daily doses of 50 mg/kg ISIS 1082 only produced focal infiltrates in liver, lung, and spleen of C57BL/6 mice, whereas the same dose produced moderate infiltrates in the other two strains. Based on results from this analysis, and our previous experience, CD-1 mice were chosen for the remainder of the experiments.
Incidence and severity of multiorgan cell infiltrate in various strains of mice after i.v. administration of ISIS 1082
Immune Stimulation by Phosphorothioate Oligodeoxynucleotides.
To characterize the immunohistochemical changes induced by phosphorothioate oligodeoxynucleotides with different sequences, CD-1 mice were treated for 14 days with 0, 2, 10, or 50 mg/kg i.v. doses of four different phosphorothioate oligodeoxynucleotides. Spleen weights were used to assess the level of immune stimulation produced by the oligonucleotides. Three of the oligonucleotides (ISIS 1082, ISIS 5132, and ISIS 12959) have sequences designed to inhibit specific mRNA (Table1), while the fourth (ISIS 12449) was designed to have optimal B cell-stimulating properties based on previous published work (Yamamoto et al., 1994; Krieg et al., 1995). All phosphorothioate oligodeoxynucleotides studied produce increased spleen weight at the 50-mg/kg dose, however, there was considerable variability in the extent of splenomegaly for different sequences (Fig.1). The oligonucleotide that produced the greatest increase in spleen weight (ISIS 12449) contained an optimal sequence (Krieg et al., 1995) for mitogenic stimulation of B cells (AACGTT). ISIS 12449 produced almost a 2-fold increase in spleen weight even at a dose of 2 mg/kg and greater than a 3.5-fold increase in spleen weight at 10 mg/kg (Fig. 1A). Extensive evidence of immune stimulation occurred in mice treated with 50 mg/kg ISIS 12449, however, spleen weights were not reported due to mortality and morbidity at this dose. By comparison, ISIS 5132 and ISIS 1082 produce more moderate effects on spleen weights, with no effect at 2 mg/kg, 1.5- to 2-fold increase at doses of 10 mg/kg, and 2- to 3.5-fold increase at 50 mg/kg (Fig. 1A). Although these oligonucleotides also contain CpG dinucleotide motifs, they did not possess optimal flanking sequences for immune stimulation. Interestingly, the oligonucleotide that produced the least amount of immune stimulation (ISIS 12959) was essentially the same sequence as ISIS 5132 with the exception of two nucleotide substitutions that changed a CpG motif to cytosine-adenosine (Table 1), confirming a role of this motif in determining the potency of immune stimulation.
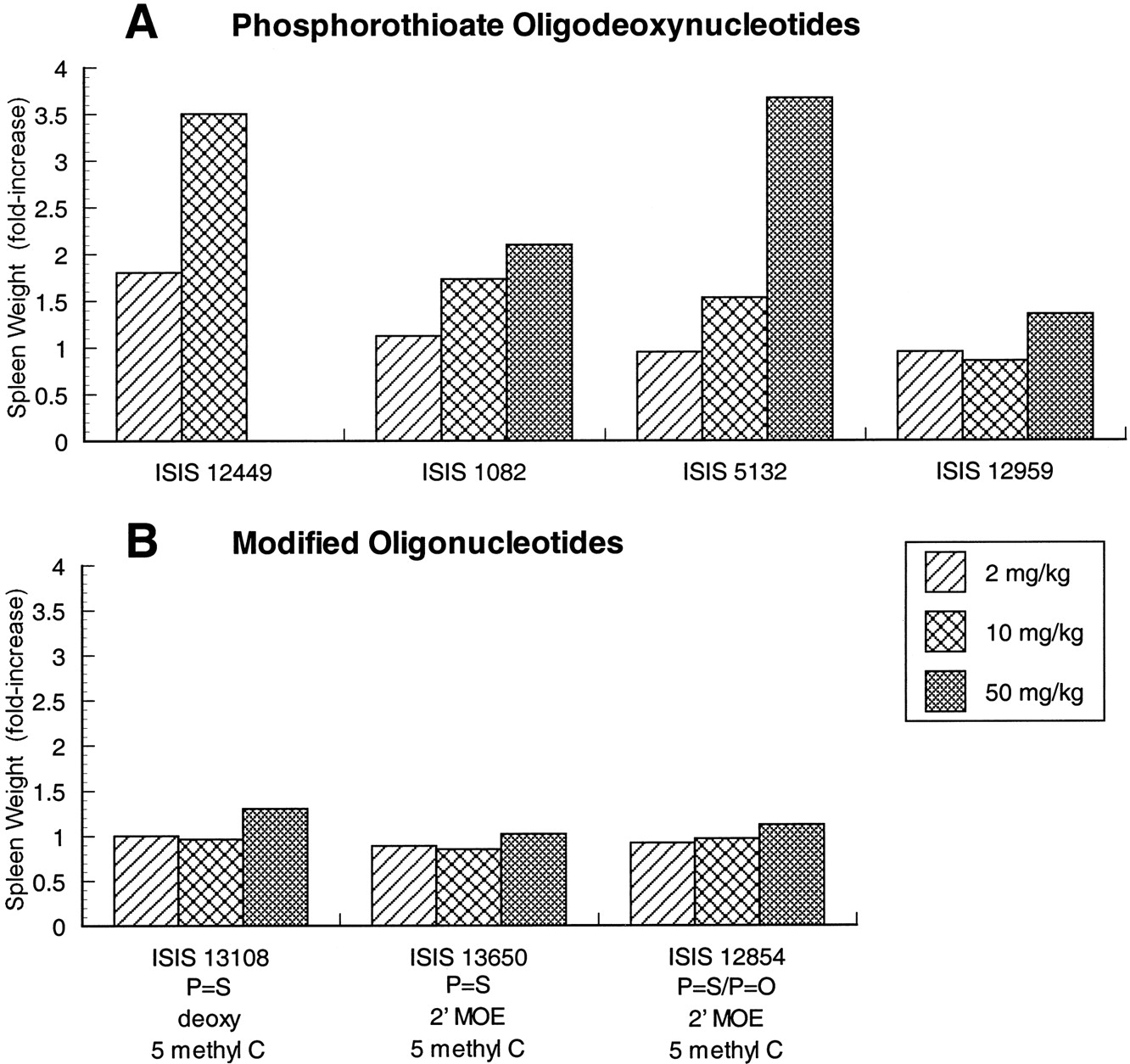
Effect of oligonucleotide treatment on spleen weights in male mice. The effects of several phosphorothioate oligodeoxynucleotides (A) are compared with modified oligonucleotides (B) containing 5-methyl C substitutions or 2′-MOE substituents. Oligonucleotides were administered by every-other-day administration for 14 days. Organ weights were determined 24 h after the last dose. Values represent the relative change in mean organ weight from control group of five or six mice in each case. Standard deviation was <25% of the mean in all cases.
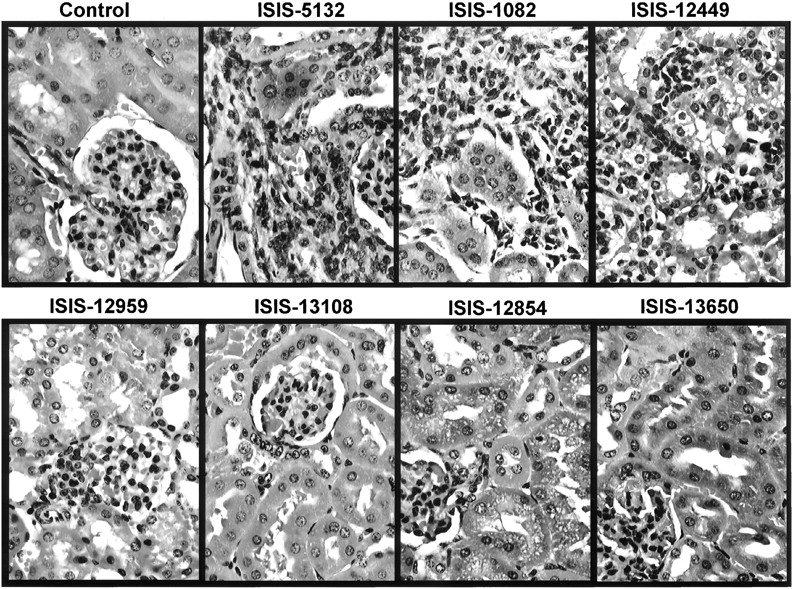
Microscopic evidence of immune stimulation included lymphoid hyperplasia (primarily follicular hyperplasia) and mononuclear cell infiltrates in multiple organs. Qualitatively these changes appeared to be proportional to the extent of splenomegaly observed for the various phosphorothioate oligodeoxynucleotides. Lymphoid hyperplasia in mice treated with 50 mg/kg ISIS 1082 or ISIS 5132 was observed in spleen and lymph nodes, and primarily characterized by hyperplasia of putative B cells in the follicles (T-cell-dependent B cells) and marginal zone (T-cell-independent B cells). Monocytic cellular infiltrates in the liver were generally perivascular (Fig.2), and in the cortical interstitium of the kidney (Fig. 3). Several other tissues, including lung and muscle, also had mononuclear cell infiltrates at 50 mg/kg (data not shown).

Mixed mononuclear cell infiltrates in liver of male mice treated with various oligonucleotides. Oligonucleotides were administered at 50 mg/kg by i.v. injection every other day for 14 days. Tissues were collected and processed for H&E 24 h after the last dose. Original magnification, 500×.

Mixed mononuclear cell infiltrates in kidney of male mice treated with various oligonucleotides. Oligonucleotides were administered at 50 mg/kg by i.v. injection every other day for 14 days. Tissues were collected and processed for H&E 24 h after the last dose. Original magnification, 500×.
To provide a better understanding of the inflammatory changes occurring within tissues following treatment with high doses of phosphorothioate oligodeoxynucleotides, liver and kidney sections were stained with antibodies recognizing a variety of leukocyte antigens and molecules up-regulated in response to inflammatory stimuli (Table4). Leukocyte function-associated antigen (LFA)-1 (CD11a) and CD18 are α- and β subunits of leukointegrins broadly expressed on most leukocytes. Cells infiltrating kidney and liver were positive for both LFA-1 and CD18, consistent with a mononuclear cell infiltrate (Table 4). To further define which leukocyte type was infiltrating into tissue, samples were stained for antigens with a more restricted expression pattern. Mac-1, also a leukointegrin, is expressed on monocytes, granulocytes, NK (natural killer) cells, and subsets of B and T lymphocytes, whereas Ly6G (Gr-1) is an antigen expressed predominantly on granulocytes. The typical pattern of leukocyte antigens and cytokines associated with cell infiltrates is represented in Fig.4, comparing standard tissue histology to immunohistochemical analysis of liver from a mouse treated with vehicle control or 50 mg/kg ISIS 1082. Many of the cells infiltrating tissue were positive for Mac-1 expression and negative for Ly6G (Table 4; Fig.4), suggesting that many of the infiltrating cells were monocytes. Identification of mononuclear cells confirms interpretation of morphological analysis (Figs. 2 and 3). Cells infiltrating tissue were also VLA-4 positive, although there were fewer VLA-4 positive cells than Mac-1 positive (Table 4). Only a minor proportion of the infiltrating cells were CD4 positive T lymphocytes.
Immunohistological characterization of adhesion molecules and chemokines responsible for mononuclear cell accumulation in tissues after phosphorothioate oligonucleotide treatment in mice

Correlation of adhesion molecular expression and cytokine expression with mixed mononuclear cell infiltrate in mice treated with 50-mg/kg ISIS 1082. Mice were treated with either vehicle control or 50-mg/kg ISIS 1082 every other day for 14 days. Tissues were processed 24 h after the last dose and stained with H&E, or with specific immunohistochemical staining for Mac-1, ICAM-1, or IL-6. The immunohistochemical staining used antibodies specific to the specified protein, immunoperoxidase, and were counterstained with hematoxylin. Original magnification, 500×.
Consistent with an increase in cell infiltrates, there was an increased expression of the adhesion molecules intercellular adhesion molecule-1 (ICAM-1) and vascular cell adhesion molecule-1 on endothelium in animals treated with high doses of ISIS 1082 (Table 4; Fig. 4). In addition, ICAM-1 was expressed on inflammatory cells migrating into the tissue in regions of severe inflammation. Of the cytokines examined, there was increased expression of IL-6 in regions of inflammatory cell infiltrate (Table 4; Fig. 4). Similar association between cell infiltrates and cell surface markers, adhesion molecules, and cytokine expression was observed in kidney (data not shown). Comparison across the phosphorothioate oligodeoxynucleotides studied revealed that the amount of ICAM-1 and Mac-1 staining in liver correlated with the severity of cell infiltrates, with ISIS 12449 producing the most severe effects and ISIS 12959 having the least potent effects (data not shown).
Immune Stimulation by Chemically Modified Oligonucleotides.
A phosphorothioate oligodeoxynucleotide (ISIS 13108) with a sequence identical with ISIS 1082, and modified to contain 5-methyl C, as opposed to cytosine, was tested in mice to confirm the ability of this substitution to reduce the potency of immune stimulation (Krieg et al., 1996). Mice were treated with ISIS 13108 at 2, 10, or 50 mg/kg every other day for 2 weeks. Spleen weights were increased ∼30% over control values at the 50-mg/kg dose compared with a >2-fold increase for ISIS 1082-treated mice (Fig. 1, A and B). The severity of lymphoid hyperplasia and mononuclear cell infiltrates in mice treated with ISIS 13108 also were decreased relative to ISIS 1082 at 50 mg/kg (Figs. 2and 3), which were paralleled by decreases in ICAM-1 and Mac-1 staining in liver and kidney (data not shown). Although the extent of immune stimulation was decreased by the 5-methyl C substitution, mononuclear infiltrates and lymphoid hyperplasia were still observed at the high dose in animals treated with ISIS 13108. These results demonstrate that 5-methyl C substitution not only decreases immune cell activation in vitro but also in vivo.
The potency of immune stimulation also was decreased in oligonucleotides containing both 5-methyl C substitutions and 2′-methoxyethoxy ribose sugar modifications (ISIS 13650 and ISIS 12854) (Table 1). ISIS 13650 and ISIS 12854 have the same nucleotide sequence as ISIS 5132. Mice treated for 14 days with 2, 10, and 50 mg/kg ISIS 13650 (phosphorothioate backbone) or ISIS 12854 (phosphodiester/phosphorothioate mixed backbone) exhibited no increases in spleen weight compared with the phosphorothioate oligodeoxynucleotide with the same sequence, ISIS 5132 (Fig. 1, A and B). There was also very little lymphoid hyperplasia or mononuclear cell infiltrates in liver and kidney in mice treated with 50 mg/kg ISIS 13650 or 50 mg/kg ISIS12854 (Figs. 2 and 3). Furthermore, there was essentially no increase in immunohistochemical staining for ICAM-1, Mac-1, or IL-6 relative to control animals (Table 4).
This observable reduction of immune stimulation by phosphorothioate oligonucleotides containing 5-methyl C and 2′-MOE modifications cannot be explained by a decrease in drug accumulation in various tissues. The tissue distribution profile for 2′-MOE modified oligonucleotides was similar to that described for phosphorothioate oligodeoxynucleotides, in that the target organs for accumulation of these 2′-alkoxy modified oligonucleotides (i.e., liver and kidney) are the same as that shown for phosphorothioate oligodeoxynucleotides (Table5). Furthermore, the increased resistance to nuclease degradation of 2′-MOE modified oligonucleotides (ISIS 13650) resulted in organ concentrations 4- to 5-fold greater than have been reported for ISIS 5132 with comparable doses and treatment schedules (Table 5) (Geary et al., 1997).
Average concentration of ISIS 5132 or ISIS 13650 in tissues after alternate day treatment for 3 weeks5-b
General Toxicology.
In addition to evaluation of immune stimulation, several general toxicity parameters were assessed in animals treated with 2′-MOE modified oligonucleotides relative to phosphorothioate oligodeoxynucleotides. Phosphorothioate oligodeoxynucleotides (i.e., ISIS 12449, ISIS 5132, and ISIS 1082) produced approximately a 1.4-fold increase in liver weight at 50 mg/kg relative to control animals (Fig. 5A). This change in organ weight was correlated with increases in ALT at 50 mg/kg (Fig. 6A) and microscopic changes in the liver that included Kupffer cell hypertrophy and single-cell hepatocyte necrosis (data not shown). These changes have been observed for a number of different phosphorothioate oligodeoxynucleotides, and have been attributed to the high concentrations of oligonucleotide attained in liver with repeated administration of high doses. ISIS 5132 and ISIS 1082 also produced dose-dependent decreases in platelet counts (Fig. 7A). At a dose of 50 mg/kg, platelet counts were ∼44 and 25% of baseline values for ISIS 5132 and ISIS 1082, respectively.

Effect of oligonucleotide treatment on liver weights in male mice. The effects of several phosphorothioate oligodeoxynucleotides (A) are compared with modified oligonucleotides (B) containing 5-methyl C substitutions or 2′-MOE substituents. Oligonucleotides were administered by every-other-day administration for 14 days. Organ weights were determined 24 h after the last dose. Values represent the relative change in mean organ weight from control group of five or six mice in each case. Standard deviation was <25% of the mean in all cases.
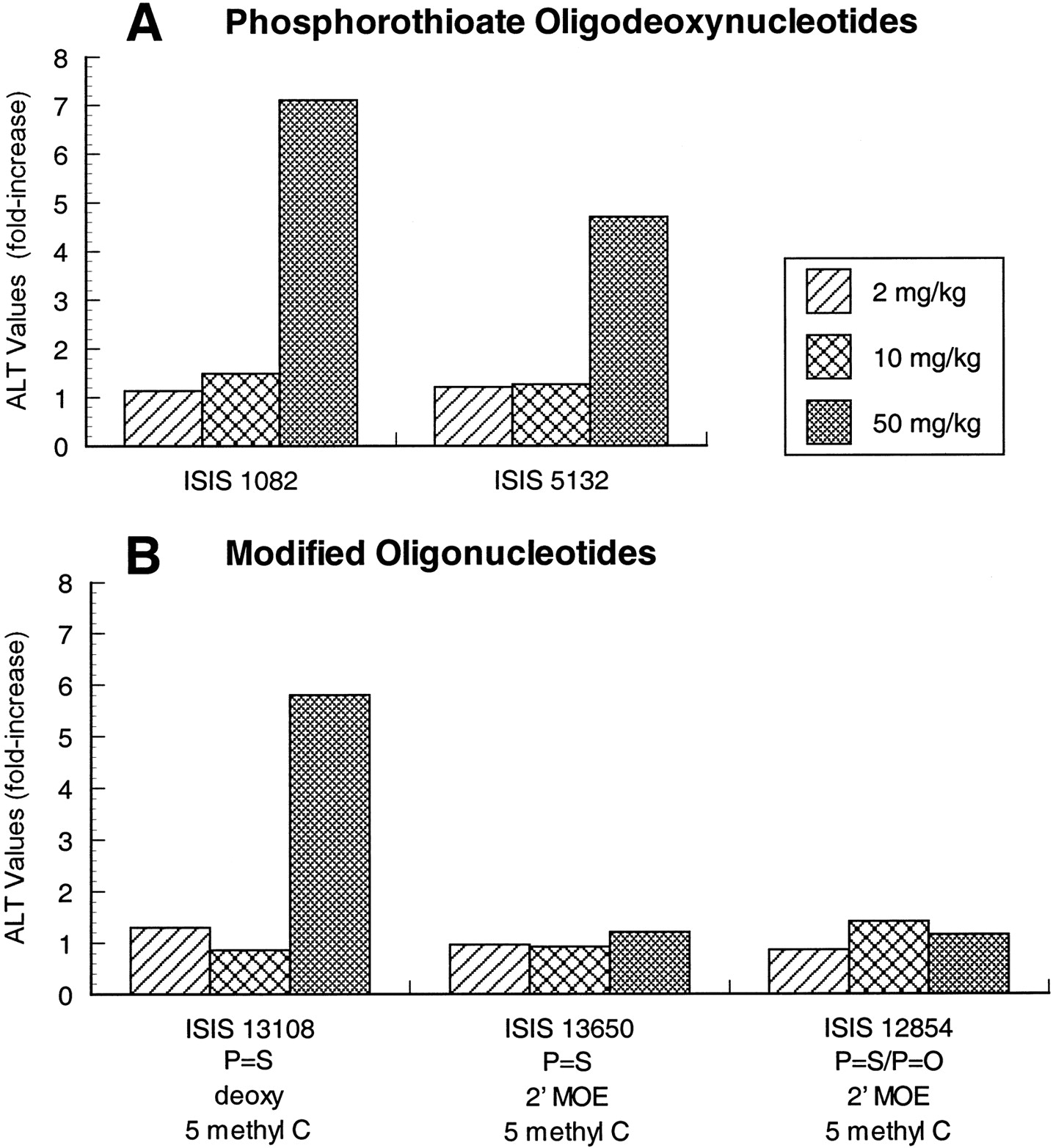
Effect of oligonucleotide treatment on ALT values in male mice. The effects of several phosphorothioate oligodeoxynucleotides (A) are compared with modified oligonucleotides (B) containing 5-methyl C substitutions or 2′-MOE substituents. Oligonucleotides were administered by every other day administration for 14 days. Blood was collected for serum chemistry analysis 24 h after the last dose. Values represent the relative change in mean ALT value from control group of five or six mice in each case. Standard deviation was generally <35% of the mean.

Effect of oligonucleotide treatment on platelet count in male mice. The effects of several phosphorothioate oligodeoxynucleotides (A) are compared with modified oligonucleotides (B) containing 5-methyl C substitutions or 2′-MOE substituents. Oligonucleotides were administered by every-other-day administration for 14 days. Blood was collected for hematology analysis 24 h after the last dose. Values represent the relative change in mean organ weight from control group of five or six mice in each case. Standard deviation was <50% of the mean in all cases.
In comparison, there was no increase in liver weight or serum transaminase, and no thrombocytopenia in mice treated with oligonucleotides containing 2′-MOE and 5-methyl C modifications. Liver weights or serum ALT levels were unaffected in mice treated with ISIS 13650 or ISIS 12854, indicating less hepatotoxicity relative to phosphorothioate oligodeoxynucleotides (Figs. 5B and 6B). Similarly, no animals treated with phosphorothioate oligonucleotides or mixed backbone oligonucleotides containing 2′-MOE experienced a decrease in platelet counts (ISIS 13650 or ISIS 12854) (Fig. 7B). Again, this apparent increase in the tolerability of 2′-MOE modified oligonucleotides is not explained by the absence of oligonucleotide exposure in target organs because there is still appreciable concentrations in liver (other organs) and concentrations are higher than have been reported for ISIS 5132 with the same dose regimen.
Interestingly, the phosphorothioate oligodeoxynucleotide that contained the 5-methyl C modification only, ISIS 13108, produced increases in liver weight and serum ALT levels similar to ISIS 1082 (Figs. 5B and6B). There was also a dose-dependent decrease in platelet count in ISIS 13108-treated mice, but the severity was decreased relative to ISIS 1082 (Fig. 7B).
Discussion
Antisense phosphorothioate oligodeoxynucleotides have been shown to possess very specific and discrete activity against the targeted mRNA in cell culture, as well as animal models of disease, that is dependent on the nucleotide sequence (Monia and Dean, 1998). Like any class of drugs, there are potential nonspecific effects produced by administration of the compounds. The biophysical properties of these compounds allow interaction with other components, such as serum or cellular proteins. These hybridization-independent interactions can potentially alter protein function. Immune stimulation is a well-characterized hybridization-independent effect of unmodified phosphorothioate oligodeoxynucleotides that may arise from the interaction of these agents with surface receptors leading to the activation of certain cells, such as B cells (Liang et al., 1996). In the absence of very specific sequence motifs, most of these oligonucleotide-protein interactions are relatively nonspecific, are of low affinity (generally micromolar binding constants), and are potentially lower potency compared with the intended pharmacologic effects (Srinivasan et al., 1995; Bennett et al., 1997). For example, an antisense oligonucleotide specific for ICAM-1 reduced inflammation in several animal models of inflammatory disease at doses ranging from 1 to 10 mg/kg, whereas toxicity in mice occurs at doses of 20 to 100 mg/kg (Bennett et al., 1997, Bennett and Condon, 1998). Although potency for antisense effects is generally greater than hybridization-independent effects, such properties can complicate interpretation of pharmacologic activity in animal models, particularly in a sensitive species such as mice.
Immune stimulation by phosphorothioate oligodeoxynucleotides has been studied most extensively in vitro with isolated splenocytes or peripheral blood leukocytes (Pisetsky and Reich, 1994; Krieg et al., 1995; Liang et al., 1996). In isolated cell populations, immune stimulation is characterized by increased mitogenic activity (i.e., increased DNA synthesis), polyclonal antibody production (IgG and IgM), and stimulation of cytokine release (IL-6, IL-12, and interferon-γ) (Pisetsky and Reich, 1994; Branda et al., 1996; Klinman et al., 1996;Liang et al., 1996). Investigations with partially purified cell populations or specific cell activation inhibitors have indicated that these effects are primarily due to activation of B cells. Nonlymphoid cells may contribute to immune stimulation, as human keratinocytes have been shown to produce IL-1α when exposed to phosphorothioate oligodeoxynucleotides (Crooke et al., 1996).
Mice treated with high doses of phosphorothioate oligodeoxynucleotide exhibit alterations that are consistent with the immunostimulatory properties described in vitro. The mitogenic effect on B cells is reflected in dose-dependent increases in spleen weights and histologic evidence of lymphoid hyperplasia (Branda et al., 1993; Sarmiento et al., 1994; Henry et al., 1997b; Monteith et al., 1997). Increases in circulating concentrations of serum immunoglobulins and cytokines also have been reported (Zhao et al., 1995; Branda et al., 1996). Additional evidence of immune stimulation in mice treated with these compounds includes cell infiltrates in multiple organs (Sarmiento et al., 1994;Henry et al., 1997b).
Immunohistochemical staining was used to further characterize the cell infiltrate in liver and kidney. Areas of cell infiltrates stained positive for the presence of IL-6, a member of the proinflammatory cytokine family. Production of IL-6 has been reported previously following activation of B cells in vitro or in serum of mice treated with phosphorothioate oligonucleotides (Klinman et al., 1996; Zhao et al., 1997). However, it is not clear if its production is a consequence of direct oligonucleotide exposure, or possibly secondary to some other cytokine or cellular activation process. Infiltrating cells stained positive for LFA-1, CD18, and Mac-1, but not Ly6G(Gr1) suggesting that the infiltrates were predominantly monocytic. Expression of surface adhesion molecules ICAM-1 and vascular cell adhesion molecule-1 were increased on vascular endothelium, as well as infiltrating cells, consistent with the migration of cells from the vasculature into tissues. These findings further confirm the correlation between in vitro and in vivo effects of phosphorothioate oligodeoxynucleotides, and strongly suggest that the cellular infiltrates in tissues are a component of the immune stimulation produced by these molecules. However, it is worth mentioning that data collected in this study represent a single time point (24 h after last dose). Typical of immune reactions, this response to phosphorothioate oligodeoxynucleotides is probably a dynamic process subject to transient sequence of events.
The different phosphorothioate oligodeoxynucleotides examined in this study demonstrated a spectrum of potency for immune stimulation. Qualitatively, the extent of splenomegaly corresponded with the severity of cell infiltrates (Henry et al., 1997c, Monteith et al., 1997). The sequence of ISIS 12449 was designed to produce optimal B-cell stimulation based on sequence motifs characterized by Krieg and Yamamoto (Yamamoto et al., 1994; Krieg et al., 1995). This oligonucleotide contains four CpG dinucleotide motifs, one of which contains the optimal flanking sequences for immune stimulation. This CpG motif (AACGTT) with two 5′ purines and two 3′ pyrimidines is capable of forming palindromic structures, which appear to enhancing the potency of this hybridization-independent effect. Indeed, treatment of mice with ISIS 12449 produced the most extensive immune stimulation observe among the compounds examined in this study. ISIS 1082 and ISIS 5132 are targeted toward the UL13 mRNA of herpes simplex virus-1 and human C-raf kinase, respectively. These oligonucleotides do not contain the optimal immunostimulatory sequence, and treatment of mice produced an intermediate level of immune stimulation. In contrast, ISIS 12959, which is targeted toward baboon C-raf kinase, and has the same sequence as ISIS 5132 with the exception of two base changes, produced the least amount of stimulation. One of the base substitutions changes a CpG motif to a cytosine-adenosine motif, suggesting that even nonoptimal CpG dinucleotide motifs contribute to the potency of immune stimulation.
Chemical modification of the above-mentioned phosphorothioate oligodeoxynucleotide sequences studied produced much less evidence of immune stimulation. Substitution of 5-methyl C for cytosine (ISIS 13108) produced much less splenomegaly or cell infiltrates compared with unmodified parent compound, ISIS 1082. This confirms the observation by Krieg et al. (1996) that methylation of cytosine reduces the in vitro mitogenic activity of phosphorothioate oligodeoxynucleotides. The level of immune stimulation in mice treated with oligonucleotides containing a combination of 5-methyl C substitution and 2′-MOE modifications was also low (ISIS 13650 and ISIS 12854). These modified oligonucleotides produced essentially no splenomegaly, even at the 50-mg/kg dose. These in vivo data are consistent with other reports showing that 2′-methoxy modifications also reduce the level of immune stimulation relative to phosphorothioate oligodeoxynucleotides (Zhao et al., 1995). Oligonucleotides containing 2′-MOE modifications only (i.e., no 5-methyl C) have been tested for mitogenic activity in vitro. Although it appears that the 2′-alkoxy modification is able to reduce the extent of immune stimulation to some extent, the most significant differences were obtained with a combination of 5-methyl C and 2′-MOE modifications (R. Crooke and K. Lemonidis, Isis Pharmaceuticals, manuscript in preparation).
The exact mechanism by which these modifications reduce immune stimulation is unclear. However, it has been suggested that the presence of 5-methyl C in an oligonucleotide more closely resembles mammalian DNA compared with unmethylated cytosine that is more prevalent in bacterial DNA (Krieg et al., 1996). Because bacterial DNA is immunostimulatory and mammalian DNA is not, it has been suggested that there has been some phylogenetic mechanism developed to distinguish this subtle difference in structure (Krieg et al., 1995). Given the hypothesis that immune stimulation results from oligonucleotide-protein interactions, one possible explanation for the effects of cytosine methylation might be that the increased lipophilicity at these sites prevents interaction with a key protein involved in the recognition of foreign DNA. Similarly, modification of the 2′ position on ribose with alkyl substituents, may alter other protein-binding interactions.
Besides a decrease in immune stimulation, modified oligonucleotides also resulted in a reduction in other general toxicity parameters. Oligonucleotides containing both 5-methyl C and 2′-MOE modifications had essentially no effect on liver weights, ALT level, or platelet counts. Although partially attenuated in the oligonucleotide containing 5-methyl C substitution, there was still an increase in liver weights and in serum transaminases, and decrease in platelet counts. The decrease in immune stimulation and other toxicity parameters in 2′-MOE-modified oligonucleotides is significant considering this modification does not alter the tissue distribution profile, and actually increases the tissue half-life (Cummins et al., 1995) due to increased resistance to nuclease degradation. As a result, organ concentrations of oligonucleotide containing the 2′-MOE modification at a defined dose regimen are greater than achieved by unmodified phosphorothioate oligodeoxynucleotides.
The findings in this study demonstrated a close correlation between the cellular infiltrate and splenomegaly in both severity and dose response. Investigation of local cytokine production, cell adhesion molecule expression, and lymphocyte surface markers provided new insights into the basis for cellular infiltrate and revealed a pattern of immune stimulation in mice that is consistent with the production of proinflammatory cytokines from isolated splenocytes. Variability in the potency for immune stimulation between phosphorothioate oligodeoxynucleotides of different sequence in mice was also dependent upon sequence motifs similar to that described in vitro. Importantly, chemical modification of phosphorothioate oligonucleotides can effectively reduce the potency of immunostimulatory properties. Ultimately, these modifications may help to reduce the hybridization-independent effects of antisense oligonucleotides, thus increasing the potential therapeutic index.
Footnotes
-
Send reprint requests to: Scott P. Henry, Ph.D., D.A.B.T., Associate Director, Toxicology, Isis Pharmaceuticals, Inc., 2292 Faraday Ave., Carlsbad, CA 92008. E-mail:Shenry{at}isisph.com
- Abbreviations:
- IL
- interleukin
- CpG
- cytosine-guanine
- 5-methyl C
- 5-methyl cytosine
- MOE
- methoxyethyl
- ALT
- alanine aminotransferase
- LFA
- leukocyte function-associated antigen
- ICAM-1
- intercellular adhesion molecule-1; Mac-1
- Received May 27, 1999.
- Accepted October 25, 1999.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}