


Abstract
In vitro, (S)-2-(chloro-5-fluoro-indol-1-yl)-1-methylethylamine 1:1 C4H4O4 and (S)-2-(4,4,7-trimethyl-1,4-dihydro-indeno[1,2-b]pyrrol-1-yl)-1-methylethylamine 1:1 C4H4O4 exhibited high-affinity binding to the serotonin2C (5HT2C) receptors and stimulated turnover of inositol 1,4,5-triphosphate. Affinity to several of the other 5-HT receptor subtypes and to numerous nonserotonergic receptors was much lower. In rats, both compounds elicited behavioral signs of 5-HT2C receptor agonism but not 5-HT2A receptor agonism. Hypomotility induced in rats by high doses of these compounds was reversed by the 5-HT2Creceptor antagonist N-(2-naphthyl)-N′-(3-pyridyl)-urea 1:1 HCI. In addition, these compounds were active in tests used to demonstrate anticompulsive effects: reducing schedule-induced polydipsia in rats (prevented by the 5-HT2C/2B receptor antagonist N-(1-methyl-5′-indolyl)-(3-pyridyl)urea 1:1 HCl, reversing increased scratching induced with 8-hydroxy-dipropylaminotetralin 1:1 HCl in squirrel monkeys (no tolerance developed), decreasing responding in the marble-burying task in mice, and decreasing excessive eating of palatable food in rats. In contrast to these compounds, fluoxetine was much less potent, and in some tasks less efficacious, in reducing excessive behavior in these models. These two 5-HT2Creceptor agonists do not show anxiogenic effects in the plus-maze in rats. (S)-2-(4,4,7-trimethyl-1,4-dihydro-indeno[1,2-b]pyrrol-1-yl)-1-methylethylamine 1:1 C4H4O4 reduced the olfactory bulbectomy-induced passive avoidance impairment in rats, a result that indicates antidepressant potential. Similarly, in the differential-reinforcement-of-low rate 72-s operant schedule task in rats, (S)-2-(chloro-5-fluoro-indol-1-yl)-1-methylethylamine 1:1 C4H4O4 increased (and (S)-2-(4,4,7-trimethyl-1,4-dihydro-indeno[1,2-b]pyrrol-1-yl)-1-methylethylamine 1:1 C4H4O4 showed a tendency to increase) total reinforcements received, which is suggestive of antidepressant activity. The electroencephalography defined sleep-waking pattern in rats produced by these two 5-HT2Cagonists, as well as fluoxetine, included increased quiet-waking and decreased rapid-eye-movement sleep, which is characteristic of antidepressant drugs. These results suggest that 5-HT2Creceptor agonism is associated with therapeutic potential in obsessive compulsive disorder and depression.
SSRIs are now well established as efficacious treatment for depression, OCD and bulimia (Den Boer and Westenberg, 1995; Kennedy and Goldbloom, 1994; Rasmussen et al., 1993). Various pharmacological effects of the neurotransmitter 5-HT have been demonstrated to depend predominantly on its interaction with specific 5-HT receptor subtypes (cf. Saxena, 1995). This, in turn, has led to the development of such drugs as buspirone, a 5-HT1A receptor partial agonist, for treatment of anxiety disorders (Goa and Ward, 1986) and ondansetron, a 5-HT3 receptor antagonist, for reduction of emesis (Milne and Heel, 1991).
5-HT2C receptors are present at very high levels in choroid plexus (as the only 5-HT receptor subtype located there), but they also occur in various other brain regions (Pompeiano et al., 1994). 5-HT2C receptor mRNA is found in many brain areas in addition to those autoradiographically shown to have binding sites. 5-HT2C mRNA was located in noradrenergic, dopaminergic and cholinergic nuclei. Lower, but nonetheless still high, receptor densities are found in the limbic system and cortex (especially frontal cortex), a result consistent with a possible important role of 5-HT2C receptors in affective disorders. Specific antibodies have also been used to study the location of 5-HT2C receptors, recognizing sites in rat choroid plexus, hippocampus, cerebral cortex, striatum and substantia nigra and in human cerebral cortex, substantia nigra and cerebellum (Abramowski et al., 1995).
The potential use of 5-HT2C receptor ligands in psychiatry is suggested by a number of observations. Transgenic mice that lacked functional 5-HT2C receptors exhibited abnormal feeding behavior leading to overweight, as well as susceptibility to seizures (Tecott et al., 1995). 5-HT2C receptor-mediated effects, such as hypophagia, resemble those induced by SSRIs (Leander, 1987; Lucki et al., 1988), and furthermore, the interoceptive cue produced by SSRIs has been reported to exhibit similarity to that of the 5-HT2C receptor agonist MK212, only partially to the 5-HT1A/7 receptor agonist 8-OH-DPAT and not to the 5-HT2A receptor agonist DOI (Berendsen and Broekkamp, 1994). Such results suggest that some of the therapeutic effects of the SSRIs may be mediated at least in part by 5-HT2C receptor agonism. Certainly there are a number of common effects in rodents that have now been noted for SSRIs and 5-HT2C receptor agonists, including inhibition of escape from aversive periaqueductal-gray stimulation, hypophagia, decreased defensive burying, induction of penile erection and inhibition of muricide (Broekkamp and Berendsen, 1992). A considerable body of preclinical and clinical data on m-CPP, which exhibit high-affinity binding to 5-HT2C receptors (as well as affinity to several other 5-HT receptor subtypes and nonserotonergic receptors) indicates that under some circumstances it exacerbates clinical symptoms in patients with OCD or panic anxiety (Kahn and Wetzler, 1991). However, recent results suggest that such effects are probably 5-HT1D receptor-mediated (Zohar and Cohen, 1995; Loiet al., 1995). Unfortunately, only nonselective 5-HT2C receptor agonists have been available as research tools. The present study provides an extensive biochemical and pharmacological characterization of two novel 5-HT2Creceptor agonists.
Novel agents acting at the central 5-HT2C receptors may offer therapeutic advantages. The absence of appreciable affinity to certain other 5-HT subtypes may provide the basis for an improved therapeutic index compared with the general activation of all the different 5-HT receptor subtypes by SSRIs. For example, 5-HT1A receptor agonism has been associated with evidence of sexual dysfunction and anxiogenesis in animals (e.g.,Ahlenius et al., 1986; Moser et al., 1990;Maswood et al., 1996), and 5-HT2A receptor agonism has been implicated in cardiovascular effects (Saxena, 1995). The experimental results described here with novel selective 5-HT2C receptor agonists in different animal models of psychiatric disorders support this view. Ro 60-0175/ORG 35030 and Ro 60-0332/ORG 35035 are 5-HT2C receptor agonists that offer the potential advantages of therapeutically relevant effects in animal models of OCD and depression similar to or better than those of fluoxetine, combined with an improved side-effect profile. SSRIs enhance endogenously released 5-HT and thereby activate all postsynaptic 5-HT receptor subtypes. A direct agonistic effect at the 5-HT2C receptor (especially in the absence of any appreciable 5-HT2A receptor agonism) defines the probable mechanism responsible for the favorable pharmacological profiles of Ro 60-0175/ORG 35030 and Ro 60-0332/ORG 35035.
Materials and Methods
In Vitro Pharmacology
In vitro binding assays.
In most binding assays, the compounds were tested at a single concentration (10−5M) using the methodology described in table1. In those assays where the compounds displaced more than 50% of the specific binding, the compounds were retested at multiple concentrations to estimate pKi values. With respect to the 5-HT1A, 5-HT2A and 5-HT2C receptors, experiments were performed with membranes obtained from NIH 3T3 cell lines expressing human 5-HT1A, human 5-HT2A or human 5-HT2C receptors (kindly donated by Dr. Nico Stam, N.V. Organon). For each receptor subtype, a single batch of cells was grown using fermentation techniques previously described (Sleight et al., 1996). Radioligand binding assays were as previously described for the human 5-HT2A receptor, with minor modifications for labeling human 5-HT1A and human 5-HT2C receptors. Briefly, on the day of the experiment, membranes were thawed and resuspended in 10 times the original volume of assay buffer to give a concentration of approximately 4 × 105 cells per assay tube. The assay buffer consisted of Tris-HCl 50 mM, pargyline 10−5 M, MgCl2 5 mM and ascorbic acid 0.1%, pH 7.4. All compounds were dissolved in 10% dimethyl sulfoxide and diluted in assay buffer. Assays were similar for each receptor and consisted of 100 μl of membrane preparation (depending on the assay), 50 μl of radioligand ([3H]-5-HT 1 nM final concentration for labeling human 5-HT1A and human 5-HT2C receptor binding sites and [3H]-DOB 1 nM final concentration for labeling human 5-HT2Areceptors). Nonspecific binding was defined in the presence of 10 μM 5-HT in the case of the human 5-HT1A and 5-HT2Creceptor and in the presence of 10 μM methysergide in the case of the human 5-HT2A receptor. The specific activities of [3H]-5-HT and [3H]-DOB were 29.7 and 15.0 Ci/mmole, respectively (New England Nuclear, Boston, MA). The incubations were performed at room temperature for 1 h. For all binding experiments, reactions were stopped by rapid filtration through either Whatmann GF/B or GF/C filters. Filters were washed with 3 × 2 ml of Tris-HCl (50 mM, pH 7.4), and the radioactivity retained on the filters was measured by scintillation spectroscopy. All displacement experiments were performed in triplicate and were repeated at least three times. Saturation analyses were performed for each receptor using at least eight concentrations of each radioligand (concentrations ranging from 0.05 to 10 nM). Dissociation constants (Kd) were calculated using the EBDA/LIGAND program (McPherson, 1985; Munson and Rodbard, 1980). Where displacement experiments were performed, curves were constructed using seven concentrations of the displacing agents (one data point per logarithmic unit of concentration: 10−11 to 10−5 M). Displacement curves were analyzed using EBDA/LIGAND to calculate pKi values.
Assay conditions for radioligand binding experiments
Tissue preparation and incubation for measurement of IP3 production.
5-HT2C-mediated stimulation of IP3 production was measured in the choroid plexus of the rat. The choroid plexus was removed, placed in 200 μl of oxygenated Krebs solution and incubated with 0.35 nmol myoinositol and 0.35 nmol [3H]-myoinositol for 1 h at 37°C. During this incubation, the tubes were gassed with 95% oxygen/5% CO2 every 20 min. A mixture of LiCl and pargyline was then added (final concentration: LiCl = 10 mM, pargyline = 10 μM), followed 10 min later by addition of the test compounds (final incubation volume = 250 μl). Dose-response curves were constructed from data obtained from three separate measurements per data point. The mixture was incubated for a further 30 min at 37°C. The assays were stopped by the addition of 25 μl of a stopping solution (HClO4 2.64 N + EDTA 40 mM). Assay tubes were frozen on dry ice for 15 min, thawed and then kept on ice for 1 h. The tubes were then centrifuged for 20 min at 24,000 × g. Then 250 μl of the supernatant was removed and placed in Eppendorf tubes together with 25 μl 4 M KOH. The samples were mixed well and kept on ice for 15 min. These samples were then recentrifuged for 15 min at 14,000 rpm, 230 μl of supernatant was removed and 30 μl of phytic acid was added. The isolation of IP3 was as described in a previous report (Bourson et al., 1996). A concentration-response curve was constructed for 5-HT, m-CPP and the test compounds. Six concentrations were used per test compound, the highest concentration tested being 0.1 mM. The maximal effect produced by each compound was compared with the stimulation induced by 10 μM 5-HT to calculate the relative intrinsic activity.
In Vivo Pharmacology
Animals and maintenance conditions.
Adult male and female MORO mice and 3-week-old female DBA/2J mice (Biological Research Laboratories, CH-4414 Füllinsdorf, Switzerland) and male Swiss mice (Charles River, Sulzfeld, Germany) were used. Adult male and female RORO rats (Biological Research Laboratories, CH-4414 Füllinsdorf, Switzerland), male and female Sprague-Dawley rats (Biological Research Laboratories, CH-4414 Füllinsdorf, Switzerland and Charles River, Sulzfeld, Germany) and male Long-Evans rats (Hsd/Cpbs: Harlan; Zeist, The Netherlands) were used. The mice and rats were delivered to the laboratory colony at least one day before testing. These animals were housed in group cages with sawdust bedding under standard maintenance conditions (12:12 h light-dark cycle; 21–23°C; 55–65% relative humidity). Both mice and rats received laboratory chow and tap water ad libitum in the home cage (except when otherwise specified by the experimental procedure). All testing was done during the light portion of the day-night cycle. At the conclusion of testing, the rodents were euthanized by CO2 exposure.
Adult male squirrel monkeys (Saimiri sciureus) of approximately 1 kg b.w.t. were used. The monkeys were maintained in an isolated facility under the supervision of veterinary staff. All monkeys had been in the laboratory colony for several years and were drug-experienced. Before the start of any of the present experiments, the monkeys had received no drug treatment for at least 1 month. The monkeys were maintained either in groups of 6 to 12 in a room-size volière or in pairs in stainless steel cages (0.85 m × 0.6 m × 1.2 m) with two elevated platforms and a hanging chain for climbing. Both the temperature (28–30°C) and the humidity (50–60%) of the animal quarters were regulated. A 12:12 hour light-dark cycle with light onset at 6 a.m. was used. A dry, nutritionally sufficient diet was available ad libitumand was supplemented each day with fresh fruits and vegetables. Tap water was available continuously in the home cage.
Behavioral observation in rats.
Adult male RORO rats were injected s.c. with vehicle, Ro 60-0175/ORG 35030 or Ro 60-0332/ORG 35035, as well as the reference compounds (±)-DOI and m-CPP and then placed individually in Macrolon cages (29 × 24 × 9 cm) with sawdust bedding for observation of the intrinsic effects of each test compound. Groups of up to eight rats representing different treatment conditions were observed simultaneously. A mirror placed behind the cages permitted an all-round view of the rats. The experimental compounds were given in 0.3% (w/v) Tween-80 in distilled water in a volume of 2 ml/kg b.wt. The doses 0.1, 0.32, 1, 3.2 and 10 mg/kg were evaluated (m-CPP was also tested at the doses 0.01 and 0.032 mg/kg). The rats were drug-experienced (at least 3 days washout between successive tests). Observation began 5 min after injection. The presence or absence of penile erection (see Berendsen and Gower, 1986) was scored during successive 5-min intervals for 40 min. Head shakes (total number of separate bouts) were scored during four 5-min blocks at 5, 15, 25 and 35 min after injection and cumulated for the whole session. These behavioral signs provide a preliminary in vivo indication of 5-HT2C receptor (i.e.,penile erection) and 5-HT2A receptor (i.e., head shake) activation (Berendsen et al., 1990). The ED50 value was calculated by probit analysis.
Behavioral observation in squirrel monkeys.
Adult male squirrel monkeys were administered p.o. different doses of fluoxetine (1, 3, 10 or 30 mg/kg), Ro 60-0175/ORG 35030 (0.3, 0.6, 1, 3 or 10 mg/kg), Ro 60-0332/ORG 35035 (0.3, 1, 3 or 10 mg/kg), or vehicle and then placed in an individual stainless steel cage (0.85 m × 0.6 m × 1.2 m) with two elevated platforms and a hanging chain for climbing. Groups of 5 to 10 monkeys per treatment condition were used. The test compounds were given in 0.3% (w/v) Tween-80 in distilled water in a volume of 2 ml/kg b.wt. Behavioral observations of the squirrel monkeys by a trained observer began immediately after administration and continued over a period of at least 6 h.
Spontaneous motor activity in rats.
Locomotor activity was monitored in naive adult male RORO rats via a Digiscan Animal Activity Monitoring System (Omnitech Electronics, Columbus, Ohio). The test compounds were administered in 0.3% (w/v) Tween-80 in distilled water in a volume of 2 ml/kg b.wt. All testing was carried out in an illuminated room during the light portion of the day-night cycle. The parameter total distance (centimeters) was measured. The 5-HT2C receptor antagonist Ro 60-0491 (pKivalues: h5-HT2C = 7.7; h5-HT2A = 6.0, reversal of m-CPP induced penile erection: ID50 = 7.7 mg/kg s.c.; unpublished results) at the dose 10 mg/kg or vehicle was injected i.p., followed 15 min later by a second i.p. injection of vehicle, m-CPP (6 mg/kg), Ro 60-0175/ORG 35030 (10 mg/kg) or Ro 60-0332/ORG 35035 (10 mg/kg). Immediately after the second injection, each rat was placed into an individual Plexiglas test cage (41 × 41 × 28 cm) with sawdust bedding, and its activity was measured for a 30-min period. A different group of eight rats was used for each treatment condition. Statistical comparison was done with a two-tailed Mann-Whitney U test with a P value ≤ .05 accepted as significant.
Anticonvulsant effects in mice.
Protection from audiogenic seizures was investigated in naive female 21-day-old DBA/2J mice (a strain that exhibits age-dependent susceptibility to seizure). Oral treatment with a test compound or vehicle was given 30 min before testing. Ro 60-0175/ORG 35030 (21, 32, 46, 68, 100 or 150 mg/kg) and Ro 60-0332/ORG 35035 (3.2, 10, 32 or 100 mg/kg) were given in 0.3% (w/v) Tween-80 in distilled water in a volume of 10 ml/kg b.wt. After treatment, each mouse was placed in a separate open transparent Plexiglas box composed of half of a circular container (diameter = 40 cm, height = 20 cm) containing sawdust bedding. Testing was done in a sound-isolated chamber and involved exposure to a 14-kHz sinusoidal tone at 95 dB measured 2 cm above the floor (zero dB was defined as a pressure level of 20 μPa) for 1 min, during which observations were made. After vehicle treatment, such acoustic stimulation typically induced wild running, clonic seizures and tonic convulsions in approximately 90% of the mice tested. Groups of 6 to 8 mice were used to evaluate each dosage condition and the vehicle condition, and the proportion of the group that failed to exhibit tonic convulsions was recorded. ED50 values were calculated by probit analysis.
Elevated plus-maze task in rats.
Exploration was measured in an elevated plus-maze task in adult naive Sprague-Dawley male rats weighing 110 to 200 g at the time of testing. Drugs that increase exploration within this situation (i.e., benzodiazepine anxiolytics) are considered to exhibit anxiolytic-like effects. The apparatus was 50 cm above the floor and consisted of two open arms (50 cm × 10 cm) perpendicular to two closed arms (50 cm × 10 cm × 50 cm high) extending from an open central area (10 cm × 10 cm). The light intensity on the central platform was 225 lux. All parts of the apparatus were made of grey polyvinylchloride plastic. The effects of i.p. administration of fluoxetine (5 mg/kg), Ro 60-0175/ORG 35030 (1.5 mg/kg) and Ro 60-0332/ORG 35035 (7.5 mg/kg) were evaluated. The test compounds were given in 0.3% (w/v) Tween-80 in distilled water in a volume of 5 ml/kg b.wt. Thirty minutes after injection, the rat was placed in the center of the plus-maze facing one of the closed arms and observed for 5 min via a closed-circuit television camera by an observer located in an adjacent room. Number of entries and total time spent in the open arms were measured. The number of closed-arm entries was also recorded as a parameter of general locomotion, and the number of attempts at entry into open arms followed by avoidance behavior was recorded as a measure of risk assessment. Entry into an arm was defined as the rat placing all four paws into the arm. The floor of the maze was thoroughly cleaned after each test trial. The effects of both the first injection and the final injection in a series of four successive daily injections were investigated. During the subchronic experiment, the rats were group-housed under the previously described maintenance conditions with five rats per cage. Each treatment group was composed of 16 rats. Differences between vehicle and drug treatments were evaluated with a single-factor ANOVA followed by a Bonferroni/Dunn post-hoc test for multiple comparisons with a P value ≤ .05 accepted as statistically significant.
Burying behavior in mice.
In this assessment, the mice are individually placed in a cage with glass marbles located on a layer of sawdust bedding; the marbles make it easy to quantify burying behavior. Naive adult Swiss mice were tested in a Macrolon-type II test cage (24 × 18 × 13 cm) containing 25 marbles (1.5 cm in diameter) placed together in the middle on a 5-cm layer of sawdust. Fluoxetine (10, 22 or 46 mg/kg), Ro 60-0175/ORG 35030 (2.2, 4.6 or 10 mg/kg) and Ro 60-0332/ORG 35035 (4.6, 10 or 22 mg/kg) were evaluated. Vehicle or different doses of each experimental compound were administered s.c. 30 min before testing. The experimental compounds were given in 5% Mulgofen (EL 719R, GAG Corp., New York, NY) in physiological saline. The volume of administration was 10 ml/kg b.wt. Groups of nine mice were used to evaluate each treatment condition. The mice were placed in individual test cages for a 30-min test. The number of marbles covered at least two-thirds by sawdust was counted at the conclusion of testing. The median for each dosage group was calculated. The ED50 values were calculated by probit analysis.
Palatable food intake in rats.
Adult female RORO rats were used to evaluate hypophagia in a palatable food paradigm. The nondeprived rats were repeatedly given boiled potato in separate Macrolon test cages (30 × 25 × 10 cm) lacking bedding until a stable level of consumption was obtained during the 30-min test on several successive days. Subsequently, the same group of drug-experienced rats were tested with vehicle and with all dosage conditions for a given test compound, a minimum of 2 days intervening between successive tests. The compounds tested were fluoxetine (3, 10, 30 or 60 mg/kg), m-CPP (0.1, 0.3, 1 or 3 mg/kg), Ro 60-0175/ORG 35030 (0.3, 1, 3 or 10 mg/kg) and Ro 60-0332/ORG 35035 (10, 30 or 60 mg/kg). Several test sessions done with vehicle were interspersed with those of the different doses of a test compound (a mean value was then calculated). These test compounds were given in 0.3% (w/v) Tween-80 in distilled water in a volume of 2 ml/kg b.wt. Groups of 20 to 30 animals were used. The treatment was administered p.o. 30 min before testing in individual cages. Palatable food intake was determined (weighed to the nearest 1 g) for a 30-min test session. Such test sessions were alternated with training sessions in which no treatment was administered but otherwise the test was carried out in the same manner as previously described. Evaluation of the results was first done with the Friedman two-way analysis of variance to demonstrate overall statistical significance. Whenever overall statistical significance was found, subsequent analysis was carried out with a two-tailed Wilcoxon test to compare the effect of each individual dosage condition for a test compound with that obtained with vehicle. A P value of ≤ .05 was accepted as statistically significant. The minimal effective dose (MED) to produce a statistically significant reduction in palatable food intake was determined.
Schedule-induced polydipsia task in rats.
Excessive drinking was induced in adult female RORO rats through the use of a fixed-time operant schedule (FT-1 min). The rats were drug-experienced and were food-deprived overnight before each test session. The test apparatus consisted of a sound-attenuated chamber surrounding a Plexiglas test box (30 × 25 × 30 cm) that was equipped with a stainless steel grid floor and a mechanism to permit the automatic delivery of one 45-mg food pellet (Formula A/I; P.J. Noyes Company, Inc., Lancaster, NH) each minute into a food cup. The test session was 1 h. The test compounds were given in 0.3% (w/v) Tween-80 in distilled water in a volume of 2 ml/kg b.wt. In an initial experiment, either vehicle or a test compound was administered p.o. 30 min before the start of testing. The reference compound fluoxetine was tested at the doses 10, 30 and 60 mg/kg. Ro 60-0175/ORG 35030 and Ro 60-0332/ORG 35035 were tested at the doses 1, 3, and 10 mg/kg. A single group of 16 to 20 rats was used in each experiment to test vehicle and all of the selected doses of a given compound. Test days alternated with training days, on which the session proceeded in the same manner as on test days except that no treatment was given. In addition, we interspersed control sessions in which 60 food pellets were placed in the food cup at the start of the session and there was no other delivery of food during the entire 1-h test. A bottle containing tap water was always available on the test apparatus, but intake was measured (to the nearest 1 g) only for the test sessions and the control sessions. Evaluation of the results obtained in this test paradigm for both schedule-induced and control water intake was first done with the Friedman two-way analysis of variance to demonstrate overall statistical significance. Whenever overall statistical significance was found, subsequent analysis was carried out with a two-tailed Wilcoxon test to compare the effect of each individual dosage condition for a test compound to that obtained with vehicle. A P value of ≤ .05 was accepted as statistically significant.
In a subsequent set of experiments using this polydipsia paradigm, an initial i.p. dose of either vehicle or the selective 5-HT2C/2B receptor antagonist SB 200646A at the dose 30 mg/kg (Kennett et al., 1994) was administered 30 min before the test, followed 10 min later by p.o. administration of vehicle or the test compound (10 mg/kg Ro 60-0175/ORG 35030 or 20 mg/kg Ro 60-0332/ORG 35035). In a preliminary test it was shown that this dose of SB 200646A did not appreciably reduce stress-induced polydipsia. A single group of 14 to 16 rats was used in an experiment to evaluate each of these two test compounds. Testing then proceeded as previously described. Evaluation of the results obtained in this test paradigm was done using a two-tailed Wilcoxon test, a P value of ≤ .05 being accepted as statistically significant.
Compulsive behavior in monkeys.
Adult male squirrel monkeys first received p.o. administration of vehicle, fluoxetine (1, 3, 10 or 30 mg/kg), Ro 60-0175/ORG 35030 (0.3, 0.6, 1, 3 or 10 mg/kg) or Ro 60-0332/ORG 35035 (0.3, 1, 3 or 10 mg/kg). This dose of 8-OH-DPAT (0.1 mg/kg s.c.) was used because it was the lowest dose consistently to induce compulsive whole-body scratching (“displacement behavior”) occurring periodically in bouts in squirrel monkeys (Moreau et al., 1992). The test compounds were given in 0.3% (w/v) Tween-80 in distilled water in a volume of 2 ml/kg b.wt. Groups of 4 to 5 monkeys per dose were used, and 14 monkeys were given vehicle treatment. Each monkey was tested only once in this entire experiment. Immediately after injection of 8-OH-DPAT, the monkeys were placed in individual test cages, and scratching bouts were scored over the 2-h observation period after injection. The observer was not blind with respect to the treatment condition. The ID50 values were calculated with probit analysis.
In a subsequent experiment, different groups of 10 monkeys received p.o. administration of vehicle, 1 mg/kg Ro 60-0175/ORG 35030 or 10 mg/kg fluoxetine once daily for 15 days. A challenge test with 8-OH-DPAT was done as described above on the first and the final treatment day to permit an assessment of tolerance development. Evaluation of the results was done using a two-tailed Mann-WhitneyU test to compare treatment conditions and a two-tailed Wilcoxon test to compare the anticompulsive effect of each treatment condition on the first vs. the final treatment day. A P value of ≤ .05 was accepted as statistically significant.
Passive avoidance deficit in olfactory-bulbectomized rats.
Bulbectomized rats exhibit disturbed emotional behavior, which can be seen in impaired passive avoidance acquisition that is ameliorated by antidepressant treatment (Broekkamp et al., 1980). Naive Sprague-Dawley rats underwent bilateral bulbectomy via a vacuum suction method under pentobarbital anesthesia (60 mg/kg i.p. Nembutal, supplemented as needed). During convalescence the operated rats were housed in groups of five in Macrolon®-type II cages (24 × 18 × 13 cm). Passive avoidance training began 10 days after surgery. The apparatus was a black plastic box (40 × 40 × 40 cm) with a transparent plastic lid and a stainless-steel grid floor. A plastic platform (40 × 8 cm) was attached to a wall 4 cm above the grid floor. Vehicle (physiological saline) or 5 mg/kg Ro 60-0332/ORG 35035 was injected s.c. 30 min before the start of testing. The volume administered was 5 ml/kg b.wt. A trial began by placing a rat on the elevated platform above an electrified grid (0.7-mA scrambled shock). When the rat stepped down onto the grid with all four paws, foot shock was delivered for 2 s. The stepdown latency for the first trial was recorded. After each trial the rat was returned to its home cage for a period of 1 min. Training continued until the rat stayed on the elevated platform for 5 min or for a maximum of 10 trials. Groups of 8 to 10 rats per treatment condition were used. The total trials required to meet the learning criterion was evaluated using a two-tailed Mann-Whitney U test to compare groups, a P value ≤ .05 being accepted as significant. After the completion of testing, the rats were sacrificed and the olfactory bulb ablations verified.
DRL-72 s task in rats.
In a task employing the DRL-72 s, antidepressants have generally been found to dose-dependently decrease response rates but to increase the total number of reinforcements obtained in a session (O’Donnell and Seiden, 1983). Adult male Long-Evans rats (325–425 g) were individually housed and maintained on a schedule of restricted feeding (15–20 g per day) with water available ad libitum. Testing took place in standard operant chambers (Med Associates, Inc., Georgia, VT) equipped with a houselight, two cue lamps placed directly over two levers and a centrally placed pellet dispenser with cue light. The testing schedule was controlled by a DIG 700 series interface connected to an IBM P/S 2 PC running MedState notation software. The daily test sessions lasted 1 h. In the DRL-72 s task, the rat must wait a minimum of 72 s between lever presses in order to obtain food reinforcement; an earlier response resets the clock and is not reinforced. Only the right-hand lever was active in this procedure. Rats underwent an intensive training schedule until stable responding was obtained (Andrews et al., 1994), and evaluation of the test compounds followed. All rats were well trained and drug-experienced at the start of testing; difference groups of performance-matched rats were used to evaluate the different treatment conditions (N = 10–12). Vehicle (physiological saline/mulgofen), Ro 60-0175/ORG 35030 (1, 3 or 10 mg/kg), or Ro 60-0332/ORG 35035 (1, 3 or 10 mg/kg) was injected i.p. 30 min before the start of testing. The volume administered was 1 ml/kg b.wt. The total responses and total reinforcements per session were recorded. Data were analyzed with a one-factor ANOVA for independent groups, and post-hoccomparisons were planned with the Tukey multiple comparisons test using the Statistica software package (StatSoft Inc.), a P value < .01 being accepted as significant.
Sleep-wake behavior in rats.
Sleep-wake stages were evaluated in rats using an automated system that records and analyzes bioelectrical signals arising from parieto-occipital EEG, nuchal EMG and a movement detector. Male Hsd/Cpb rats were anesthetized (40 mg/kg i.p. Nembutal, supplemented as needed) and stereotaxically implanted with two epidural screw electrodes over the right parieto-occipital cortex with an epidural grounding screw electrode implanted over the frontal cortex. Two Teflon-coated stainless steel wire electrode were implanted bilaterally into the dorsal neck musculature and connected to screws fastened to the skull. After convalescence for at least 2 weeks, testing was done in a sound-attenuated box (100 × 40 × 200 cm) with the electrodes connected via a swivel to the data acquisition system. Gross movements were measured as capacitative artifacts generated in an open-ended wire in the flat cable between rat and swivel joint. Six sleep-wake stages are distinguished: 1) active waking characterized by movement, theta activity and high EMG, 2) quiet waking without movement, 3) quiet sleep, characterized by EEG spindles, 4) deep slow-wave sleep with prominent deltaactivity, 5) pre-REM sleep with spindles against a background oftheta activity and low EMG and 6) REM sleep withtheta activity and low EMG. The experimental compounds were given in physiological saline in a volume of 5 ml/kg b.wt. Treatment was given immediately before the start of testing. Fluoxetine was tested at the doses 1, 2.2, 3.2, 4.6, 10 and 32 mg/kg i.p. Ro 60-0175/ORG 35030 was tested at the doses 2.2 mg/kg i.p. and 3.2, 10 and 22 mg/kg p.o. Ro 60-0332/ORG 35035 was tested at the doses 1, 3.2 and 10 mg/kg i.p. and 3.2, 10 and 32 mg/kg p.o. Treatment was done at the start of the light cycle. Data for the initial 3 h after administration were evaluated. To reduce the variance in the analysis, percentage changes with respect to median values in the vehicle group were used. Statistical analysis was done with Student’s ttests with a significance level set at P < .05. A washout period of 2 to 3 weeks between successive tests was allowed. Groups of 6 to 9 rats were used for each treatment condition. Additional methodological details are provided elsewhere (Ruigt et al., 1989a,b).
Drugs
Ro 60-0175/ORG 35030 and Ro 60-0332/ORG 35035 were synthesized at F. Hoffmann-La Roche Ltd. (Basel, Switzerland) and are shown in figure 1. Fluoxetine HCl was purchased from Sigma Chemie (Buchs, St. Gallen, Switzerland). (±)-DOI HCl and m-CPP dihydrochloride were purchased from Research Biochemicals International (Natick, MA). SB200646A was synthesized at N.V. Organon. Ro 60-0491/001 and 8-OH-DPAT hydrochloride were synthesized at F. Hoffmann-La Roche Ltd. The conditions of administration are provided in the test descriptions. All doses were calculated for the salt.

Structures of Ro 60-0175/ORG 35030 and Ro 60-0332/ORG 35035.
Results
In Vitro Pharmacology
Receptor binding assays.
The in vitro binding results for 5-HT receptors are shown for Ro 60-0175/ORG 35030 and Ro 60-0332/ORG 35035 in table 2. Ro 60-0175/ORG 35030 exhibits high-affinity binding to the human 5-HT2C receptor (pKi = 9.0) with affinity for several other 5-HT receptor subtypes (1A, 1B, 1D, 3, 4, 6, 7) found to be at least 3 logarithmic units lower; affinity to the human 5-HT2A receptor was intermediate (pKi = 7.5). Ro 60-0332/ORG 35035 exhibits high-affinity binding to human 5-HT2C receptor (pKi = 8.5) with affinity for several other 5-HT receptor subtypes (1A, 3, 4, 6, 7) found to be at least 2 logarithmic units lower; affinity to the human 5-HT1Dα receptor (pKi = 7.4) and the human 5-HT2A receptor (pKi = 7.0) was intermediate. To assess the binding specificity, we performed a broad evaluation of an additional 26 receptors. IC50 values for Ro 60-0175/ORG 35030 were found to be >1 μM for all of the non-5-HT receptors listed in table 1 except the human beta-2 adrenoceptor (pIC50 = 6.6). IC50 values for Ro 60-0332/ORG 35035 were found to be >1 μM for all the non-5-HT receptors listed in table 1 except for rat dopamine D3 receptor and the human muscarinic M4 receptor, for which the approximate IC50 values were only slightly less than 1 μM.
Binding affinity (pKi) for 5-HT receptors
Stimulation of IP3 formation.
Both Ro 60-0175/ORG 35030 (pEC50 = 6.7, α = 1.1) and Ro 60-0332/ORG 35035 (pEC50 = 6.7, α = 1.0) exhibited a concentration-related increase in IP3 formation in rat choroid plexus that achieved a maximal effect similar to that of 5-HT itself.
Behavioral observation in rats.
Ro 60-0175/ORG 35030 (ED50 = 0.2 mg/kg s.c.), Ro 60-0332/ORG 35035 (ED50 = 1.9 mg/kg s.c.) and the reference compound m-CPP (ED50 = 0.3 mg/kg s.c.) induced penile erection in male rats (an effect that is only very infrequently observed in this test situation after vehicle treatment alone). The lowest doses to induce penile erection were 0.1 mg/kg Ro 60-0175/ORG 35030 (3/8 rats), 1 mg/kg Ro 60-0332/ORG 35035 (2/8 rats) and 0.1 mg/kg m-CPP (2/8 rats). A maximal effect was achieved for the two former compounds at approximately 3.2 mg/kg. These results on penile erection are illustrated in figure 2. Neither Ro 60-0175/ORG 35030 nor Ro 60-0332/ORG 35035 elicited head shakes up to the dose 10 mg/kg (the highest dose tested). In contrast, p.o. administration of (±)-DOI failed to induce penile erection up to 10 mg/kg s.c. but dose-dependently increased head twitches, reaching a plateau effect of about 20 bouts (8/8 rats) at the dose 1 mg/kg and above (ED50 = 0.3 mg/kg s.c.). In a preliminary experiment it was shown that pretreatment with the 5-HT2C receptor antagonist Ro 60-0491 completely blocked the elicitation of penile erection by Ro 60-0175/ORG 35030 and Ro 60-0332/ORG 35035 (tested up to 10 mg/kg s.c.) without revealing any new behavioral symptoms—for example, head shakes.

Elicitation of penile erection in rats with (±)-DOI, Ro 60-0175/ORG 35030 and Ro 60-0332/ORG 35035.
Behavioral observation in squirrel monkeys.
Observation of the effects produced by high p.o. doses of fluoxetine or the two experimental compounds was done in male monkeys. Vehicle treatment was used to define the base-line behavioral pattern. Compared with vehicle, the reference compound fluoxetine produced some signs of sedation in 2/10 and of retching and/or vomiting in 5/10 monkeys at 10 mg/kg; 30 mg/kg fluoxetine produced sedation in 5/10, retching and/or vomiting in 7/10, tremors in 2/10 and convulsions in 1/10 monkeys. Ro 60-0175/ORG 35030 induced signs of sedation in half of the monkeys at both 10 and 30 mg/kg, as well as retching and/or vomiting in 1/10 at 10 mg/kg and in only 2/10 at 30 mg/kg. Ro 60-0332/ORG 35035 induced some postural relaxation in 3/5 monkeys at 10 mg/kg, with signs of relaxation and/or sedation in 5/5 monkeys at 30 mg/kg; retching and/or vomiting was seen in only 2/5 monkeys at 30 mg/kg. Penile erection was not observed after any of the treatments in these male monkeys.
Spontaneous motor activity in rats.
We evaluated the effect of prior injection of the 5-HT2C receptor antagonist Ro 60-0491 on the hypoactivity induced by a high dose of each of the 5-HT2C receptor agonists m-CPP (6 mg/kg i.p.), Ro 60-0175/ORG 35030 (10 mg/kg i.p.) and Ro 60-0332/ORG 35035 (10 mg/kg i.p.). The results are illustrated figure3. When given after a vehicle injection, the selected dose of each of these three 5-HT2C receptor agonists significantly reduced total distance (P < .05) compared with vehicle plus vehicle treatment. (The selected dose of Ro 60-0491 given in combination with vehicle somewhat reduced locomotion in these separate experiments, but this reduction reached statistical significance only in the experiment with Ro 60-0332/ORG 35035). Of primary interest, however, was a determination of whether the antagonist Ro 60-0491 would attenuate the hypomotility induced by these 5-HT2C receptor agonists. In fact, Ro 60-0491 significantly reversed the hypoactivity induced by each of these three 5-HT2C receptor agonists (P < .05), although a full return to the base-line activity level was not obtained.

Antagonism of the hypomotility in rats induced with m-CPP, Ro 60-0175/ORG 35030 and Ro 60-0332/ORG 35035. When given together with vehicle, each of these three 5-HT2C receptor agonists significantly reduced activity vs. the vehicle plus vehicle condition. (* P < .05 for comparison with group treated with vehicle and the 5-HT2C receptor agonist.) [Note: The selected dose of the 5-HT2C receptor antagonist Ro 60-0491 given together with vehicle significantly reduced activity compared to the vehicle plus vehicle condition only in the experiment with Ro 60-0332/ORG 35035 (significance not shown in figure).]
Anticonvulsant effects in mice.
Ro 60-0175/ORG 35030 was anticonvulsant in the audiogenic seizure paradigm in DBA/2J mice with an ED50 of 56 mg/kg p.o. In contrast, Ro 60-0332/ORG 35035 protected maximally 25% of the mice against seizures at each of the doses 10, 32 and 100 mg/kg p.o.
Elevated plus-maze task in rats.
A single injection of fluoxetine (5 mg/kg i.p.) significantly reduced, in comparison with vehicle, the number of open arms entered and the time spent in open arms (P = .004 and P = .006, respectively), which indicates an anxiogenic effect, but did not reduce the number of closed-arm entries or attempts; these results show that neither general motor activity nor risk assessment, respectively, was affected. Furthermore, the same pattern of effects was observed after the fifth in a series of daily treatments. In contrast, acute treatment with Ro 60-0175/ORG 35030 (1.5 mg/kg i.p.) failed to affect these same parameters significantly when compared with vehicle. This pattern of results was unchanged after repeated daily injections made on four successive days and followed on the final day by a test in the plus-maze. Ro 60-0332/ORG 35035 (7.5 mg/kg i.p.) did not affect the time spent in the open arms but significantly decreased the number of open-arm entries, closed-arm entries and attempts (P = .02, P = .0001 and P = .02, respectively). However, this pattern of results (especially reduced closed-arm entries) suggests general sensorimotor impairment rather than anxiogenesis. After the fifth in a series of daily treatments, none of these parameters was differentially affected by vehicle and Ro 60-0332/ORG 35035. The detailed results are provided in table 3.
Effects of equieffective doses of fluoxetine, Ro 60-0175/ORG 35030 and Ro 60-0332/ORG 35035 on behavioral parameters in a plus-maze task in rats
Burying behavior in mice.
The lowest dose of fluoxetine that was tested, 10 mg/kg s.c., reduced burying by 55%. The ED50 value for reducing burying behavior was 3.8 mg/kg s.c. for Ro 60-0175/ORG 35030 and 4.4 mg/kg s.c. for Ro 60-0332/ORG 35035.
Reduction of palatable food intake in rats.
All of the test compounds were found to produce a statistically significant overall reduction in palatable food intake for the dose ranges evaluated (P < .05). Fluoxetine (MED = 60 mg/kg p.o.), m-CPP (MED = 0.3 mg/kg p.o.), Ro 60-0175/ORG 35030 (MED = 10 mg/kg p.o.) and Ro 60-0332/ORG 35035 (MED = 30 mg/kg p.o.) significantly reduced consumption (P < .01) in nondeprived rats during the 30-min session. At these MED values, the reduction in palatable food intake for these compounds was approximately 40%, except for m-CPP, where 0.3 mg/kg reduced intake by only 19% (but at 1 mg/kg, intake was reduced by 45%). These data are shown in figure4.

Reduction of palatable food intake in rats by fluoxetine, m-CPP, Ro 60-0175/ORG 35030 and Ro 60-0332/ORG 35035. (* P < .05 for comparison with vehicle controls.)
Reversal of schedule-induced polydipsia task in rats.
All of the test compounds were found to produce a statistically significant overall reduction in both schedule-induced drinking and control drinking (i.e., under the nonstress condition) across the dose ranges evaluated (P < .05). Fluoxetine significantly reduced schedule-induced polydipsia at the high dose of 60 mg/kg p.o., whereas control water intake assessed in the same apparatus was already significantly reduced at 30 mg/kg p.o. In contrast, Ro 60-0175/ORG 35030 significantly reduced schedule-induced polydipsia beginning at 1 mg/kg p.o., a reduction in control water intake first being observed at 10 mg/kg. Ro 60-0332/ORG 35035 significantly decreased both schedule-induced polydipsia and control water intake at 10 mg/kg. These results are shown in figure 5.

Amelioration of schedule-induced polydipsia in rats by fluoxetine, Ro 60-0175/ORG 35030 and Ro 60-0332/ORG 35035. (** P < .01 for comparison with controls.)
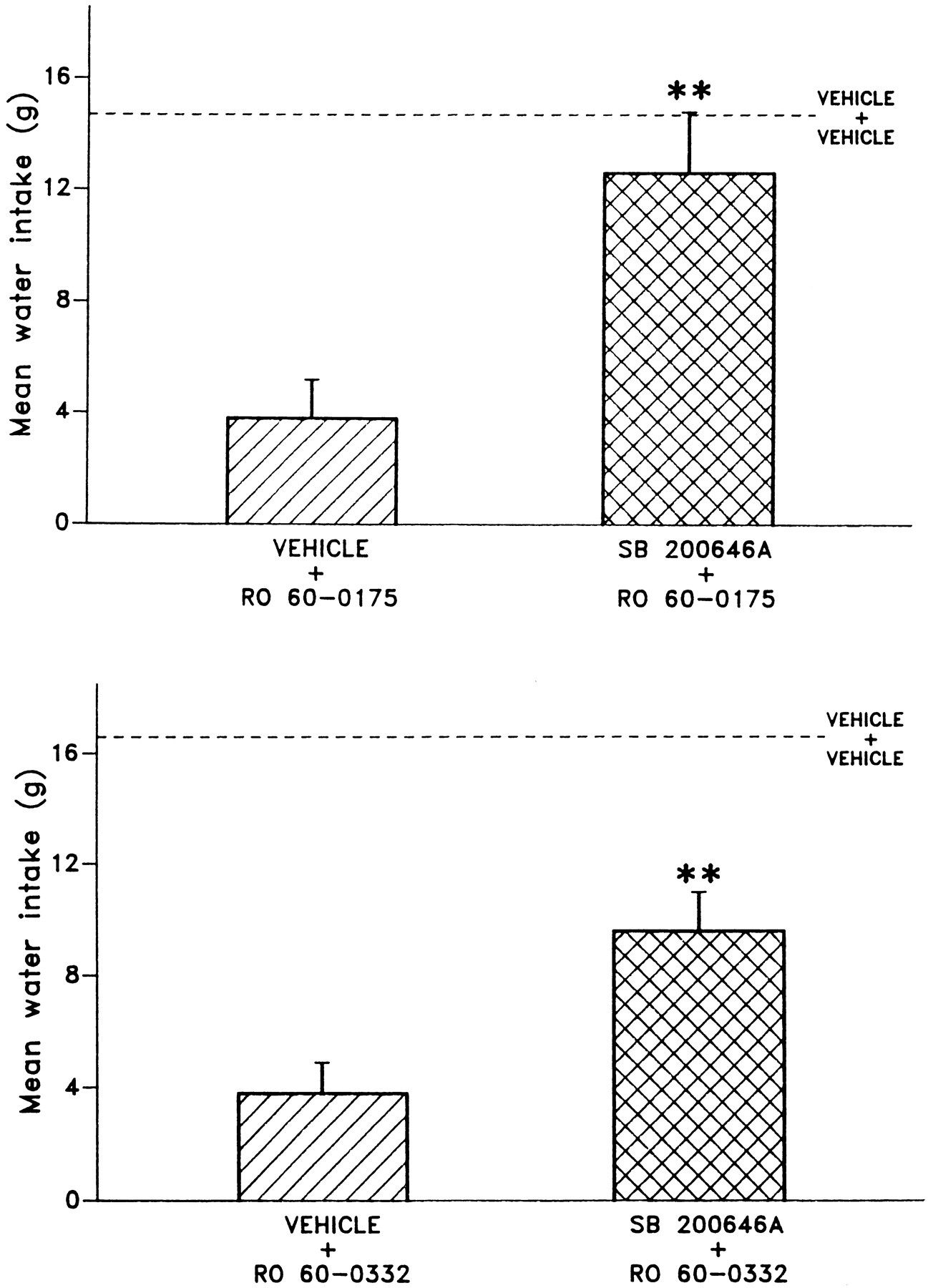
A subsequent experiment was done using the 5-HT2C/2Breceptor antagonist SB 200646A. In combination with a vehicle pretreatment, the selected dose of Ro 60-0175/ORG 35030 (10 mg/kg p.o.) and that of Ro 60-0332/ORG 35035 (20 mg/kg p.o.) significantly reduced schedule-induced polydipsia in comparison with a vehicle plus vehicle condition (P < .01). Pretreatment with SB 200646A (30 mg/kg i.p.) significantly reduced the schedule-induced polydipsia produced by either of these 5-HT2C receptor agonists (P < .01) compared with pretreatment with vehicle. These results are shown in figure 6.

Pretreatment with 5-HT2C/2B receptor antagonist SB 200646A decreases the ameliorative effect of Ro 60-0175/ORG 35030 and Ro 60-0332/ORG 35035 in schedule-induced polydipsia paradigm. (** P < .01 for comparison with vehicle controls.)
Reduction of compulsive behavior in monkeys.
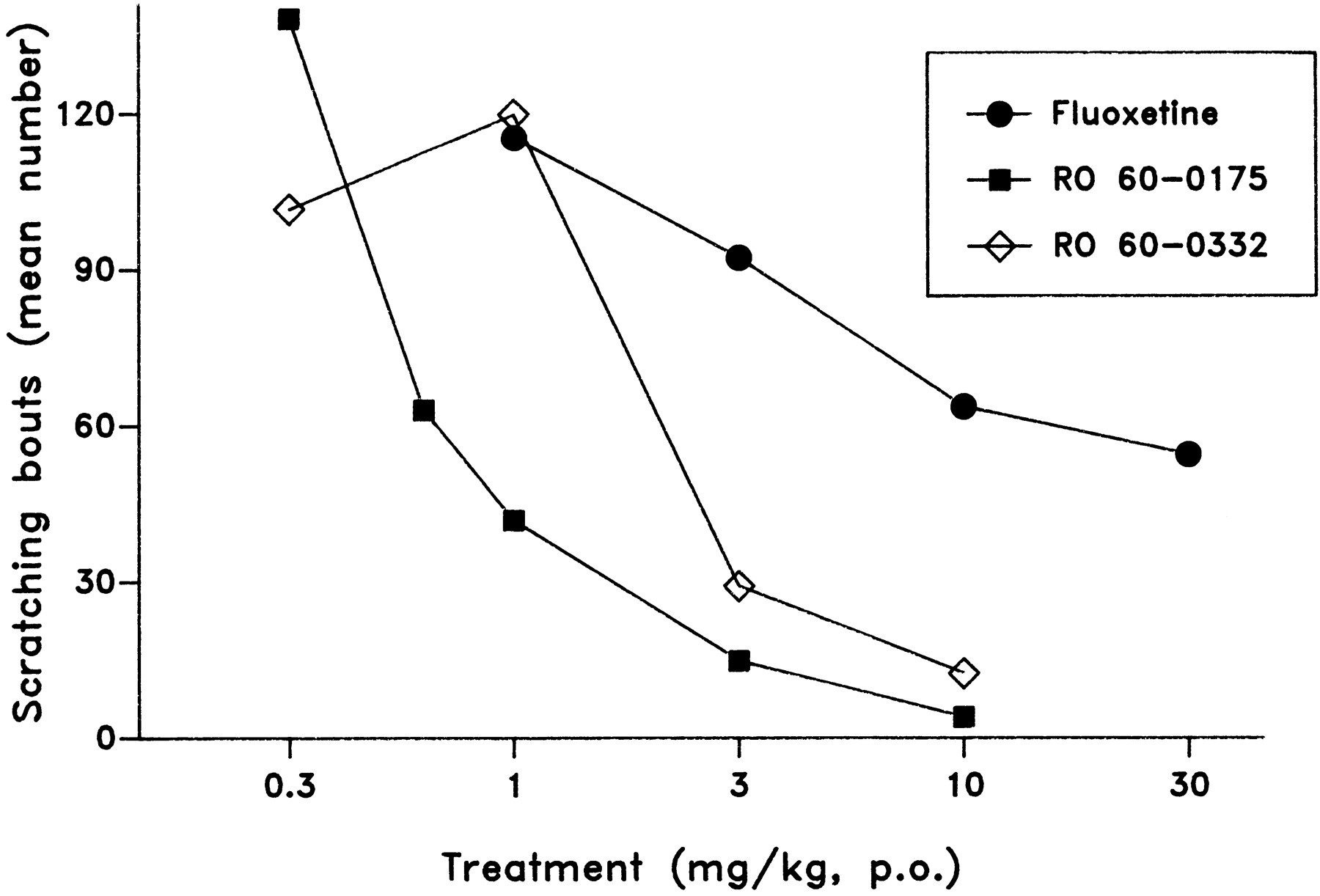
Compulsive whole-body scratching bouts are induced in squirrel monkeys by 8-OH-DPAT, and the effect can be fully reversed by subsequent treatment with 5-HT2C receptor agonists. This provides a robust pharmacodynamic model in monkeys. 8-OH-DPAT produced multiple scratching bouts throughout the 2-h observation period when it was given after vehicle (mean ± S.E. = 86.2 ± 11.4). Compared with vehicle, fluoxetine reduced scratching only beginning at 10 mg/kg (about 26% reduction), with 30 mg/kg (the highest dose tested) producing an only somewhat greater effect (37% reduction). In contrast, both Ro 60-0175/ORG 35030 (ID50 = 0.8 mg/kg) and Ro 60-0332/ORG 35035 (ID50 = 1.1 mg/kg) potently reduced scratching, achieving a full reversal at the dose 3 mg/kg. These results are illustrated in figure 7. In comparison, the nonselective 5-HT2C receptor agonist m-CPP had an ID50 value of 0.3 mg/kg p.o. in this test paradigm (data not included in fig. 7). In the chronic experiment, the anticompulsive effects of 1 mg/kg p.o. Ro 60-0175/ORG 35030 and 10 mg/kg p.o. fluoxetine were of approximately the same magnitude, with a significant difference from vehicle treatment (P < .05). There was no difference observed between the first and the final day for either of these treatments, which indicates that tolerance had not developed.

Reduction of 8-OH-DPAT-induced scratching in squirrel monkeys with fluoxetine, Ro 60-0175/ORG 35030 and Ro 60-0332/ORG 35035 (in these groups, S.E. values were typically about 20–35% of the mean). After vehicle treatment, 8-OH-DPAT induced a mean (± S.E.) of 86.2 ± 11.4 scratching bouts during the 2-h observation period.
Reversal of passive avoidance deficit in olfactory-bulbectomized rats.
Compared with sham-operated control rats receiving vehicle, the rats with verified bilateral olfactory bulbectomies given vehicle required significantly more trials to reach the learning criterion in a passive avoidance task (P < .05). Treatment with Ro 60-0332/ORG 35035 (5 mg/kg s.c.) in bulbectomized rats significantly reduced the number of trials to acquisition compared with vehicle (P < .05). These results are shown in figure 8. The step-down latency on the first trial (before any shock exposure) was comparable in the sham-operated, vehicle-treated group (10.9 ± 2.6 s) and the bulbectomized group receiving Ro 60-0332/ORG 35035 (9.4 ± 1.2 s), whereas that of the bulbectomized, vehicle-treated group was lower (4.6 ± 0.6 sec).

Amelioration of passive avoidance deficit in olfactory bulbectomized rats with Ro 60-0332/ORG 35035. (* P < .05 for comparison with vehicle-treated olfactory bulbectomized rats.)
Responding in DRL-72 s task in rats.
Both Ro 60-0175/ORG 35030 and Ro 60-0332/ORG 35035 significantly decreased the rate of lever pressing only at 10 mg/kg i.p. (P < .01) with no accompanying signs of observable sedation. However, at the highest dose that failed to reduce total number of responses in a session (3 mg/kg i.p.), Ro 60-0175/ORG 35030 significantly increased the number of reinforcements obtained during the session (2.6-fold vehicle base line; P < .01), and Ro 60-0332/ORG 35035 nonsignificantly increased this parameter (1.7-fold vehicle base line).
Sleep-wake pattern in rats.
It has previously been shown that in the EEG-defined rat sleep-wake pattern, an increase in duration of the quiet-waking component, accompanied by a decrease in the REM component, was typical for the general class of antidepressants (Ruigtet al., 1993). Fluoxetine was found to increase quiet waking appreciably and to decrease REM sleep at 10 and 32 mg/kg i.p. Ro 60-0175/ORG 35030 produced a similar pattern of effects at 2.2 mg/kg i.p. and at 10 and 22 mg/kg p.o. Ro 60-0332/ORG 35035 showed this typical antidepressant profile at 3.2 and 10 mg/kg i.p. and at 32 mg/kg p.o. These results are summarized in table4.
Pharmaco-EEG evaluation in free-moving rats (the results are shown as percentage of base-line duration)
Discussion
Serotonin neurotransmission plays an important role in numerous physiological processes both in health and in psychiatric disorders. Determination of the functions of the multiple 5-HT receptor subtypes identified in recent years has been dependent on the availability of selective agonists and antagonists. The present study focused on investigating the putative involvement of 5-HT2C agonism in psychiatric disorders by testing the 5-HT receptor agonists Ro 60-0175/ORG 35030 and Ro 60-0332/ORG 35035 in animal models of compulsive behavior, depression and anxiety.
A broad evaluation of receptor binding affinity was carried out for 35 different receptors. It was demonstrated that both Ro 60-0175/ORG 35030 and Ro 60-0332/ORG 35035 exhibited high affinity for the human 5-HT2C receptor (pKi = 9.0 and 8.5, respectively) and considerably lower affinity for the human 5-HT2A receptor (pKi = 7.5 and 7.0, respectively). Affinity of these two compounds for numerous other selected receptors was at least 2 to 3 logarithmic units weaker than for the 5-HT2C receptor. Both of these experimental compounds produced a concentration-dependent stimulation of IP3 formation in rat choroid plexus in vitro in a manner similar to 5-HT itself, an effect that demonstrates their 5-HT2C receptor agonism. In an experiment using NIH 3T3 cells expressing either h5-HT2C or h5-HT2Areceptors, EC50 values for stimulation of IP3formation were also determined: Ro 60-0175/ORG 35030 (2C: 287 nM; 2A: 671 nM), Ro 60-0332/ORG 35035 (2C: 315 nM; 2A: 1496 nM) and reference compound m-CPP (2C: 357 nM; 2A: 1384 nM). Under this set of test conditions, selectivity for h5-HT2Cvs.h5-HT2A receptors was about 2- to 5-fold (unpublished data). Both experimental compounds elicited penile erection in rats, a behavioral sign of the serotonin syndrome in rodents, which is indicative of a 5-HT2C agonistic profile (Berendsenet al., 1990; Millan et al., 1997); the virtual absence of concomitant head shakes is a clear indication of the absence of any appreciable 5-HT2A receptor agonism in the in vivo situation (it is interesting to note that SSRIs also elicit penile erections in rats; see Berendsen, 1995).
Results from the isolated rat fundus strip assay indicate that Ro 60-0175/ORG 35030 (pD2 = 7.8, 8.0; with a maximal response of only 70% of that obtained with 5-CT) and Ro 60-0332/ORG 35035 (pD2 = 6.1) act as agonists at the 5-HT2Breceptor (unpublished data). These results, together with data on the potencies of these two compounds for stimulating IP3formation in vitro in tissue and NIH 3T3 cells, suggest that there may be at most only minimal selectivity for 5-HT2Cvs. 5-HT2B receptors. Little is currently known about the function of 5-HT2B receptors, in part because of the lack of highly selective ligands. It has been reported that this receptor is found in the GI tract, the heart and possibly the brain (Baxter et al., 1995; Duxon et al., 1995; Loricet al., 1992), as well as in the meninges (Schmuck et al., 1996). It has been reported that the preferential 5-HT2B agonist BW723C86 exhibits both anxiolytic and hyperphagic effects in rats (Ainsworth et al., 1996; Kennettet al., 1996). At the present time, however, it is difficult to define the possible contribution of 5-HT2B receptor agonism to the pharmacological profiles of Ro 60-0175/ORG 35030 and Ro 60-0332/ORG 35035; this should receive attention in further studies.
Ro 60-0175/ORG 35030 and Ro 60-0332/ORG 35035 were well tolerated in squirrel monkeys, signs of sedation being the most commonly observed side effect at high doses. Tests with a number of structurally different 5-HT2C receptor agonists, including these two compounds, consistently produced penile erection in rats but not in squirrel monkeys. In addition, these two compounds were shown to be weakly anticonvulsant in mice, which, in view of the high doses required, is probably not related to their 5-HT2C receptor agonism. Hypomotility induced in rats by a high dose of either of these two compounds was significantly attenuated by pretreatment with the 5-HT2C receptor antagonist Ro 60-0491. In the elevated plus-maze task in rats, Ro 60-0175/ORG 35030 and Ro 60-0332/ORG 35035 failed to exhibit either an anxiogenic effect or an anxiolytic effect (whereas fluoxetine was anxiogenic).
In the burying test in mice, Ro 60-0175/ORG 35030 and Ro 60-0332/ORG 35035 were effective in significantly reducing this compulsive burying of novel objects. Similarly, excessive consumption of palatable food by nondeprived rats was significantly decreased by these two compounds, as previously reported for nonselective 5-HT2C receptor agonists in tests of food consumption (Kennett, 1993). However, this may represent an anorectic effect rather than attenuation of compulsive responding per se. Schedule-induced polydipsia in rats has been proposed, on the basis of pharmacological validation, as an animal model of OCD (Wood et al., 1993). Ro 60-0175/ORG 35030 and Ro 60-0332/ORG 35035 significantly attenuated excessive drinking in this task, a result that suggests their possible therapeutic value in OCD (fluoxetine was less potent and exhibited a weaker effect up to the highest tolerated dose). The ameliorative activity of both compounds could be reversed by pretreatment with the 5-HT2C/2Breceptor antagonist SB 200646A. However, in this same experimental paradigm the preferential 5-HT2A receptor antagonist ketanserin tartrate (30 mg/kg i.p.) was ineffective in reversing the ameliorative effect of either of these two compounds (unpublished data). In squirrel monkeys, 8-OH-DPAT injection elicits excessive whole-body scratching that lasts for several hours. Pretreatment with Ro 60-0175/ORG 35030 and Ro 60-0332/ORG 35035 potently and fully prevented this excessive behavior, which, in view of their low affinity for the 5-HT1A receptor, is probably due to an inhibitory interaction with 5-HT2C receptors (Berendsen et al., 1990; Jenck et al., 1994). Furthermore, there was no tolerance to this pharmacological effect over a 2-week treatment period with either Ro 60-0175/ORG 35030 or fluoxetine. Interestingly, fluoxetine (which, along with other SSRRs, is approved for the therapy of OCD) was effective in attenuating excessive behavior in these diverse animal tests. However, either the potency of fluoxetine was lower than that of Ro 60-0175/ORG 35030 and Ro 60-0332/ORG 35035 or the effect size was smaller for fluoxetine (or both).
Antidepressants have been demonstrated to ameliorate the impairment in passive avoidance learning induced by olfactory bulbectomy in rats (Broekkamp et al., 1980). In the present investigation, Ro 60-0332/ORG 35035 also significantly reduced the deficit in bulbectomized rats. In addition, Ro 60-0175/ORG 35030 significantly, and Ro 60-0332/ORG 35035 nonsignificantly, increased the total number of reinforcements obtained by rats in the DRL-72 s task, an effect that is suggestive of possible antidepressant activity (O’Donnell and Seiden, 1983). As further evidence of possible therapeutic potential of 5-HT2C receptor agonists in depression, it was shown that in a pharmaco-EEG paradigm in free-moving rats, treatment with fluoxetine, Ro 60-0175/ORG 35030 and Ro 60-0332/ORG 35035 appreciably increased the quiet-waking component and decreased REM sleep, an EEG profile typical of antidepressants (Kupfer et al., 1981). Such results are consistent with these compounds’ effectiveness in ameliorating mild stress-induced anhedonia in rats (Moreau et al., 1996).
In conclusion, using the 5-HT2C receptor agonists Ro 60-0175/ORG 35030 and Ro 60-0332/ORG 35035 as research tools, it was possible to demonstrate both their excellent tolerability and their therapeutic potential for OCD and depression. Recently reported evidence for the effectiveness of both of these compounds in preventing and in curing mild stress-induced anhedonia and attenuating experimentally induced panic in rats further substantiates the therapeutic potential of 5-HT2C receptor agonists in psychiatry (Jenck et al., in press; Moreau et al., 1996).
Acknowledgments
The authors wish to thank P. Biry, J.-C. Buob, C. Carolo, F. Kahn, S. Koch, P. Mortas, R. Mory, A. Rudler, M. Schmitt, B. Thomas and C. Zwingelstein for their valuable technical assistance in conducting the experiments and gratefully acknowledge the critical advice of Drs. C. D. Nicholson and U. Widmer. In addition, we thank Drs. D. Leysen and U. Widmer for the synthesis of several of the compounds used as research tools.
Footnotes
-
Send reprint requests to: Dr. J. R. Martin, PRPN-D, Bldg. 69/410, F. Hoffmann-La Roche Ltd, Grenzacherstrasse 124, CH-4070 Basel, Switzerland.
- Abbreviations:
- m-CPP dihydrochloride
- 1-(3-chlorophenyl)piperazine 1:2 HCl
- 5-CT
- 5-carboxamidotryptamine
- DOB
- 1-(4-bromo-2,5-dimethoxyphenyl)-2-aminopropane
- (±)-DOI hydrochloride
- rac-4-iodo-2,5-dimethoxy-α-methylbenzylamine 1:1 HCl
- 8-OH-DPAT hydrochloride
- 8-hydroxy-dipropylaminotetralin 1:1 HCl
- SB200646A
- N-(1-methyl-5′-indolyl)-(3-pyridyl)urea 1:1 HCl
- Ro 60-0175/ORG 35030
- (S)-2-(chloro-5-fluoro-indol-1-yl)-1-methylethylamine 1:1 C4H4O4
- Ro 60-0332/ORG 35035
- (S)-2-(4,4,7-trimethyl-1,4-dihydro-indeno[1,2-b]pyrrol-1-yl)-1-methylethylamine 1:1 C4H4O4
- Ro 60-0491/001
- N-(2-naphthyl)-N′-(3-pyridyl)-urea 1:1 HCl
- 5-HT
- serotonin
- DRL-72 s
- differential-reinforcement-of-low-rate 72-s operant schedule
- EEG
- electroencephalogram
- EMG
- electromyogram
- FT-1 min
- fixed-time 1-min operant schedule
- IP3
- inositol 1,4,5-triphosphate
- OCD
- obsessive compulsive disorder
- p.o.
- per os
- REM
- rapid eye movement
- SSRI
- selective serotonin reuptake inhibitor
- Received November 21, 1996.
- Accepted April 22, 1998.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}