


Abstract
The role of p38 mitogen-activated protein (MAP) kinase, and extracellular-regulated protein kinase -1 and -2 in regulating constitutive apoptosis and interleukin (IL)-5-induced survival of human eosinophils have been investigated. Two populations of donors were identified whose eosinophils, in the absence of exogenous cytokines, underwent apoptosis at different rates. Eosinophils were thus arbitrarily classified as either “fast”- or “slow”-dying cells, where greater or less than 15% of the cells were apoptotic at 2 days, respectively. The selective p38 MAP kinase inhibitor, SB 203580, increased constitutive eosinophil apoptosis in both populations (EC50 ∼2 μM) as evinced from morphological analysis, flow cytometry, and DNA laddering. The ability of SB 203580 to kill eosinophils was not due to nonspecific toxicity or through the inhibition of prostanoid or leukotriene production. Exposure of eosinophils to IL-5, at a concentration (10 pM) that enhanced survival maximally, abolished SB 203580-induced apoptosis. In contrast PD 098059, which selectively blocks MAP kinase kinase (MEK) 1, did not affect apoptosis of fast- or slow-dying eosinophils, or the enhanced survival of cells effected by IL-5. Collectively, these results suggest that: 1) the basal activity of p38 MAP kinase may regulate the survival of cytokine-deprived eosinophils through inhibition of apoptosis, 2) the enhancement of eosinophil survival effected by IL-5 is mediated by a mechanism(s) divorced from the activation of p38 MAP kinase, and 3) neither spontaneous eosinophil apoptosis nor their enhanced survival by IL-5 involves the activation of MEK-1.
Classically, eosinophils were thought to be involved in immune defense against parasitic infection. However, it is now recognized that the accumulation and activation of eosinophils within tissue, in particular the airway mucosa, is implicated in the pathogenesis of a number of diseases, including asthma and eosinophilic pneumonias (Giembycz and Lindsay, 1999). Of great importance in determining the number of eosinophils found in the blood and in tissues is the balance between cell maturation and cell death (Yousefi et al., 1997; Walsh, 1997). Eosinophils cultured in the absence of cytokines undergo apoptosis or programmed cell death (Yamaguchi et al., 1991; Stern et al., 1992), a process that can be inhibited by a number of cytokines principally interleukin (IL)-3, IL-5, and granulocyte/macrophage colony-stimulating factor (Yamaguchi et al., 1991; Tai et al., 1991; Stern et al., 1992). These in vitro observations are corroborated by an in vivo study where IL-5 was shown to orchestrate the eosinophilia in human nasal polyps through its ability to inhibit apoptosis (Simon et al., 1997).
Apoptosis is characterized by specific biochemical and morphological changes including cell shrinkage, surface blebbing, chromatin condensation, and endonuclease-catalyzed DNA breakdown. These events then are followed by fragmentation of the cell into discrete apoptotic bodies that are recognized and engulfed by phagocytic cells (Stern et al., 1992, 1996). Apoptosis is distinct from necrosis, which is characterized by cell lysis and the uncontrolled release of cellular contents that may be harmful to surrounding tissues. For these reasons, the induction of apoptosis may be a potential therapeutic objective in the resolution of eosinophilic inflammation (Anderson, 1996).
Although eosinophils undergo apoptosis by processes that can be inhibited by certain cytokines, little is known of the precise signaling pathways that control this process(es). Recently, evidence has emerged that a group of proline-directed, protein serine/threonine kinases, collectively known as mitogen-activated protein (MAP) kinases, play a role in the promotion (Frasch et al., 1998; Graves et al., 1996; Verheij et al., 1996; Sutherland et al., 1996; Kummer et al., 1997; Schwenger et al., 1997) and inhibition (Sutherland et al., 1996, Gardner and Johnson, 1996; Nemoto et al., 1998) of apoptosis. Currently, three subfamilies have been biochemically classified: the extracellular-regulated kinases (ERKs) ERK-1 and ERK-2, the 46- and 54-kDa c-jun N-terminal kinases (JNK; JNK-46 and JNK-54, respectively), and the p38 MAP kinase family. ERK-1 and ERK-2 are typically activated by mitogens and growth factors whereas JNK and p38 MAP kinases are sensitive to pro-inflammatory cytokines, heat shock, hyperosmolarity, and cellular stress (Denhardt, 1996; Kyriakis and Avruch, 1996). A survey of the current literature suggests that the regulation of cell survival by MAP kinases is probably cell-specific and almost certainly dependent on the stimulus, which will ultimately lead to different profiles of enzyme activity. For example, activation of JNK and/or p38 MAP kinase has been proposed to mediate CD95-, ceramide-, and sodium salicylate-induced apoptosis in human and other mammalian cell lines (Verheij et al., 1996; Schwenger et al., 1997). Similarly, activation of ERK-2 correlates with apoptosis of WEHI-231 B-lymphoma cells in response to ligation of the antigen receptor, whereas in the same cells the concurrent engagement of CD40 results in the additional activation of JNK and p38 MAP kinase and prevents antigen-driven apoptosis (Sutherland et al., 1996).
In eosinophils, evidence is available that picomolar concentrations of IL-5 promote the phosphorylation of ERK-1 and/or ERK-2 (Pazdrak et al., 1995b; Bates et al., 1996; Hiraguri et al., 1997; Coffer et al., 1998) by a mechanism that is inhibited by transforming growth factor-β (Pazdrak et al., 1995a). Immediate upstream regulators of ERK-1 and ERK-2 in eosinophils include the enzyme mitogen-activated protein kinase kinase (MEK)-1. This protein is a member of the MEK superfamily that, in turn, is controlled by a protein kinase cascade involving the sequential activation of lyn, Ras, and Raf-1 (Pazdrak et al., 1995b). The finding that antisense deoxyoligonucleotides directed against Raf-1 and lyn promote death of IL-5-treated human eosinophils suggests that activation of ERK-1 and/or ERK-2 via thelyn-Ras-Raf-1-MEK cascade may play a primary role in opposing spontaneous eosinophil apoptosis (Pazdrak et al., 1995b;1998). In addition, IL-5 activates the Jak2-STAT1 pathway in human eosinophils, which also has been implicated in eosinophil survival (Bates et al., 1996; Pazdrak et al., 1995a, 1998).
Given the role of MAP kinases in regulating the longevity of a variety of cells, we elected to assess the extent to which p38 MAP kinase MEK-1 are involved in apoptosis of cytokine-deprived human eosinophils and the survival-enhancing activity of IL-5.
Materials and Methods
Isolation of Human Eosinophils.
Venous blood (50–100 ml) from healthy or asthmatic individuals not receiving glucocorticoid therapy was collected into 10 to 20 ml of acid citrate dextrose anticoagulant. The supernatant obtained after sedimentation with 3% hydroxyethyl starch was layered onto Ficoll and centrifuged at 500g for 30 min at 18°C. The mononuclear cell layer was discarded and the pellet containing granulocytes and red blood cells was washed in Hanks’ balanced salt solution. Contaminating red blood cells were lysed by hypotonic lysis. The remaining granulocytes were washed, counted, and resuspended in 300 μl RPMI 1640 containing 2% fetal calf serum (FCS) and 5 mM EDTA (RPMI/FCS/EDTA). Eosinophils were purified from neutrophils using immunomagnetic anti-CD16 antibody-conjugated beads (1 μl of beads per 2 × 106 neutrophils). After the addition of beads, cells were incubated at 4°C for 40 min, resuspended in 6 ml RPMI/FCS/EDTA, loaded onto a separation column positioned within a magnetic field, and eluted with 40 ml RPMI/FCS/EDTA. The CD16+ cells are retained by the column while the eluted eosinophils were collected, washed in RPMI 1640, counted, and resuspended at 106 cells/ml. Using this method eosinophil purity was >99%. The cells then were cultured in RPMI 1640 medium with 10% FCS plus antibiotics (RPMI/FCS).
Determination of Eosinophil Apoptosis and Viability.
Eosinophil apoptosis was determined by propidium iodide (PI) staining of DNA fragmentation and flow cytometry (FACScan, Becton Dickinson, San Jose, CA). Briefly, the 200-g cell pellet was gently resuspended in hypotonic fluorochrome solution (PI 25 μg/ml in 0.1% sodium citrate plus 0.1% Triton X-100). The tubes were left in the dark overnight at 4°C before flow cytometric analysis. Apoptosis was confirmed by morphological analysis of cells spun onto cytospin slides and stained with May-Grünwald-Giemsa. Eosinophil viability was assessed by PI exclusion in isotonic buffer and analyzed by flow cytometry. Cells impermeable to PI were considered viable.
Determination of DNA Fragmentation.
Oligonucleosomal DNA fragmentation, a characteristic feature of eosinophil apoptosis, was analyzed by agarose gel DNA electrophoresis. Eosinophils (106/ml in RPMI 1640) were cultured in the absence or presence of SB 203580, SB 202190, PD 098059, and IL-5 for 24 h, pelleted by centrifugation (200g × 7 min), and suspended in 0.5 ml of digestion buffer (10 mM Tris-HCl pH 8.0, 100 mM NaCl, 25 mM EDTA, 0.5% SDS, and 0.2 mg/ml proteinase K) for 12 h at 50°C. Digested cells were extracted with phenol/chloroform/isoamyl alcohol (25:24:1; v/v/v) and buffered with Tris-EDTA buffer (pH 8.0) and chloroform/isoamyl alcohol (24:1; v:v). DNA was then precipitated with 2.5 M ammonium acetate and two volumes of ethanol at −20°C for at least 24 h. The DNA precipitates were recovered by centrifugation at 12,000g for 30 min. After drying, DNA was dissolved in TE buffer (10 mM Tris-HCl, 5 mM EDTA, pH 8.0), mixed with orange G, and loaded on to 2.0% agarose gels containing 0.5 μg/ml ethidium bromide. Electrophoresis was carried out in 40 mM Tris base, 1.1 mM glacial acetic acid, and 1 mM EDTA, pH 8.0. After electrophoresis, gels were visualized by ultraviolet light and photographed.
Extraction of Cytosolic Proteins
Eosinophils were incubated at 106/ml in RPMI/FCS in the absence and presence of IL-5 (10 pM). The incubations were stopped at the appropriate times (indicated in the text and figure legends) by centrifugation, resuspension in 50 μl of medium, and addition of equal volumes of ice-cold lysis buffer (20 mM Tris base, pH 7.4, 1% Triton X-100, 0.5% w/v sodium deoxycholate, 0.1% w/v SDS, 100 mM NaCl, 1 mM EDTA, 1 mM phenylmethylsulfonyl fluoride, 2 mM sodium orthovanadate, 10 μg/ml leupeptin, 25 μg/ml aprotinin, 1.25 mM NaF, 1 mM sodium pyrophosphate). After extraction by incubation on ice for 15 min, samples were centrifuged and the resulting supernatant was boiled for 5 min in sample buffer (62.5 mM Tris-HCl, 20% glycerol, 2% SDS, and 10 mM 2-mercaptoethanol) and stored at −70°C until used for Western immunoblot analyses.
Western Blot Analysis.
The activation status of p38 MAP kinase, ERK1, ERK2, JNK-46, and JNK-54 was assessed by Western immunoblot analysis using antibodies that recognize the dual phosphorylated (activated) form of the enzymes. Briefly, protein samples were separated by SDS-polyacrylamide gel electrophoresis on 10% polyacrylamide gels and then transferred to nitrocellulose (HybondECL) for 2 h at 1A in transblotting buffer (183 mM glycine, 25 mM Tris base, and 20% methanol). The nitrocellulose was incubated for 1 h in TBS-T (25 mM Tris base, 150 mM NaCl, 0.1% Tween 20, pH 7.4) containing 5% (w/v) nonfat dry milk to block nonspecific antibody binding, and incubated overnight in TBS-T containing 5% BSA and the relevant primary antibody. Membranes were washed with TBS-T (5 × 5 min) and incubated with either horseradish peroxide-linked antirabbit IgG (diluted 1:4000) in TBS-T/5% nonfat dry milk for 1 h at room temperature. The nitrocellulose then was washed in TBS-T (5 × 5 min) and developed using enhanced chemiluminescence (ECL) Western blotting detection agents on Kodak X-OMAT-S film. Relevant bands were quantified by laser-scanning densitometry.
Drugs, Chemicals, and Analytical Reagents.
Actinomycin D, aprotinin, fetal calf serum, flurbiprofen, indomethacin, leupeptin, PI, RPMI 1640, and Triton X-100 were purchased from Sigma Chemical Company (Poole, Dorset, UK). HybondECL, HyperfilmECL, ECL Western blotting reagents, and Rainbow protein molecular weight markers were obtained from Amersham International (Buckinghamshire, UK). All other reagents were obtained as follows: anti-CD16 microbeads and magnetic cell separation system (Miltenyi Biotec Ltd, Surrey, UK); AquaPhenol (Appligene-Oncor, Co., Durham, UK); Ficoll-Paque (Pharmacia AB, Uppsala, Sweden); horseradish peroxidase-conjugated secondary antirabbit antibodies (Santa-Cruz Biotechnology, Ltd., London, UK); φX174/HaeIII DNA marker and proteinase K (Promega Corp., Madison, WI); and phosphoPlus JNK (Thr183/Tyr185), p38 MAP kinase (Thr180/Tyr182), and ERK-1/ERK-2 (Thr202/Tyr204) antibody kits (New England Biolabs, Inc., Beverly, MA). Human recombinant IL-5, BAY x1005, L-745,337, PD 098059, SB 203580, and SB 202190 were purchased from R&D Systems (Abingdon, UK), Bayer Plc (Stoke Poges, UK), Merck Frost Inc., (Montreal, Canada), Calbiochem-Novobiochem (Nottingham, UK), Alexis Corporation (Laüfelfingen, Switzerland), and Smith-Kline-Beecham (King of Prussia, PA), respectively. Stocks of all inhibitors were dissolved in dimethyl sulfoxide and diluted in the appropriate medium. At the highest maximally effective drug concentration, dimethyl sulfoxide did not exceed 0.1% v/v, which had no affect on eosinophil viability.
Statistical Analysis.
Results in the text and figures are expressed as mean ± S.E.M. of n independent determinations. Where appropriate, statistical evaluation was performed by ANOVA supported by Tukey-Kramer multiple comparison test. The null hypothesis was rejected when P < .05.
Results
Characteristics of Human Eosinophil Apoptosis.
Initially, studies were undertaken to monitor the rate of apoptosis in human eosinophils over 5 days of culture. In the absence of cytokines, eosinophils have been previously reported to undergo “spontaneous” apoptosis over a period of 1 to 3 days (Yamaguchi et al., 1991; Yousefi et al., 1994; Walsh, 1997). Although we observed a similar profile of apoptosis in the majority of donors (Fig.1A), we also identified a population of individuals whose circulating eosinophils exhibited low levels of apoptosis over the same time frame (Fig. 1B). A comparison of eosinophils isolated three times from six individuals over a period of 11 to 35 weeks showed that the percentage that became apoptotic after 48 h of culture remained constant (Table1), indicating that the rate of apoptotic death is intrinsic to the donors. Eosinophils were subsequently classified as either “fast”- or “slow”-dying cells, using an arbitrary criterion where greater or less than 15% of the cells were apoptotic at 48 h respectively. Of 35 individuals examined, eosinophils from 22 of them died rapidly whereas the remainder died apoptotically at a relatively slow rate. A comparison of the ratio of fast- to slow-dying eosinophils taken from healthy (1 of 9) and asthmatic (12 of 26) subjects showed that there was no significant difference in the number of apoptotic cells after 48 h of culture (35.2 ± 4.0% and 26.4 ± 4.2%, respectively;P > .05).

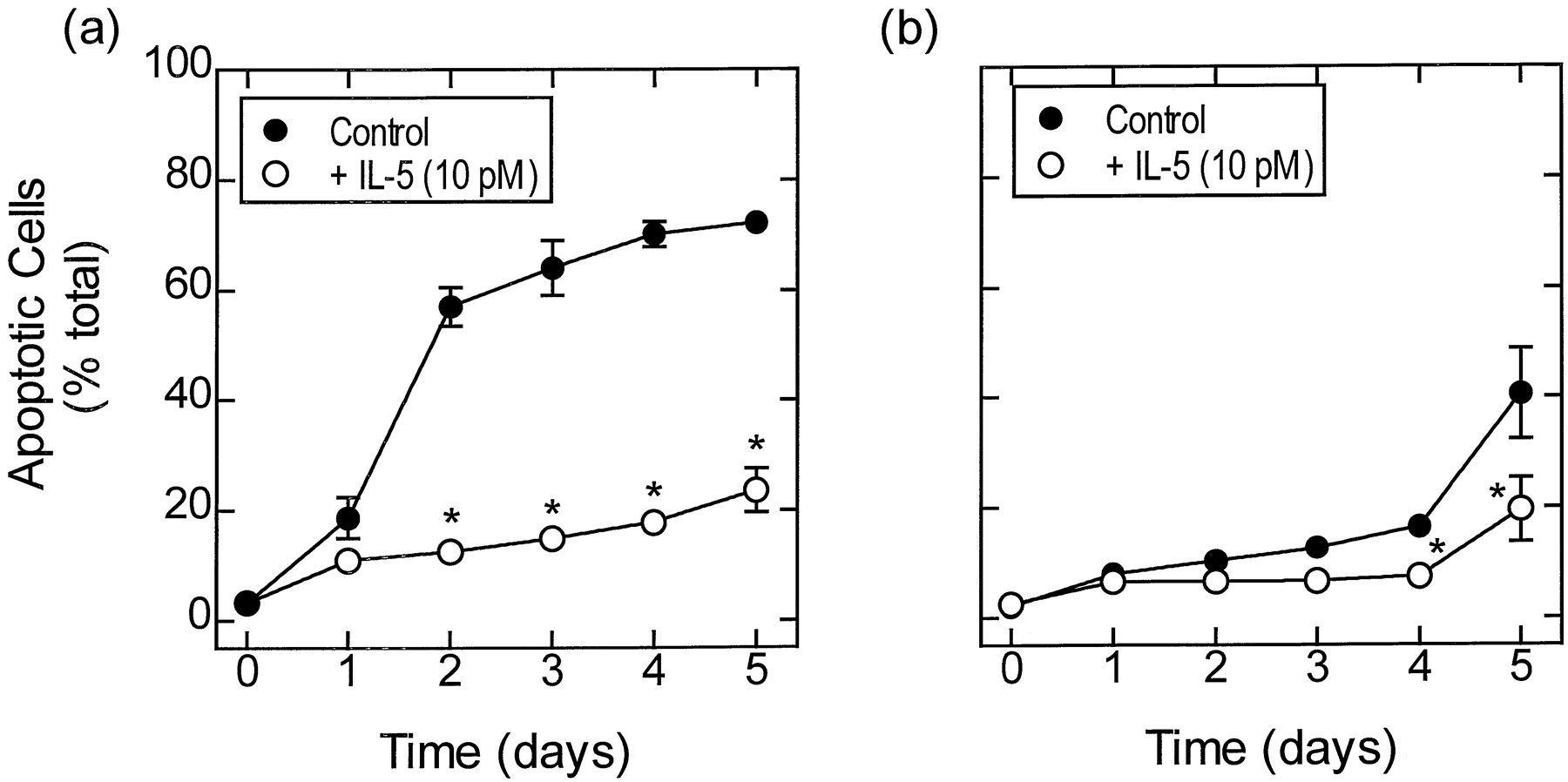
Effect of IL-5 on spontaneous apoptosis of human eosinophils in vitro. Eosinophils were cultured for 5 days in the absence (●) or presence (○) of 10 pM IL-5. (A) and (B) show the time course of apoptosis from donors whose eosinophils died quickly and slowly, respectively. Apoptosis was assessed by flow cytometry measuring the relative DNA content of PI-stained eosinophils. Each data point represents the mean ± S.E.M. of three independent determinations using eosinophils from different donors. SeeMaterials and Methods for further details. *P< .05 significant inhibition of apoptosis by IL-5 compared with time-matched control cells.
Reproducibility of human eosinophil apoptosis in vitro within donors
Consistent with previous reports, culture of fast-dying human eosinophils with IL-5 (10 pM) enhanced viability (Fig. 1); this effect was concentration-dependent (EC50 ∼0.1 pM) and was the result of inhibiting apoptosis (data not shown). At the maximally effective concentration of IL-5 (10 pM) the percentage of apoptotic eosinophils present in the cultures was significantly reduced at days 1 (18.7–11.1%), 2 (57.1–12.5%), 3 (64.1–14.9%), 4 (70.2–17.8%), and 5 (72.8–23.6%) when compared with cytokine-deprived cells (Fig. 1A). IL-5 (10 pM) also caused a small but significant reduction in the apoptosis of slow-dying eosinophils (15.7–7.8% and 40.6–19.5% after 4 and 5 days in culture, respectively; Fig. 1B). Qualitatively identical results were obtained from morphological analyses of fast-dying eosinophils spun on to cytospin slides. Thus, after 2 days’ culture, the percentage of apoptotic cells in the absence and presence of IL-5 (10 pM) was 36.2 ± 5.2 and 7.0 ± 0.5%, respectively (n= 5, P < .05).
Effect of SB 203580 and SB 202190 on Eosinophil Longevity.
To examine the role of p38 MAP kinase in eosinophil apoptosis, a pharmacological approach was initially adopted where eosinophil viability was assessed in cultures supplemented with purportedly selective inhibitors of this enzyme family.
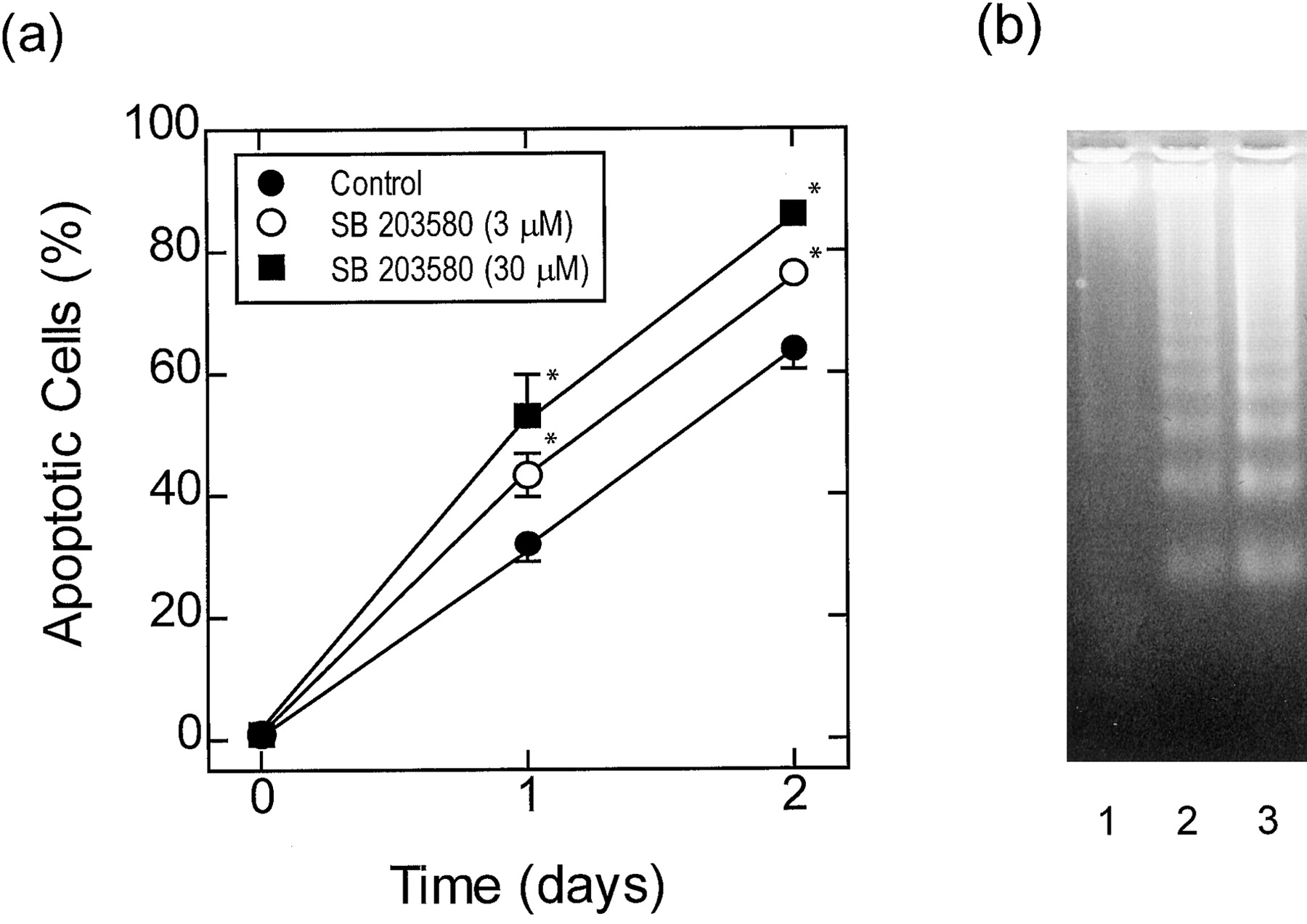
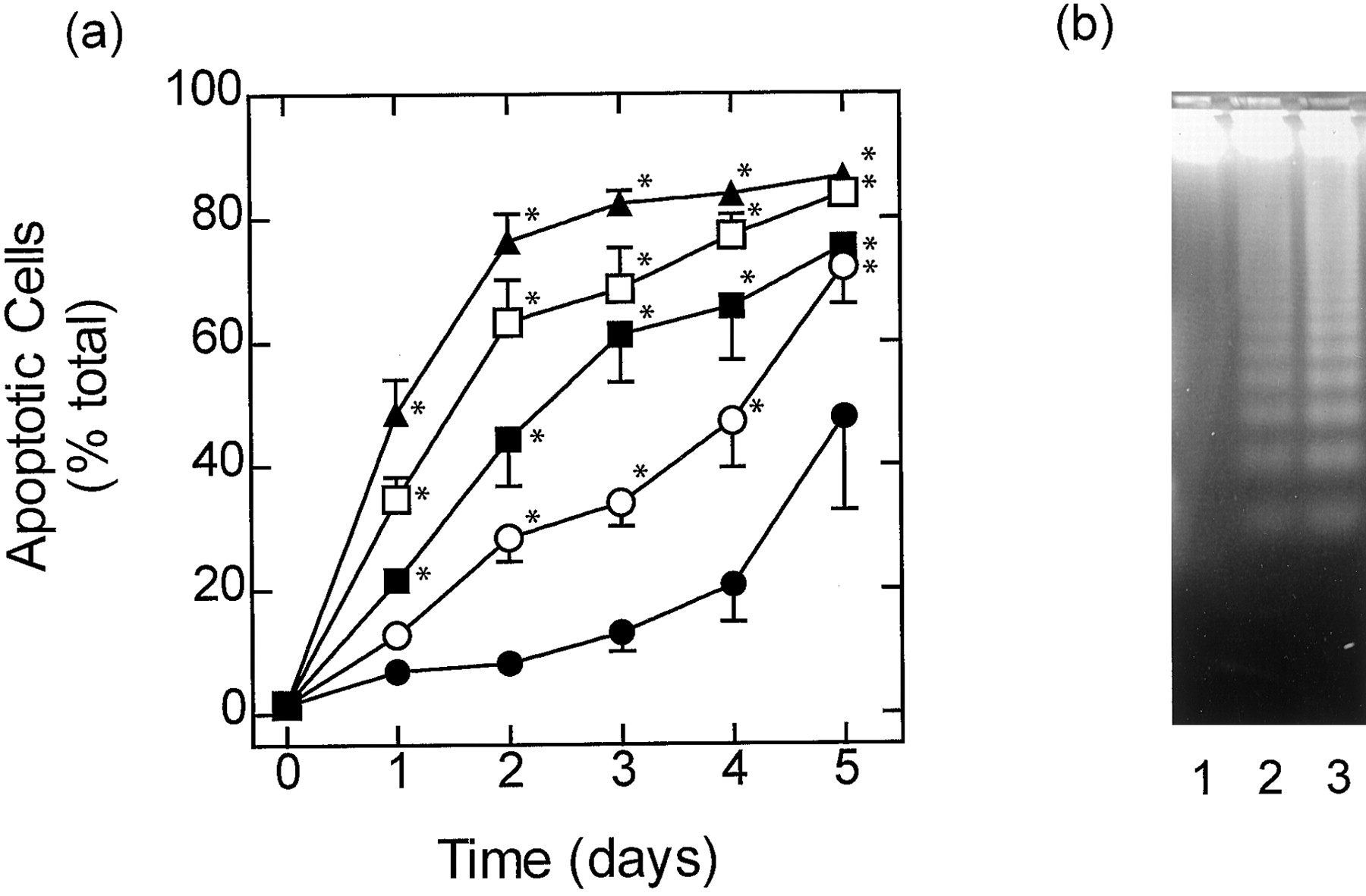
In both fast- and slow-dying cells, SB 203580 (0.1–30 μM), an inhibitor of the α and β isoforms of p38 MAP kinase, augmented constitutive apoptosis in a concentration-dependent manner as assessed by flow cytometry (Figs. 2A, 3A, and 4A); the EC50values for SB 203580 in eosinophils that died slowly were 2.6, 1.95, and 1.52 μM on days 2, 3, and 4, respectively (Fig.5). Apoptosis was also inferred from the enhanced DNA fragmentation observed in fast- and slow-dying eosinophils exposed to SB 203580 (10 μM) for 2 days when compared with vehicle-treated cells under identical experimental conditions (Figs. 2B and 3B). Another inhibitor of p38 MAP kinase, SB 202190, also promoted apoptosis of slow-dying eosinophils with similar potency to SB 203580 (EC50 = 1 μM; Fig. 4B). Significantly, apoptosis effected by both p38 MAP kinase inhibitors was prevented in eosinophils treated with IL-5 (Fig. 4, A and B). Using the complementary techniques of flow cytometry and DNA fragmentation it was established that the antiapoptotic action of IL-5 was concentration-dependent with complete protection achieved at 10 pM (Fig. 4).

Effect of SB 203580 on cytokine-deprived eosinophils that undergo rapid constitutive apoptosis. Cytokine-deprived eosinophils were cultured for 2 days in the absence (●) and presence of 3 (○) and/or 30 μM (▪) SB 203580 and apoptosis was assessed by flow cytometric analysis of PI-stained DNA content (A) and by oligonucleosomal DNA fragmentation (B). In (A) each data point represents the mean ± S.E.M. of four independent determinations using eosinophils from different donors. The gel shown in (B) is typical of three determinations. See Material and Methods for further details. Lane 1, freshly prepared eosinophils; lane 2, cytokine-deprived eosinophils cultured for 2 days; lane 3, cytokine-deprived eosinophils cultured for 2 days in the presence of SB 203580 (10 μM). *P < .05 significant augmentation of apoptosis effected by SB 203580 relative to time-matched control.

Concentration-dependence of SB 203580-induced apoptosis of slow-responding eosinophils. The percentage of apoptotic cells at day 2, 3, and 4 in the experiment shown in Fig. 5A are re-plotted as a function of the concentration of SB 203580; each data point represents the mean ± S.E.M. of four independent determinations using eosinophils from different donors. The EC50 of SB 203580 (∼2 μM) was essentially independent of the day apoptosis was measured.

Effect of SB 203580 on cytokine-deprived eosinophils that undergo slow constitutive apoptosis. Cytokine-deprived eosinophils were cultured for 5 days in the absence (●) and presence of 1 (○), 3 (▪), 10 (■), and/or 30 μM (▴) SB 203580 and apoptosis was assessed by flow cytometric analysis of PI-stained DNA content (A) and by oligonucleosomal DNA fragmentation (B). In (A) each data point represents the mean ± S.E.M. of four independent determinations using eosinophils from different donors. The gel shown in (B) is typical of three determinations. See Materials and Methods for further details. Lane 1, freshly prepared eosinophils; lane 2, cytokine-deprived eosinophils cultured for 2 days; lane 3, cytokine-deprived eosinophils cultured for 2 days in the presence of SB 203580 (10 μM). *P < .05 significant augmentation of apoptosis effected by SB 203580 relative to time-matched control.

Effect of IL-5 on the pro-apoptotic activity of SB 203580 and SB 202190 in slow-dying eosinophils. Eosinophils were cultured for 2 days in medium containing SB 203580 (0.1–30 μM; A) or SB 202190 (0.1–30 μM; B) in the absence (●) and presence of 100 fM (○), 10 pM (▪), and/or 1 nM (■) IL-5 and apoptosis was assessed by flow cytometric analysis of PI-stained DNA content (A and B) and by oligonucleosomal DNA fragmentation (C). In (A) and (B) each data point represents the mean ± S.E.M. of four independent determinations using eosinophils from different donors. The gel shown in (B) is typical of three determinations. See Materials and Methods for further details. Lane 1, cytokine-deprived eosinophils cultured for 2 days in the presence of SB 203580 (10 μM); lane 2, eosinophils cultured for 2 days in the presence of IL-5 (10 pM); lane 3, IL-5-treated eosinophils cocultured for 2 days with SB 203580 (10 μM).
If SB 203580 promotes apoptosis by inhibiting p38 MAP kinase, then logic dictates that this enzyme must be activated at the time eosinophils begin to apoptose. To test this hypothesis, Western blotting was performed using an antibody that recognizes the dual phosphorylated (activated) form of the enzyme. In freshly prepared, cytokine-deprived eosinophils, which underwent constitutive apoptosis rapidly, a low basal level of dual phosphorylated p38 MAP kinase was routinely detected. After 24 h of culture, at a time when ∼20% of the eosinophils were apoptotic, the phosphorylation of p38 MAP kinase was significantly increased by a mechanism that was not significantly altered by 10 pM IL-5 (Fig.6). p38 MAP kinase also was activated under resting conditions in cytokine-deprived, slow-dying eosinophils (Fig. 6). However, in contrast to their fast-dying counterparts, no significant increase in p38 MAP kinase phosphorylation was detected in this population after 24 h in the absence or presence of 10 pM IL-5 (Fig. 6), which was consistent with the presence of relatively few apoptotic cells at that time (see Fig. 1B).

Activation status of p38 MAP kinase in human eosinophils in culture. Fast- and slow-dying eosinophils were cultured in the absence or presence of IL-5 (10 pM) for 24 h and dual phosphorylated p38 MAP kinase was assessed by Western blotting. The results are representative of three experiments performed on eosinophils harvested from different donors.
It has been reported that certain isoforms of JNK, in particular the 46- and 54-kDa β splice variants of JNK-2 are inhibited by SB 203580 at concentrations in the low micromolar range (Whitmarsh et al., 1997;Clerk and Sugden, 1998). Given that JNK has been implicated in the regulation of cell longevity (see introduction), it was possible that the apoptotic effect of SB 203580 and SB 202190 was due the inhibition of a JNK-2. However, this was discounted, as neither the 46- nor the 54-kDa JNK-2 isoform were activated in cytokine-deprived human eosinophils or after exposure of cells to 10 pM IL-5 (data not shown).
To confirm that SB 203580-induced eosinophil death was not due to primary necrosis, the number of apoptotic, nonviable, and necrotic cells was determined (Fig. 7A). In eosinophils that died slowly, SB 203580 (10 μM) significantly increased the number of apoptotic cells at 24 h. This was followed at 48 h by an increase in the number of nonviable eosinophils but not necrotic cells, which is characteristic of programmed cell death. In fact, the number of necrotic cells remained low and constant for up to 5 days of culture with SB 203580, eliminating the possibility of toxicity (Fig. 7A). The induction of apoptosis and absence of necrotic cell death in slowly dying cells was also confirmed morphologically (Fig. 7, B and C). After 48-h culture, vehicle-treated eosinophils showed the typical morphology (multi-lobed nucleus, granular cytoplasm) of normal freshly isolated eosinophils (Fig. 7B). In contrast, cells cultured with SB 203580 (10 μM) exhibited the typical morphology of apoptotic cells such as cell shrinkage and nuclear chromatin condensation. Furthermore, the number of late apoptotic cells without visible nuclei or “ghosts” was markedly increased (Fig. 7C).

Effect of SB 203580 on the number of apoptotic, necrotic, and nonviable eosinophils. Cytokine-deprived, slow-responding eosinophils were cultured for 5 days in the presence of SB 203580 (10 μM) and the number of apoptotic (●), necrotic (▪), and nonviable (○) eosinophils was assessed by flow cytometric analysis of PI-stained DNA content, relative DNA content associated with membrane permeability, and PI exclusion respectively (A). Each data point represents the mean ± S.E.M. of four independent determinations using eosinophils from different donors. B and C, typical morphology of May-Grünwald-Giemsa-stained eosinophils that were cultured for 2 days in the absence and presence of SB 203580 (10 μM), respectively. Magnification: 400×. See Materials and Methods for further details.
Inhibitors of p38 MAP kinase have also been shown to suppress arachidonic acid metabolism in some cell types (Guan et al., 1997;Pouliot et al., 1997). To exclude the possibility that a prostanoid or leukotriene was responsible for enhancing eosinophil viability, the effect of cyclooxygenase and 5-lipoxygenase inhibitors on apoptosis was studied. Flurbiprofen and indomethacin, inhibitors of both cyclooxygenases 1 and 2 (Meade et al., 1993) did not affect human eosinophil apoptosis. L-745,337, a selective inhibitor of cyclooxygenase-2 (Chan et al., 1995) and Bay x1005, an inhibitor of 5-lipoxygenase (Fruchtmann et al., 1993) similarly were inactive (Table2).
Effect of inhibitors of cyclooxygenase and 5-lipoxygenase on human eosinophil apoptosis
Effect of PD 098059 on Eosinophil Longevity.
A pharmacological approach also was used to examine the role of MEK-1 in eosinophil apoptosis. Eosinophil viability was assessed in cultures supplemented with PD 098059, which inhibits ERK-1 and ERK-2 by binding to the inactive form of MEK-1, thereby preventing its phosphorylation and activation by Raf-1.
In contrast to the results obtained with the p38 MAP kinase inhibitors, PD 098059 (0.1–30 μM) did not significantly affect apoptosis in fast-or slow-dying eosinophils or the enhanced survival of cells cultured with IL-5 (10 pM; Fig. 8). The inability of PD 098059 to reverse the effect of IL-5 was confirmed morphologically. Thus the percentage of apoptotic eosinophils in IL-5 (10 pM)-treated cultures after 48 h was 6.2 ± 0.2, 8.3 ± 1.3 and 8.3 ± 1.9% (n = 3) in absence and in presence of 10 and 30 μM PD 098059, respectively.

Effect of PD 098059 on spontaneous apoptosis of eosinophils and on the survival-enhancing effect of IL-5. Fast- and slow-responding eosinophils were cultured for 2 days in medium containing PD 098059 (1–30 μM) in the absence (●) and presence of 10 pM (μm) IL-5, and apoptosis was assessed by flow cytometric analysis of PI-stained DNA content. Data points represent the mean ± S.E.M. of four independent determinations using eosinophils from different donors. See Materials and Methods for further details.
To establish that PD 098059 was active in the aforementioned studies, Western immunoblot analyses of cytosolic proteins extracted from human eosinophils were performed using a polyclonal antibody that recognizes dual phosphorylated forms of ERK-1 and ERK-2. As IL-5 has previously been reported to activate ERK-1 in eosinophils (Pazdrak et al., 1995a,b), experiments were performed initially to determine the concentration dependence of this effect and to assess whether ERK-2 was also activated. In untreated cells, no evidence for activity was detected at 30 min or 24 h after eosinophils were placed in culture, suggesting that these kinases are not central to the induction of spontaneous apoptosis (data not shown). In contrast, IL-5 (10 pM), after a lag of approximately 5 min, promoted the phosphorylation of ERK-1 and ERK-2 in human eosinophils in a time-dependent manner (Fig.9A). Maximal phosphorylation was achieved 30 min after IL-5; thereafter, the activity of ERK-1 and ERK-2 gradually declined such that at 2 h both kinases were dephosphorylated (Fig. 9A). Accordingly, in experiments where PD 098059 was used, measurements were made 30 min after exposure of eosinophils to 10 pM IL-5. Under these conditions, PD 098059 (10 μM) markedly suppressed (>90%) the appearance of phospho-ERK-1 and phospho-ERK-2 (Fig. 9B).

Effect of IL-5 on the dual phosphorylation status of ERK-1 and ERK-2 in human eosinophils and the effect of PD 098059. Eosinophils were cultured in the presence of IL-5 (10 pM) for 0 to 24 h and dual phosphorylated ERK-1 and ERK-2 was assessed by Western blotting (A). B, shows the effect of PD 098059 (10 μM) on the ability of IL-5 (10 pM) to activate ERK-1 and ERK-2 at 30 min. The results are representative of three experiments performed on eosinophils harvested from different donors.
Effect of Preincubation with IL-5 on Eosinophil Survival.
Previous studies have shown that ERK-1 and/or ERK-2 are activated by IL-5 in human eosinophil (Pazdrak et al., 1995a,b; Bates et al., 1996;Hiraguri et al., 1997; Coffer et al., 1998). To further investigate their potential role in IL-5-induced survival, we determined whether brief activation of these MAP kinases was sufficient to prevent apoptosis. Thus, eosinophils were preincubated for 15 to 180 min with 10 pM IL-5, washed, and resuspended in fresh medium containing no IL-5. Consistent with our previous experiments, the majority of eosinophils (95 ± 0.4%) incubated with IL-5 (10 pM) for the whole culture period (48 h) were not apoptotic (n = 3). In contrast, brief exposure of eosinophils to IL-5 (10 pM) for 15, 30, 60, or 180 min before culture inhibited apoptosis by 9 ± 3 (P > .05), 10 ± 7 (P > .05), 10 ± 1 (P > .05), and 20 ± 4% (P > .05), respectively, when compared with vehicle-treated, time-matched control samples. Thus, the eosinophils required continuous exposure to IL-5 for the survival-prolonging action to be maintained.
Discussion
Retrospective analysis of the time course of constitutive apoptosis of cytokine-deprived, human cultured eosinophils purified from the peripheral blood of 35 subjects identified two distinct populations: donors whose eosinophils underwent apoptosis quickly or relatively slowly (see Results for selection criterion). This distinction has not been reported previously although our finding that eosinophils from certain individuals undergo rapid apoptosis is consistent with that observed by the majority of investigators (Yamaguchi et al., 1991; Stern et al., 1992; Yousefi et al., 1994;Walsh, 1997). Significantly, this phenomenon was not dependent upon gender, disease, or atopic status, and the constitutive rate of apoptosis was reproducible within individuals over consecutive bleeds many weeks apart, indicating that the basis of the difference is intrinsic to each donor. Initially, it was reasoned that one explanation for this striking difference was due to the differential release of autocrine factors between donors that are either pro- or antiapoptotic. Although this is an attractive possibility, studies with neutralizing antibodies, receptor antagonists, and enzyme inhibitors excluded a role for IL-3, IL-4, IL-5, transforming growth factor-β1, tumor necrosis factor-α (TNF-α), CD40, eotaxin, platelet-activating factor, prostanoids, and leukotrienes B4, C4, and D4 (H.K., M.A.L., M.A.G., and P.J.B, unpublished observations). Subsequent studies, however, discovered that the survival of human eosinophils in culture is influenced by prior drug therapy. Thus, eosinophils purified from the blood of steroid-naı̈ve patients with asthma who take β2-adrenoceptor agonists on a regular basis, died apoptotically at a slower rate than eosinophils prepared identically from normal healthy individuals (H.K., M.A.L., M.A.G., and P.J.B., unpublished observations).
Biochemical and pharmacological evidence suggests that the activation of p38 MAP kinase can be both pro- (Graves et al., 1996; Sutherland et al., 1996; Gardner and Johnson, 1996; Kummer et al., 1997; Schwenger et al., 1997) and antiapoptotic (Sutherland et al., 1996; Gardner and Johnson, 1996; Nemoto et al., 1998). Studies were therefore performed to determine the extent to which p38 MAP kinase was involved in the constitutive apoptosis of human eosinophils and the survival-enhancing effect of IL-5. Using the complementary techniques of flow cytometry and morphology, together with an assessment of oligonucleosomal DNA fragmentation, it was found that SB 203580 and SB 202190, inhibitors of p38 MAP kinase, promoted apoptosis of cytokine-deprived eosinophils over a concentration range similar to that required to suppress a number of p38 MAP kinase-dependent responses including: 1) IL-6 generation from IL-1β-stimulated human fibroblast-like synoviocytes (Miyazawa et al., 1998); 2) IL-6 generation from TNF-α-stimulated murine fibrosarcoma L929 cells (Beyaert et al., 1996); 3) complement receptor 3-dependent adhesion and adhesion-dependent oxidative burst of human neutrophils in response to lipopolysaccharide and TNF-α (Detmers et al., 1998); and 4) the phosphorylation, in intact cells, of mitogen-activated protein kinase-activated protein kinase-2 (Miyazawa et al., 1998), a downstream substrate of p38 MAP kinase, and heat shock protein-27, a downstream substrate of mitogen-activated protein kinase-activated protein kinase-2 (Ridley et al., 1997). This pro-apoptotic effect was most prominent in slow-dying eosinophils but still was evident in cells that apoptosed quickly. The possibility that prostanoids and/or leukotrienes might mediate SB 203580-induced apoptosis (Guan et al., 1997; Pouliot et al., 1997) was excluded on the basis that inhibitors of 5-lipoxygenase and cyclooxygenases 1 and 2 were inactive alone and in SB 203580-treated eosinophils.
The mechanism of SB 203580- and SB 202190-induced apoptosis of human eosinophils is unclear. The finding that p38 MAP kinase was activated above basal levels in cytokine-deprived, fast-dying eosinophils at a time that coincided with the onset of apoptosis suggested that a causal relationship might exist between these two responses. However, two inhibitors of p38 MAP kinase, SB 203580 and SB 202190, enhanced apoptosis suggesting that activation of p38 MAP kinase may, paradoxically, represent an antiapoptotic signal in human eosinophils. Support for this proposal derives from identical studies performed in slow-dying cells in which SB 203580 and SB 202190 also promoted apoptosis at the same time point. However, in this population of cells the activation state of p38 MAP kinase was not increased, indicating that the basal activity of p38 MAP kinase is central to cell survival. Indeed, this conclusion is consistent with a recent paper by Aoshiba et al. (1999) in which the basal activity of p38 MAP kinase was shown to regulate longevity of human neutrophils, although in that cell SB 203580 inhibited spontaneous apoptosis. The above discussion notwithstanding, SB 203580 and SB 202190 may enhance constitutive eosinophil apoptosis through a p38 MAP kinase-independent mechanism.
Consistent with previous studies, we have demonstrated that eosinophils cultured in the absence of cytokines undergo apoptosis and that this process can be reversed by IL-5 (Yamaguchi et al., 1991; Tai et al., 1991; Stern et al., 1992). Moreover, IL-5 rescued eosinophils from apoptosis induced by SB 203580 and SB 202190. That effect was concentration-dependent but unidirectional in that the p38 MAP kinase inhibitors could not overcome the survival-enhancing activity of low concentrations of IL-5. Pharmacologically, these results suggest that IL-5 does not prolong eosinophil survival by activating p38 MAP kinase. This interpretation was supported by Western analysis of IL-5-treated, fast-dying eosinophils at 24 h in which no significant increase in dual phosphorylated p38 MAP kinase was seen when compared to cytokine-deprived cells at the same time point. A role of p38 MAP kinase in mediating the eosinophil survival-enhancing activity of IL-5 would have been surprising given that a src-related tyrosine kinase, lyn, is activated by IL-5 in eosinophils and is believed to regulate this response (Yousefi et al., 1996; Pazdrak et al., 1998). The downstream substrates of lyn are not unequivocally established but there is good evidence that the Ras-Raf-1-MEK-ERK pathway is activated (Pazdrak et al., 1995a, b; Bates et al., 1996; 1998; Coffer et al., 1998). Indeed, a reduction in the expression of lyn and Raf-1 in human eosinophils by the use of antisense deoxyoligonuceotides is reported to prevent IL-5 from prolonging survival (Pazdrak et al., 1998). Because MEK-1 is downstream from lyn and Raf-1 and that its only known substrates are ERK-1 and ERK-2, it seemed plausible that pharmacological inhibition of MEK activity should also promote apoptosis. However, contrary to expectation, the MEK-1 inhibitor, PD098059, did not affect eosinophil survival at concentrations that suppressed ERK-1/ERK-2 phosphorylation by >90%, which is entirely consistent with recent data found in human neutrophils (Aoshiba et al., 1999). Clearly, these results are contrary to the antisense experiments reported by Pazdrak et al. (1998) and are difficult to explain. Nevertheless, they tempt speculation that activation of the Jak2-STAT1 or Jak2-phosphatidylinositol 3-kinase pathways by IL-5 may be a primary determinant of eosinophil longevity (Hiraguri et al., 1997; Pazdrak et al., 1998). In this respect tyrphostin AG490, a selective inhibitor of Jak2 without effects onsrc-related tyrosine kinases, is reported to prevent IL-5-enhanced eosinophil survival (Pazdrak et al., 1998). The ability of IL-5 to activate ERK-1 and/or ERK-2 via MEK indicates that other, possibly more immediate, responses are regulated by this protein kinase cascade such as adherence and CD69 expression, priming of degranulation, chemotaxis, and the activation of the NADPH oxidase (seeGiembycz and Lindsay, 1999).
In conclusion, this study identified two distinct populations of individuals whose eosinophils, when placed in culture in the absence of cytokines, die apoptotically at significantly different rates. Moreover, SB 203580 and SB 202190 augmented constitutive apoptosis in both populations of eosinophil, suggesting that the activation state of p38 MAP kinase can have a profound effect on eosinophil longevity. Finally, despite indications to the contrary, our studies with PD 098059 suggest that MEK-1 is not involved in the processes that control spontaneous apoptosis or the enhanced survival of eosinophils effected by IL-5.
Footnotes
-
Send reprint requests to: Dr. Mark A. Lindsay, Thoracic Medicine, National Heart & Lung Institute, Imperial College of Science, Technology & Medicine, Dovehouse Street, London SW3 6LY, UK. E-mail:m.lindsay{at}i.c.ac.uk
-
↵1 This work was supported by the Academy of Finland, the Medical Research Council (MRC) (UK), British Lung Foundation (BLF) and the Wellcome Trust (UK) Grant 056814.
-
↵2 Medical School/B, University of Tampere, P.O. Box 607 FIN-33101, Tampere, Finland.
- Abbreviations:
- IL
- interleukin
- ERK
- extracellular-regulated kinase
- MAP
- mitogen-activated protein
- MEK
- mitogen-activated protein kinase kinase
- JNK
- c-jun N-terminal kinase
- TNF-α
- tumor necrosis factor-α
- FCS
- fetal calf serum
- PI
- propidium iodide
- TBS-T
- 25 mM Tris base, 150 mM NaCl, 0.1% Tween 20, pH 7.4
- ECL
- enhanced chemiluminescence
- Received December 17, 1998.
- Accepted April 21, 1999.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}