


Abstract
The mechanism for the efflux of taurocholic acid (TC) across the blood-brain barrier (BBB) was studied by examining the elimination of [3H]TC after microinjection into the cerebral cortex. The efflux of [3H]TC from the brain was saturable with a Vmax of 15.0 pmol/min/g brain and aKm value of 0.396 nmol/0.2 μl injectate. Efflux was inhibited by cholic acid (CA), a cationic cyclic octapeptide (octreotide; a somatostatin analogue) and an anionic cyclic pentapeptide (BQ-123; an endothelin receptor antagonist), with an IC50 value of 1.09 nmol/0.2 μl injectate, 1.12 nmol/0.2 μl injectate and 0.12 nmol/0.2 μl injectate, respectively. Probenecid (20 nmol/0.2 μl injectate), but notp-aminohippuric acid (10 nmol/0.2 μl injectate), inhibited the brain efflux of [3H]TC. In addition, elimination of [3H]BQ-123 after microinjection was saturable with a Vmax of 20.8 pmol/min/g brain and aKm of 2.92 nmol/0.2 μl injectate; it was also inhibited by TC with an IC50 value of 0.074 nmol/0.2 μl injectate. In contrast, no significant efflux of [14C]octreotide from the brain was observed until 60 min after microinjection. These results suggest that both TC and BQ-123 are transported from the brain to the circulating blood across the blood-brain barrier via specific mechanisms. Although mutual inhibition was observed between TC and BQ-123, kinetic analysis suggested that the two transport systems differ.
Because the BBB consists of cerebral endothelial cells connected to each other by tight junctions, the ligand molecules in the circulating blood must be transported transcellularly to enter the CNS (Pardridge, 1991). Moreover, pinocytotic vesicles or fenestra are rarely observed in cerebral endothelial cells (Pardridge, 1991). Due to these anatomical features, BBB penetration of ligands with high hydrophilicity and/or high molecular weight is restricted (Pardridge, 1991). In addition to these static phenomena, the BBB can actively transport some ligands from the brain; it is well established that the brain entry of amphipathic cationic or natural compounds is restricted by P-glycoprotein located on the luminal membrane of cerebral endothelial cells as summarized by Tsuji and Tamai (1997). This concept is based on the findings that 1) P-glycoprotein is localized on the luminal membrane of cerebral endothelial cells, 2) concomitant administration of inhibitors of P-glycoprotein results in increased uptake of its substrates by the brain and 3) the transport function of P-glycoprotein is associated with cerebral endothelial cells cultured in vitro (Tsuji and Tamai, 1997). Finally, this concept was established by in vivo experiments with mdr-knock out mice (Schinkel et al., 1997); it was shown that the brain uptake of P-glycoprotein substrates is markedly enhanced in mdr-knock out mice (Schinkel et al., 1997; Kusuhara et al., 1997).
Furthermore, cumulative evidence suggests the presence of an efflux transporter for organic anions on the BBB as summarized recently (Suzuki et al., 1997). By analyzing the time-dependent change in the brain uptake index, Cornford et al. (1985)suggested asymmetrical transport of valproic acid across the BBB. Using cefodizime, a β-lactam antibiotic, as a model compound, we have suggested the presence of an active efflux transport system for organic anions across the BBB by a kinetic analysis of the time-profiles of the ligand concentration in the brain and CSF after i.v. and intracerebroventricular administration (Suzuki et al., 1997). Based on such kinetic analysis, we also ascribed the limited brain distribution of new quinolones to active efflux across the BBB (Ooie et al., 1997). Dykstra et al. (1993) also reported the effect of probenecid on the efflux of [3H]azidothymidine from the brain ECF after its administration via a microdialysis probe. We have recently confirmed the findings of Dykstra et al. (Takasawa et al., 1997a, b). The presence of an asymmetric transport system on the BBB has also been suggested using probenecid (Deguchi et al., 1997). In addition, by analyzing the amount remaining in the brain after microinjection into the cerebral cortex (Leininger et al., 1991; Kakee et al., 1996), we and others have succeeded in demonstrating a specific transport system for several organic anions including PAH and 1-naphthyl-β-d-glucuronide (Leininger et al., 1991; Kakee et al., 1997).
In addition to the substrates for these transporters, bile acids may be pumped out from the brain; it has been reported that the serum concentration of bile acids is markedly increased in patients suffering from hepatitis and/or cirrhosis (Seeff, 1996). In particular, during the icteric phase of acute viral hepatitis, the plasma concentration of bile acids is increased 50- to 100-fold (Seeff, 1996). To maintain the homeostasis of neuronal function under such disease states, the brain entry of bile acids may be restricted by some transporter(s) located on the BBB. The purpose of our study is to examine this hypothesis using TC. Because we found that the brain penetration of TC is restricted inin situ brain perfusion experiments, the efflux of TC from the brain was further examined by injecting TC into the cerebral cortex. In view of the previously described mutual inhibition by a cationic cyclic octapeptide (octreotide; a somatostatin analogue) of the hepatic uptake of TC (Terasaki et al., 1995; Yamadaet al., 1996, 1997), along with the competitive inhibition of the hepatic uptake of an anionic cyclic pentapeptide (BQ-123; an endothelin receptor antagonist) by TC (Nakamura et al., 1996), we examined the mutual inhibitory effect on the efflux of TC and these peptides after microinjection into the cerebral cortex.
Materials and Methods
Materials.
[3H]TC (2 Ci/mmol), [14C]carboxy inulin (2.3 mCi/g) and [3H]inulin (325 mCi/g) were purchased from NEN Life Science Products, Inc. (Boston, MA). Unlabeled TC, CA and PAH were purchased from Wako Pure Chemical Industries Ltd. (Osaka, Japan). ICG was purchased from Daiichi Pharma Ltd. (Tokyo, Japan). BSP, probenecid, TEMA and d-tubocurarine were purchased from Sigma Chemical Co. Ltd. (St. Louis, MO). DBSP was purchased from Societe d’Etudes et de Recherches Biologiques (Paris, France). Unlabeled and [3H] labeled BQ-123 (31 Ci/mmol) were kindly supplied by Banyu Pharma Ltd. (Ibaraki, Japan). Unlabeled and [14C] labeled octreotide (Sandostatin) (46.2 Ci/mmol) were kindly supplied by Sandoz Pharma Ltd (Basel, Switzerland). All other chemicals were of reagent grade.
Male Wistar rats (weight range, 250–300 g) were purchased from Nippon Ikagaku Doubutsu Ltd. (Tokyo, Japan). They had free access to food and water. Our study was approved by the University of Tokyo Animal Care Committee.
In situ rat brain perfusion of [3H]TC.
The influx clearance of [3H]TC across the BBB was determined by an in situ brain perfusion method reported previously (Takasato et al., 1984). The perfusate containing NaCl 128 mM, NaHCO324 mM, KCl 4.2 mM, NaH2PO4 2.4 mM, CaCl2 1.5 mM, MgCl2 0.9 mM, D-glucose 6.0 mM, (pH 7.4), [3H]TC (0.14 mCi/ml) and [14C]inulin (0.07 mCi/ml) was infused into the external carotid artery at a rate of 5 ml/min. Rats were decapitated 0.5, 1 and 2 min after infusion. The radioactivity in the hemisphere ipsilateral to the perfusion was determined. The volume of distribution of TC at each time point was determined by dividing the amount of isotope associated with the brain (dpm/g brain) by the isotope concentration in the perfusate (dpm/ml perfusate) after correcting for the isotope content remaining in the cerebral vasculature (Takasato et al., 1984). The PS product for [3H]TC across the BBB was calculated as reported previously (Takasato et al., 1984).
Intracerebral microinjection technique.
The in vivo brain efflux experiments were carried out according to the method described previously (Kakee et al., 1996). Male Wistar rats were anesthetized with intramuscular doses of ketamine hydrochloride (235 mg/kg) and xylazine (2.3 mg/kg), and were placed on a warm-up plate whose surface temperature was maintained at 37.0°C by circulating hot water. After exposure of the skull, a 1.0-mm hole was made with a dental drill 0.20 mm anterior and 5.5 mm lateral to the bregma. A stereotaxic frame was used to determine the coordinates of the rat brain. The microinjection needle (o.d. 330 nm), which was fitted with a 5-μl microsyringe was inserted into the hole to a depth of 4.5 mm below the bregma [Parietal Cortex Area 2 (Par 2) region] with the aid of a stereotaxic apparatus. Physiological buffer, 0.20 μl (NaCl 122 mM, NaHCO3 25 mM, KCl 3 mM, K2HPO4 0.4 mM, CaCl2 1.4 mM, MgSO4 1.2 mM, d-glucose 10 mM, HEPES 10 mM, saturated with 95 %O2-5 %CO2, pH 7.40) containing [3H]TC (4 pmol/0.2 μl; 8 nCi/0.2 μl), [3H]BQ-123 (0.258 pmol/0.2 μl; 8 nCi/0.2 μl) or [14C]octreotide (40 pmol/0.2 μl; 1.85 nCi/0.2 μl) was administered to the brain. As an impermeable marker, [14C] or [3H] labeled inulin was simultaneously injected with [3H] or [14C] labeled test ligands, respectively. After microinjection, CSF was collected from the cisterna magma, according to the method reported previously (Kakee et al., 1996). Immediately after the collection of CSF, rats were decapitated and the ipsilateral (left) and contralateral (right) cerebrum and cerebellum were removed. Brain specimens were dissolved in 2.5 ml 2 N NaOH at 50°C for 3 hr to determine the radioactivity. An appropriate crossover correction was made to separate the radioactivity of [3H] and [14C] using a liquid scintillation counter (LC 6000, Beckmann Instruments, Inc., Fullerton, CA). To characterize the efflux transport system, unlabeled ligands were added to the injectate.
Quantification of the ligand efflux from the brain.
The BEI was defined as (amount of ligand eliminated from the brain for a certain period of time) divided by (amount of ligand injected into the cerebral cortex) (Kakee et al., 1996). The following equation was used to calculate the BEI values.
Results
Influx of [3H]TC into the brain.
The volume of distribution of [3H]TC at each time point was determined by using an in situ brain perfusion method. The initial distribution volume of [3H]TC was calculated as 13.4 μl/g brain, being attributed to the sum of the vascular space of the brain capillary (10∼20 μl/g brain; Pardridge et al., 1990) and adhesion to the surface of the brain capillary endothelial cells. The distribution volume of [3H]TC in the brain did not increase with the perfusion period (table1), indicating no significant uptake during the experimental period.
Uptake of [3H]TC into the brain
Time-profiles for the elimination of [3H]TC efflux from the brain.
The amount of isotopes remaining in the ipsilateral cerebrum was determined as a function of time after the microinjection. No significant decrease was observed for the residual amount of radiolabeled inulin up to 40 min after microinjection, indicating that inulin can be used as an impermeable marker. Figure1 shows that the time-profile of (100−BEI) for [3H]TC can be described by a monoexponential equation. Linear regression analysis showed that the rate constant for the efflux of [3H]TC was 0.0233 min−1 with a half-life of approximately 30 min. During the efflux studies, the amount of [14C]inulin and [3H]TC in the contralateral cerebrum, cerebellum and CSF was less than 4% that observed in the ipsilateral cerebrum, indicating limited diffusion into these CNS regions from the injection site.
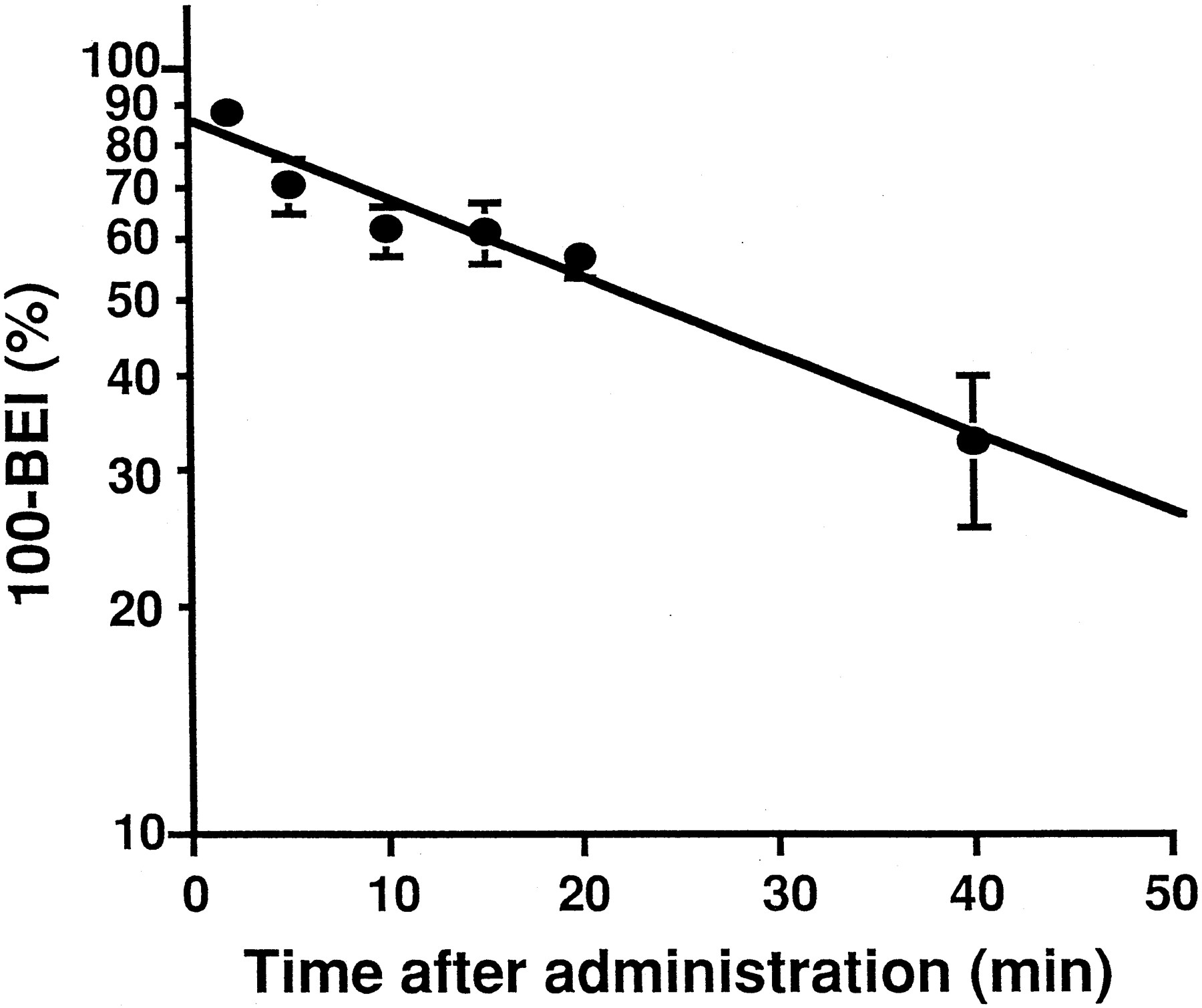
Time profile of [3H]TC remaining in the brain after microinjection into the cerebral cortex. Physiological buffer, 0.2 μl, containing [3H]TC and [14C]inulin was injected into the rat cerebral cortex. Each value represents the mean ± S.E. of (100 − BEI) of three independent experiments. The solid line represents the regression line.
Concentration-dependent efflux of [3H]TC from the brain.
The concentration-dependent efflux of TC from the brain is illustrated in figure 2A. The rate constant for the efflux of [3H]TC decreased with an increase in the amount of unlabeled TC in the injectate, suggesting that TC is pumped out from the brain via a specific transport system. Because no inhibitory effect of unlabeled TC (1 nmol/0.2 μl injectate) was observed on the efflux of [3H]3-O-methyl glucose and [14C]inulin (data not shown), the concentration-dependent efflux of [3H]TC from the brain is not attributed to a toxic effect of TC on the BBB, but to saturation of the efflux transport system. Nonlinear least-squares regression analysis provided a Vmax of 15.0 pmol/min/g brain and aKm value of 0.396 nmol/0.2 μl injectate.

Effect of unlabeled TC and CA on the efflux of [3H]TC from the brain after microinjection into the cerebral cortex. Physiological buffer, 0.2 μl, containing [3H]TC, [14C]inulin and unlabeled TC (A) or CA (B) was injected into the rat cerebral cortex. Each point represents the mean ± S.E. of three independent experiments.
Effect of unlabeled substrates on the efflux of [3H]TC from the brain.
The effect of organic anions and cations on the efflux of [3H]TC from the brain is summarized in table 2. Although probenecid (20 nmol/0.2 μl injectate) reduced the efflux of [3H]TC, no inhibitory effect of PAH (10 nmol/0.2 μl injectate), ICG (1 nmol/0.2 μl injectate), BSP (0.5 nmol/0.2 μl injectate), DBSP (1 nmol/0.2 μl injectate), d-tubocurarine (6 nmol/0.2 μl injectate) or TEMA (10 nmol/0.2 μl injectate) was observed. The rate constant for the efflux of [3H]TC decreased with an increase in the amount of unlabeled CA in the injectate with an IC50 of 1.09 nmol/0.2 μl injectate (fig. 2B).
Effect of unlabeled substrates on the efflux of [3H]TC from the brain after microinjection into the cerebral cortex
The effect of BQ-123 and octreotide on the efflux of [3H]TC from the brain is illustrated in figure3A and B. The efflux of [3H]TC was reduced by BQ-123 and octreotide with an IC50 of 0.116 nmol/0.2 μl injectate and 1.12 nmol/0.2 μl injectate, respectively (table 3).

Effect of unlabeled octreotide and BQ-123 on the efflux of [3H]TC from the brain after microinjection into the cerebral cortex. Physiological buffer, 0.2 μl, containing [3H]TC, [14C]inulin and unlabeled octreotide (A) or BQ-123 (B) was injected into the rat cerebral cortex. Each point represents the mean ± S.E. of three independent experiments.
Kinetic parameters for the efflux of [3H]TC and [3H]BQ-123 from the brain after microinjection
Efflux of [3H]BQ-123 and [14C]octreotide from the brain.
Figure4 illustrates the time-profile of (100−BEI) for [3H]BQ-123. Linear regression analysis yielded an efflux rate constant for [3H]BQ-123 of 0.00783 min−1 with a half-life of approximately 100 min. During the efflux studies, the amount of [14C]inulin and [3H]BQ-123 in the contralateral cerebrum, cerebellum and CSF was less than 4% that observed in the ipsilateral cerebrum.

Time profile of [3H]BQ-123 remaining in the brain after microinjection into the cerebral cortex. Physiological buffer, 0.2 μl, containing [3H]BQ-123 and [14C]inulin was injected into the rat cerebral cortex. Each value represents the mean ± S.E. of (100 − BEI) of three independent experiments. The solid line represents the regression line.
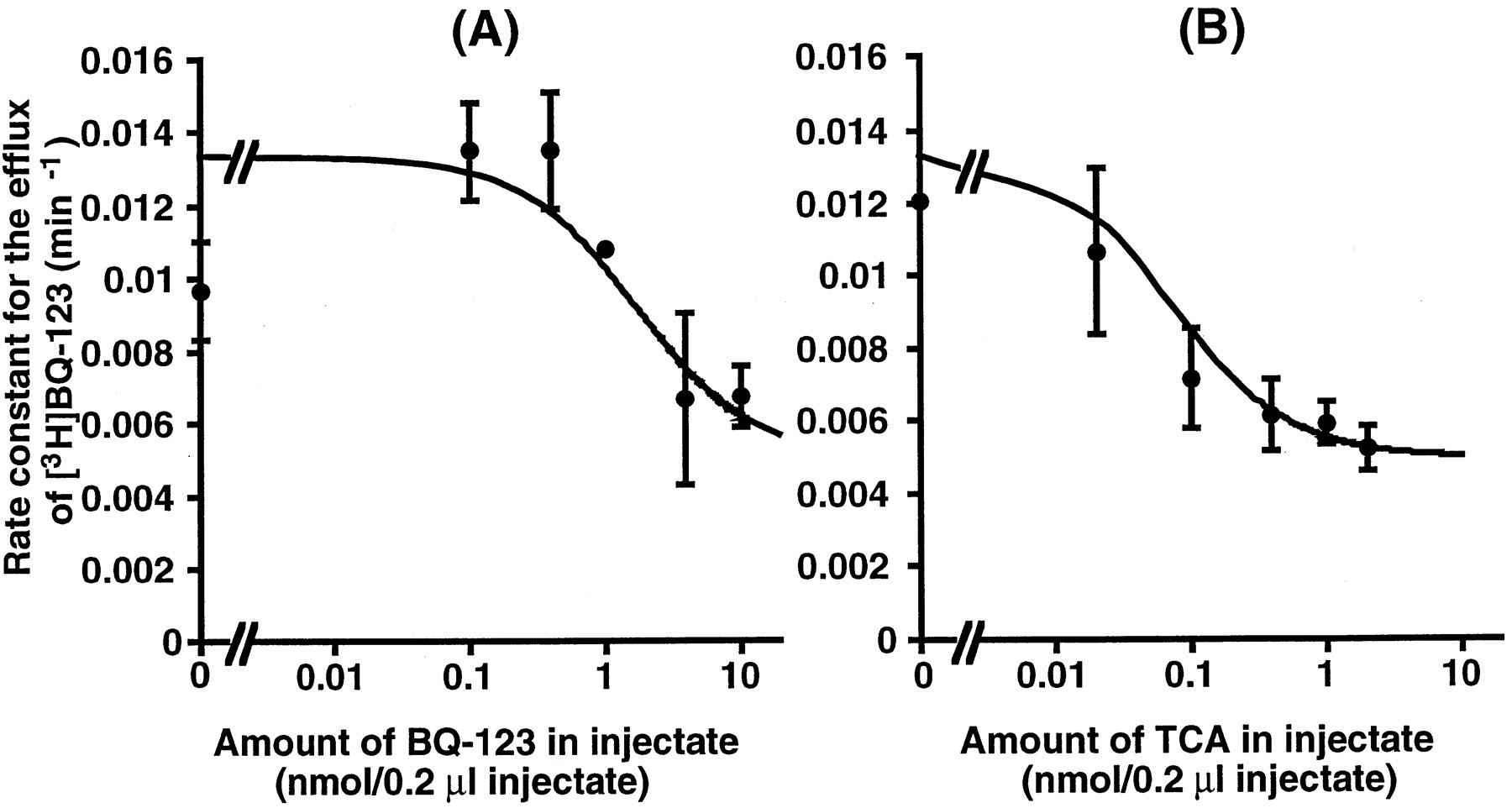
The concentration-dependent efflux of [3H]BQ-123 from the brain is illustrated in figure 5A. The rate constant for the efflux of [3H]BQ-123 decreased with an increase in the amount of unlabeled BQ-123 in the injectate, suggesting the presence of a specific mechanism for the elimination of BQ-123 from the brain. Nonlinear least-squares regression analysis provided a Vmax of 20.8 pmol/min/g brain, aKm of 2.92 nmol/0.2 μl injectate and a Kdiff of 0.00463 min−1.

Effect of unlabeled BQ-123 and TC on the efflux of [3H]BQ-123 from the brain after microinjection into the cerebral cortex. Physiological buffer, 0.2 μl, containing [3H]BQ-123, [14C]inulin and unlabeled BQ-123 (A) or TC (B) was injected into the rat cerebral cortex. Each point represents the mean ± S.E. of three independent experiments.
The effect of TC on the efflux of [3H]BQ-123 from the brain is illustrated in figure 5B. The efflux of [3H]BQ-123 was inhibited by TC with an IC50of 0.074 nmol/0.2 μl injectate (table 3). In contrast, no significant elimination of [14C]octreotide from the brain was observed after microinjection up to 60 min (data not shown).
Discussion
In our study, we examined the transport of TC across the BBB. If the drug molecules are transported across the BBB via passive diffusion, the following relationship between the PS-product and the octanol-to-water partition coefficient (Papp) holds (Levin, 1980; Pardridge et al., 1990):
The elimination of TC from the brain across the BBB should be mediated by the transcellular transport that consists of uptake into the cerebral endothelial cells across the antiluminal membrane and efflux into the blood across the luminal membrane. Although we can neither determine the rate determining process for the transcellular transport nor identify the localization of the efflux transport mechanism on the endothelial cells from our in vivo findings, the transcellular transport of TC across the BBB can be discussed in relation to that across hepatocytes; TC in blood is taken up into hepatocytes across the sinusoidal membrane via Ntcp (Na+/TC cotransporter) and oatp (Na+-independent organic anion transporter), and then excreted into bile via a primary active transporter (Meier, 1995). As shown in figure 2, the efflux of TC across the BBB was inhibited by CA in a dose-dependent manner, which is consistent with a previous observation for the hepatic uptake of TC (Meier et al., 1997; Yamazaki et al., 1996). It is reported that both the Na+-dependent uptake of TC into the sinusoidal membrane vesicles and Na+-independent uptake of TC into isolated hepatocytes was competitively inhibited by CA, which is consistent with the recent findings that the Ntcp-mediated uptake of TC is inhibited by CA and BSP and CA is a substrate for oatp (Meier, 1995; Meier et al., 1997).
Although the Ntcp- and oatp-mediated hepatic uptake of TC was sensitive to BSP, brain efflux of TC was not inhibited by BSP. Efflux of TC across the BBB was not affected by d-tubocurarine, which can inhibit the hepatic uptake of TC (Steen et al., 1992). These results, together with the previous finding that CNS expression of oatp is restricted to the brush border membrane of the choroid plexus (Angeletti et al., 1997), suggest that the transport system for TC is different in cerebral endothelial cells compared with hepatocytes. Because efflux of TC across the BBB was not inhibited by PAH (10 nmol/0.2 μl injectate) at a concentration sufficient to saturate [3H]PAH efflux across the BBB (Km : 2.4 nmol/0.2 μl injectate) (Kakeeet al., 1997), the efflux transport system for TC on the BBB differs from that for PAH.
In the hepatic uptake process, interaction between TC and some cyclic peptides has been reported; BQ-123 is taken up via Na+-dependent and Na+-independent transport systems, both of which are competitively inhibited by TC (Km for Na+-dependent uptake = 13 μM; Na+-independent uptake = 25 μM) withKi values of 9.1 and 9.7 μM, respectively (Nakamura et al., 1996). Although mutually inhibitory effects were observed between the efflux of TC and BQ-123 across the BBB, kinetic analysis indicated that the transport systems for these two ligands are different in cerebral endothelial cells; the IC50 value for TC to inhibit the efflux of BQ-123 (0.074 nmol/0.2 μl injectate) was different from theKm for the efflux of TC (0.386 nmol/0.2 μl injectate). The type of the mutual inhibition (competitive or noncompetitive) is the subject for future studies.
The mechanism for the transport of BQ-123 across the BBB still remains to be clarified. It is possible that BQ-123 is a substrate for the facilitated transporter and/or active transporter responsible for efflux from the brain to the blood; the transport properties of these two kinds of transporters have been characterized using several peptides (Banks and Kastin, 1990; Zlokovic, 1995). Using the brain perfusion method, Zlokovic and his collaborators suggested the presence of a saturable process(es) for the influx of leucine enkephalin and delta sleep inducing peptide into the brain across the BBB (Zlokovic, 1995). Although Banks, Kastin and their collaborators found that Tyr-MIF-1, enkephalins and dynorphin are eliminated from the CNS via specific mechanism(s) (Banks and Kastin, 1990), it is plausible that these peptides are transported across the choroidal epithelial cells. Previous findings by Banks et al. (1989) provided direct evidence to suggest that RC-160, a somatostatin analog, is eliminated from the brain across the BBB; they found saturable elimination of this ligand from the brain after microinjection into the brain parenchyma. Additional studies are required to clarify the relationship between the transport system for BQ-123 and those for the peptides described above.
Regarding the transport of BQ-123 across the bile canalicular membrane, we found that 1) this peptide is excreted into the bile from the hepatocytes via a cMOAT, a primary active transporter recently cloned by this and other laboratories (Paulusma et al., 1996; Itoet al., 1997; Büchler et al., 1996) and 2) the cMOAT-mediated transport of BQ-123 is inhibited by 20 μM TC (Shinet al., 1997). Because we also found that several organic anions are transported in an ATP-dependent manner in a cell line (MBEC4) derived from mouse brain endothelial cells (Kusuhara et al., 1998), it is possible that the efflux of BQ-123 across the luminal membrane of cerebral endothelial cells is mediated by such a primary active transporter.
It has also been reported that, in the presence of Na+, the hepatic uptake of octreotide (Km = 91 μM) was competitively inhibited by TC (Ki = 82 μM) and that of TC (Km = 15 μM) was inhibited by octreotide with an IC50 of 130 μM (Terasaki et al., 1995). Regarding the excretion of octreotide across the bile canalicular membrane, we found that octreotide is transported via a primary active transporter other than cMOAT (Yamada et al., 1996). Because the ATPase activity stimulated by EMD 51921, a linear renin-inhibiting cationic peptide, was not related to cMOAT or P-glycoprotein, irrespective of the fact that the transport of EMD 51921 across the bile canalicular membrane is coupled with the hydrolysis of ATP (Ziegler et al., 1994), it may be that some other primary active transporter is responsible for the excretion of these peptides. Although octreotide reduced the efflux of TC across the BBB with an IC50 value of 1.09 nmol/0.2 μl injectate (fig. 3; table 2), octreotide was not transported across the BBB. It is possible that octreotide acts as an antagonist for the transport of TC across the BBB. Alternatively, TC transport may be inhibited by octreotide in a noncompetitive manner.
In conclusion, as far as we know, this is the first direct demonstration of the presence of a specific mechanism for the active efflux of TC from the brain to the blood across the BBB, although the endogenous ligands for this transporter still remain to be identified. It is possible that this transport system restricts the brain entry of several toxic bile acids whose serum concentration is increased particularly in hepatic failure (Friedman et al., 1996). Although mutual inhibition was observed between TC and BQ-123, kinetic analysis suggests that the two transport systems differ. In addition, it is also suggested that the efflux transport mechanism on the BBB may play an important role in regulating the concentration of biologically active peptides (such as neuropeptides) in the brain ECF. Characterization of the transport mechanism on the BBB may provide us with important information on how to improve the delivery of peptides into the brain.
Footnotes
-
Send reprint requests to: Professor, Yuichi Sugiyama, Graduate School of Pharmaceutical Sciences, The University of Tokyo, Hongo 7-3-1, Bunkyo-ku, Tokyo 113-0033, Japan.
-
↵1 This work was supported in part by a grant-in-aid from the Ministry of Education, Science, Sports and Culture of Japan, and the Core Research for Evolutional Sciences and Technology of Japan Sciences and Technology Corporation.
-
↵2 Current address: Faculty of Pharmaceutical Sciences, Tohoku University, Aramaki azaaoba, Aoba-ku, Sendai, Miyagai 980-77, Japan.
- Abbreviations:
- BBB
- blood-brain barrier
- CNS
- central nervous system
- CSF
- cerebrospinal fluid
- PS product
- permeability-surface area product
- BEI
- brain efflux index
- TC
- taurocholic acid
- CA
- cholic acid
- PAH
- p-aminohippuric acid
- ICG
- indocyanine green
- BSP
- bromosulfophthalein
- TEMA
- tetraethylmethylammonium
- DBSP
- dibromosulfophthalein
- cMOAT
- canalicular multispecific organic anion transporter
- Received January 22, 1998.
- Accepted April 12, 1998.
- The American Society for Pharmacology and Experimental Therapeutics




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}