


Abstract
The effect of selective group I metabotropic glutamate receptor subtype 5 (mGluR5) antagonists 2-methyl-6-(phenylethynyl)-pyridine (MPEP) and (E)-2-methyl-6-(2-phenylethenyl)-pyridine (SIB-1893) on neuronal cell survival and post-traumatic recovery was examined using rat in vitro and in vivo trauma models. Treatment with MPEP and SIB-1893 showed significant neuroprotective effects in rat cortical neuronal cultures subjected to mechanical injury. Application of the antagonists also attenuated glutamate- andN-methyl-d-aspartate (NMDA)-induced neuronal cell death in vitro. Intracerebroventricular administration of MPEP to rats markedly improved motor recovery and reduced deficits of spatial learning after lateral fluid percussion-induced traumatic brain injury. Lesion volumes as assessed by magnetic resonance imaging were also substantially reduced by MPEP treatment. Although we show that MPEP acts as a potent mGluR5 antagonist in our culture system, where it completely blocks agonist-induced phosphoinositide hydrolysis, electrophysiological and pharmacological studies indicate that MPEP and SIB-1893 also inhibit NMDA receptor activity at higher concentrations that are neuroprotective. Taken together, these data suggest that MPEP and SIB-1893 may have therapeutic potential in brain injury, although the mechanisms of neuroprotective action for these drugs may reflect their ability to modulate NMDA receptor activity.
Activation of glutamate receptors has been implicated in the regulation of normal development and functioning of the nervous system, as well as in various pathological processes, including neurodegeneration, ischemia, and neurotrauma (Pin and Duvoisin, 1995; Nakanishi et al., 1998). Glutamate receptors are divided into two major families, ionotropic and metabotropic receptors (mGluRs). Whereas ionotropic glutamate receptors directly regulate ion flow through ligand-gated channels, mGluRs act by influencing intracellular metabolic processes via G-protein-coupled second messenger systems (Holscher et al., 1999). Eight mGluR subtypes have been described and classified into three groups based on structural homology, signal transduction mechanisms, and pharmacological characteristics (Schoepp et al., 1999). Group I mGluRs, which include mGluR1 and 5, are coupled via phospholipase C to the inositol phosphate pathway. Group II (mGluR2 and mGluR3) and group III (mGluR4, mGluR6, mGluR7, and mGluR8) are negatively coupled to adenylyl cyclase (Knopfel et al., 1995; Pin and Duvoisin, 1995; Schoepp et al., 1999).
Group I mGluRs modulate post-traumatic neuronal cell death in different experimental systems (Mukhin et al., 1996, 1997a; Nicoletti et al., 1999). However, determining a role for specific group I mGluRs in neuronal survival has been limited by the absence of highly specific agonists and antagonists. Recently, subtype-selective antagonists both for mGluR1 and mGluR5 have been developed (for review, see Schoepp et al., 1999). Among these are the highly selective, noncompetitive mGluR5 antagonists 2-methyl-6-(phenylethynyl)-pyridine (MPEP) and (E)-2-methyl-6-(2-phenylethenyl)-pyridine (SIB-1893) (Gasparini et al., 1999; Varney et al., 1999).
Based on the use of antisense oligonucleotides and less selective group I antagonists, we have suggested that mGluR1 activation contributes to post-traumatic death, whereas mGluR5 may not (Mukhin et al., 1996). In the present study we examined the influence of mGluR5 subtype-selective antagonists MPEP and SIB-1893 on neuronal cell survival after injury in rat cortical neuronal cultures and on post-traumatic recovery after lateral fluid percussion-induced traumatic brain injury (TBI) in rats. Surprisingly, these compounds exhibited substantial neuroprotective effects through mechanisms that appear to reflect their ability to modulate N-methyl-d-aspartate (NMDA) receptors.
Materials and Methods
Rat Cortical Neuronal Cultures.
Cortical neuronal cultures were derived from rat embryonic cortices (Taconic Farms, Germantown, NY) with modifications of our previously described method (Mukhin et al., 1998). Briefly, cortices from 17- to 18-day-old Sprague-Dawley rat embryos were cleaned from their meninges and blood vessels in Krebs-Ringer bicarbonate buffer containing 0.3% BSA (Life Technologies, Gaithersburg, MD). Cortices were then minced and dissociated in the same buffer with 1800 U/ml trypsin (Sigma, St. Louis, MO) at 37°C for 20 min. After the addition of 200 U/ml DNase I (Sigma) and 3600 U/ml soybean trypsin inhibitor (Sigma) to the suspension, cells were triturated through a 5-ml pipette. After the tissue was allowed to settle for 5 to 10 min, the supernatant was collected, and the remaining tissue pellet was retriturated. The combined supernatants were then centrifuged through a 4% BSA layer and the cell pellet was resuspended in neuronal seeding medium, which consisted of neurobasal medium (Life Technologies) supplemented with 1.1% 100× antibiotic-antimycotic solution (Biofluids, Rockville, MD), 25 μM sodium glutamate, 0.5 mM l-glutamine, and 2% B27 Supplement (Life Technologies). Cells were seeded at a density of 5 × 105 cells/ml onto 96-well tissue culture plates (Corning, Corning, NY) precoated with poly(d-lysine) (70–150 kDa; Sigma), or at a density of 2.5 × 105 cells/ml on 12-mm glass coverslips (Fisher Scientific, Pittsburgh, PA) in 24-well plates precoated with poly(d-lysine). On day 4 in vitro (DIV), feeding media (neuronal seeding medium without sodium glutamate and B27 supplement) in 1:2 proportion was added to cultures. All experiments were performed on cultures at 7 to 9 DIV.
In Vitro Trauma Model.
Mechanical trauma was induced by a specially designed punch device that consists of 28 stainless steel blades joined together; this produces 28 parallel cuts 1.2 mm in length uniformly distributed through the cell layer at 0.5-mm intervals in each well of a 96-well tissue culture plate. There is primary traumatic death of cells located directly under the blades, followed by progressive secondary neuronal death at a distance from the cuts over a period of 24 h. Such secondary cell death was shown to be modulated by mGluRs and NMDA receptors (Mukhin et al., 1997a, 1998). This model has been shown to deliver a highly reproducible and consistent degree of injury to the cells (Mukhin et al., 1998). mGluR5 antagonists dissolved in neurobasal medium were added to cortical neuronal cultures at 30 min before or 10 min after mechanical injury. Control cultures (sister cultures from the same plate) received the same volume of the vehicle alone. Cultures were then incubated in 5% CO2 for 24 h at 37°C before assessment of injury.
Glutamate- and NMDA-Induced Injury.
Either 150 μM sodium glutamate (Sigma) or 100 μM NMDA (Tocris Cookson, Ballwin, MO), dissolved in neurobasal medium, was administered to cell cultures 30 or 15 min after the addition of mGluR5 antagonists, with or without 15-min pretreatment with (R,S)-2-chloro-5-hydroxyphenylglycine (CHPG) (Tocris Cookson). Control cultures (sister cultures from the same 96-well plate) received the same volume of the vehicle alone.
Cell Viability Assay.
Cell viability was measured by LDH release assay (Sinensky et al., 1995) using CytoTox-96 nonradioactive cytotoxicity assay kit (Promega, Madison, WI) according to manufacturer's protocol. Relative absorbance was measured at 490 nm using Ceres 9000 microplate reader (Bio-Tek Instruments, Winooski, VT). Background LDH release determined in intact control cultures was subtracted from all experimental values.
Animals.
Male Sprague-Dawley rats (400–450 g) supplied by Taconic Farms were housed for at least 1 week before any experiments. Rats were maintained at 22°C and 12-h light/dark cycle. All neurological scoring was performed during the light cycle. Food and water were available ad libitum.
Fluid Percussion-Induced TBI.
Rats were anesthetized with sodium pentobarbital (70 mg/kg i.p.), intubated, and implanted with femoral venous and arterial catheters. Brain temperature was followed using a thermister in the temporalis muscle and body temperature was maintained through a feedback-controlled heating blanket. Arterial blood gases were analyzed periodically, and blood pressure was continuously monitored. Lateral fluid percussion-induced injury was performed as previously described (Faden et al., 1989; McIntosh et al., 1989). Animals were subjected to a 2.2- to 2.3-atm pressure pulse, which in our model produces moderate tissue damage in ipsilateral cerebral cortex and hippocampus (Sun and Faden, 1995).
In Vivo Drug Administration.
Intracerebroventricular injections were chosen to examine the effect of the mGluR5 antagonist directly in brain tissue. This technique of drug delivery to the traumatized brain was successfully used by us previously (Faden et al., 1989; Yakovlev et al., 1997; Knoblach and Faden, 1998). Stock solution of MPEP in dimethyl sulfoxide was further diluted with saline to obtain final concentrations for the injections. MPEP (55 μg; Tocris Cookson) in 5 μl of vehicle was administered to rats at 15 min before TBI. Control-treated animals received i.c.v. injections of equal volume vehicle alone. Seven to nine animals were used for each treatment.
Neurological Scoring.
Neuroscores reflect a composite of separate functional evaluations, including resistance to forced lateral pulsion (right and left), forelimb contraflexion on suspension by the tail (right and left), and ability to maintain position on an inclined plane (right, left, and vertical). These tests show high interobserver reliability, and we have used them over more than 10 years to discriminate treatment effects (Faden et al., 1989; Sun and Faden, 1995). Animals were rated from 0 (no function) to 5 (normal function) for each test, with a maximum possible composite score of 35. Evaluations were conducted by an individual unaware of treatment, using procedures detailed previously (Sun and Faden, 1995). With the level of trauma used, improvement typically is observed in untreated animals over the first 2 weeks, after which the deficits stabilize. Treatment effects for neuroprotective agents usually are seen by 1 week, with maximal differences versus controls by 2 weeks (Faden, 1993).
Spatial Learning and Memory Studies.
The Morris water maze (Morris, 1984; Fox et al., 1998) was used to assess spatial learning and memory by training rats to locate a hidden, submerged platform using extramaze visual information. The apparatus used consists of a large, white circular pool (1800 mm in diameter, 750 mm in height, water temperature 22 ± 1°C) with a Plexiglas platform (120 × 120 mm) painted white and submerged 20 mm below the surface of the water (140 mm in height), which is rendered opaque with the addition of dilute, white, nontoxic paint. During training, the platform was hidden in one quadrant 450 mm from the side wall. The rat was gently placed into the water facing the wall at one of four randomly chosen locations separated by 90°. The latency to find the hidden platform within a 90 s criterion time was recorded by a blinded observer. On the first trial, rats failing to find the platform within 90 s were assisted to the platform. Animals were allowed to remain on the platform for 15 s on the first trial and 10 s on all subsequent trials. There was an intertrial interval of 30 min, during which time the rats were towel-dried and placed under a heat lamp. A series of 16 training trials administered in blocks of four were conducted on days 14, 15, 16, and 17 postsurgery.
Lesion Volume Measurements Using MRI.
At 21 days after TBI all animals were anesthetized using sodium pentobarbital (70 mg/kg i.p.) and subjected to MRI using a Bruker 7T/21 cm BioSpec-Avance system (Bruker, Ettlingen, Germany). Briefly, animals were placed in a Plexiglas animal bed with a heating pad warmed to 37°C to maintain the animals' body temperature. Respiratory gating to reduce motion artifacts was achieved using a respiratory monitor as described previously (Albensi et al., 2000). The animal bed was positioned so an animal's head was in the center of the magnet within a 72-mm1H birdcage resonator (Bruker). Field homogeneity across the brain was optimized and a sagittal scout image acquired (RARE image, field of vision = 4 × 4 cm, 128 × 128 resolution, TR/TE = 1500/10 ms with a RARE factor of 8, making the effective TE = 40 ms). Multislice T2-weighted images were then acquired to obtain eight contiguous slices commencing at the end of the olfactory bulb and moving caudal (field of vision = 3 × 3 cm, slice thickness = 2 mm, 128 × 128 resolution, TR/TE = 1500/20 ms, four echo images, and two averages). Each scan took approximately 8 min. Lesion volume was estimated from the summation of areas of hyperintensity on each slice, multiplied by slice thickness, for both the ipsilateral and contralateral hemispheres. Average lesion volume in microliters for each treatment group (±S.E.M.) was calculated and a t test performed to determine significance of reduction in lesion volume compared with control animals.
Phosphoinositide (PI) Hydrolysis.
PI hydrolysis was measured in cortical neuronal cultures at 7 DIV as described previously (Mukhin et al., 1996) with minor modifications. Cortical neuronal cells cultured in 96-well plates were incubated overnight with myo-[3H]inositol (22.3 Ci/mmol; NEN, Boston, MA) at 1 μCi/well. Cells were washed twice with Locke's buffer and incubated at 37°C in the same buffer for 30 min, in the presence or absence of mGluR5 antagonist (0.2–200 μM MPEP). Subsequently, 1 mM CHPG, an mGluR5 agonist, was added together with 20 mM LiCl and incubation was continued for an additional 30 min. Thereafter, incubation buffer was aspirated and inositol phosphates (IP) were extracted by 0.1 M HCl containing 2 mM CaCl2 and transferred to columns with AG 1-X8 anion exchange resin (Bio-Rad, Hercules, CA). After separation according to method of Berridge et al. (1982), accumulated [3H]inositol phosphates were measured using a liquid scintillation counter LS 6500 (Beckman Instruments, Fullerton, CA).
Electrophysiology.
Cortical cultures on glass coverslips were transferred to a recording chamber that was continuously perfused with bath solution at room temperature (21°C), containing 145 mM NaCl, 5 mM KCl, 1 mM CaCl2, 5 mM HEPES, 5 mM glucose, 25 mM sucrose, and 0.01 mM d-serine, adjusted to pH 7.4 with NaOH. Electrodes were pulled from thin-walled borosilicate glass (Blu-Tip capillary tubes; Oxford Labware, St. Louis, MO) in three stages on a horizontal pipette puller (Mecanex S.A., Geneva, Switzerland). Patch pipette solution contained 145 mM potassium gluconate, 5 mM MgATP, 0.2 mM GTP sodium, 5 mM EGTA, and 10 mM HEPES, adjusted to pH 7.2 with KOH. Typical pipette resistance was 5 to 8 MΩ.
The patch-clamp technique was used in the whole-cell recording configuration. Cells were voltage clamped at −60 mV and currents were monitored with an Axopatch 1D amplifier, filtered at 1 kHz, and digitized at 10 to 20 kHz using an IBM-compatible computer equipped with a Digidata 1200 acquisition board (Axon Instruments, Foster City, CA) and pClamp 8 software (Axon Instruments). All drugs were diluted to the required concentration in bath solution, and were applied by a gravity-fed Y-tubing delivery system (Murase et al., 1989) placed within 100 μm of the cell. This delivery system allows rapid onset (<50 ms) and local distribution of the drugs.
Drugs.
Sodium glutamate was obtained from Sigma. NMDA, (5R,10S)-(+)-5-methyl-10,11-dihydro-5H-dibenzo[a,d]cyclohepten-5,10-imine [(+)-MK801], CHPG, SIB-1893, and MPEP were all purchased from Tocris Cookson. MPEP and SIB-1893 were initially dissolved in dimethyl sulfoxide. Stock solutions were thereafter diluted in saline for i.c.v. injections, in culture media for cell viability assays or in Locke's buffer for experiments on PI hydrolysis.
Data Analysis.
Changes in cell viability and PI hydrolysis levels were analyzed by ANOVA, followed by the Student-Newman-Keuls test. Neuroscores were compared with Kruskal-Wallis ANOVA, followed by individual Mann-Whitney Utests. MRI and electrophysiological data were analyzed using one- or two-tailed t tests, respectively. A P value less than 0.05 was considered statistically significant.
Results
Neuroprotective Effects of mGluR5 Antagonists MPEP and SIB-1893 in Vitro.
We analyzed the effects of various concentrations of mGluR5 antagonists MPEP and SIB-1893 on neuronal cell survival after traumatic injury in vitro using an LDH release-based cell viability assay. We have previously demonstrated that changes in LDH release precisely correlate with neuronal cell death in this model, as shown using other methods such as trypan blue staining or ethidium homodimer incorporation assay (Mukhin et al., 1997a). Pretreatment with MPEP or SIB-1893 caused significant dose-dependent neuroprotection in cortical neuronal cultures subjected to traumatic injury (Fig.1). Similar concentrations of the mGluR5 antagonists (20–200 μM) also were protective when administered beginning 10 min after injury (Fig. 2). Addition of MPEP or SIB-1893 to cortical neuronal cultures at concentrations of 20 and 200 μM also significantly attenuated cell death caused by glutamate or NMDA (data not shown).

Pretreatment with mGluR5 antagonists attenuates injury-induced LDH release in rat cortical neuronal cultures. At 7 DIV, indicated concentrations of MPEP (A) or SIB-1893 (B) were added to cultures 30 min before mechanical injury. LDH release was measured after 24 h of treatment. Histograms indicate LDH release as percentage of injured controls ± S.D.; n = 8 to 16 cultures/condition. *P < 0.05 versus vehicle-treated injured controls as shown by ANOVA, followed by the Student-Newman-Keuls test.

Postinjury administration of mGluR5 antagonists significantly reduces injury-induced LDH release in rat cortical neuronal cultures. At 7 DIV, indicated concentrations of MPEP (A) or SIB-1893 (B) were added to cultures 10 min after mechanical injury. LDH release was measured after 24 h of treatment. Histograms indicate LDH release as percentage of injured controls ± S.D.;n = 8 to 16 cultures/condition. *P < 0.05 versus vehicle-treated injured controls as shown by ANOVA, followed by the Student-Newman-Keuls test.
Neuroprotective Effects of MPEP in Vivo.
To explore a possible role for mGluR5 activation in TBI, we examined whether treatment with MPEP could affect post-traumatic recovery of motor function or spatial learning in rats after lateral fluid percussion-induced brain injury. MPEP was administered to rats intracerebroventricularly at 15 min before TBI. Motor and behavioral outcomes were measured as a function of time after injury. As shown in Fig. 3, treatment with MPEP markedly improved post-traumatic recovery. At both 1 and 2 weeks after TBI, MPEP-treated animals showed substantial improvement in composite neuroscores compared with vehicle-treated injured controls (Fig. 3A). MPEP treatment also significantly improved spatial learning after brain trauma, as determined by the Morris water maze test (Fig. 3B). Measurements of lesion volume in traumatized rats, using high field MRI, revealed a significant (about 45%) reduction in lesion volume in rats treated with MPEP (Fig. 4).

Intracerebroventricular administration of MPEP significantly improves recovery of motor function (A) and spatial learning (B) in rats after TBI. The histograms represent composite neurological scores based upon a series of somatomotor tests (A) or median latency to find hidden platform in a version of Morris water maze (B) at indicated times after TBI ± S.D.;n = 7 to 9 animals/treatment. *P < 0.01 versus vehicle-treated controls, by Kruskal-Wallis ANOVA, followed by individual Mann-WhitneyU tests. ▪, control; ▨, MPEP.

MPEP-treated animals develop significantly smaller lesions after TBI than vehicle-treated control animals. A, representative MRI T2-weighted images of control and MPEP-treated rats brains at day 21 after TBI. B, summary of the effects of MPEP treatment on lesion volume after TBI. The histograms represent average lesion volume (μl) at day 21 after injury ± S.E.M. as measured on T2-weighted images; n = 7 to 9 animals/treatment; *P < 0.05 versus vehicle-treated controls using one-tailed t test.
Modulation of mGluR5 Agonist-Induced PI Hydrolysis by MPEP.
To examine the ability of MPEP to act as PLC-coupled mGluR5 antagonist in our in vitro system, we examined the effect of this drug on PI hydrolysis induced in cultured rat cortical neuronal cells by the highly specific mGluR5 agonist CHPG (Doherty et al., 1997). PI hydrolysis was evaluated by measurement of IP accumulation in cortical neuronal cells. As is demonstrated in Fig.5, pretreatment of cultures with MPEP entirely blocked CHPG-induced IP accumulation. MPEP not only inhibited PI hydrolysis at doses shown to be neuroprotective (20 and 200 μM), but was effective at much lower concentrations (0.2 and 2 μM) where neuroprotection was not observed.

Treatment with mGluR5 antagonists blocks mGluR5 agonist-induced PI hydrolysis in rat cortical neuronal cultures. MPEP at indicated concentrations were added to 7 DIV rat cortical neuronal cultures 30 min before stimulation with 1 mM CHPG. PI hydrolysis was measured by IP accumulation within 30 min after addition of CHPG. Histograms represent IP levels as percentage of control ± S.D.;n = 6 cultures/condition. *P < 0.01 versus control cultures compared by ANOVA, followed by the Student-Newman-Keuls test.
Interactions of MPEP and SIB-1893 with NMDA Receptors in Cultured Rat Cortical Neuronal Cells.
We used the whole-cell patch-clamp technique to explore possible interactions of MPEP and SIB-1893 with NMDA receptors. NMDA receptor activation has been shown to be involved in the modulation of post-traumatic neuronal death in our in vitro system (Mukhin et al., 1997a, 1998). Surprisingly, electrophysiological results demonstrated the ability of both MPEP and SIB-1893 to modulate functional properties of NMDA receptors in cultured rat cortical neuronal cells, as indicated by a reduction of the steady-state NMDA-evoked whole-cell current (Fig. 6). Representative traces, showing the effect of a 30-s preperfusion of 200 μM MPEP or 200 μM SIB-1893 on the whole-cell current response evoked by 50 μM NMDA, are illustrated in Fig. 6, A and B. The steady-state responses to drug application were measured, and the data are summarized in Fig. 6C. MPEP produced a 65.88 ± 2.48% reduction, and SIB-1893 caused a 62.05 ± 12.52% reduction of NMDA-evoked current (Fig. 6C). These findings were further confirmed by whole-cell current measurements using lower concentrations of the drugs (20 μM) as well as by the experiments using single-channel recording technique where both MPEP and SIB-1893 (at concentrations of 20 and 200 μM) significantly reduced the open duration of NMDA channels recorded from outside out patches excised from the cortical neurons (O'Leary et al., 2001).
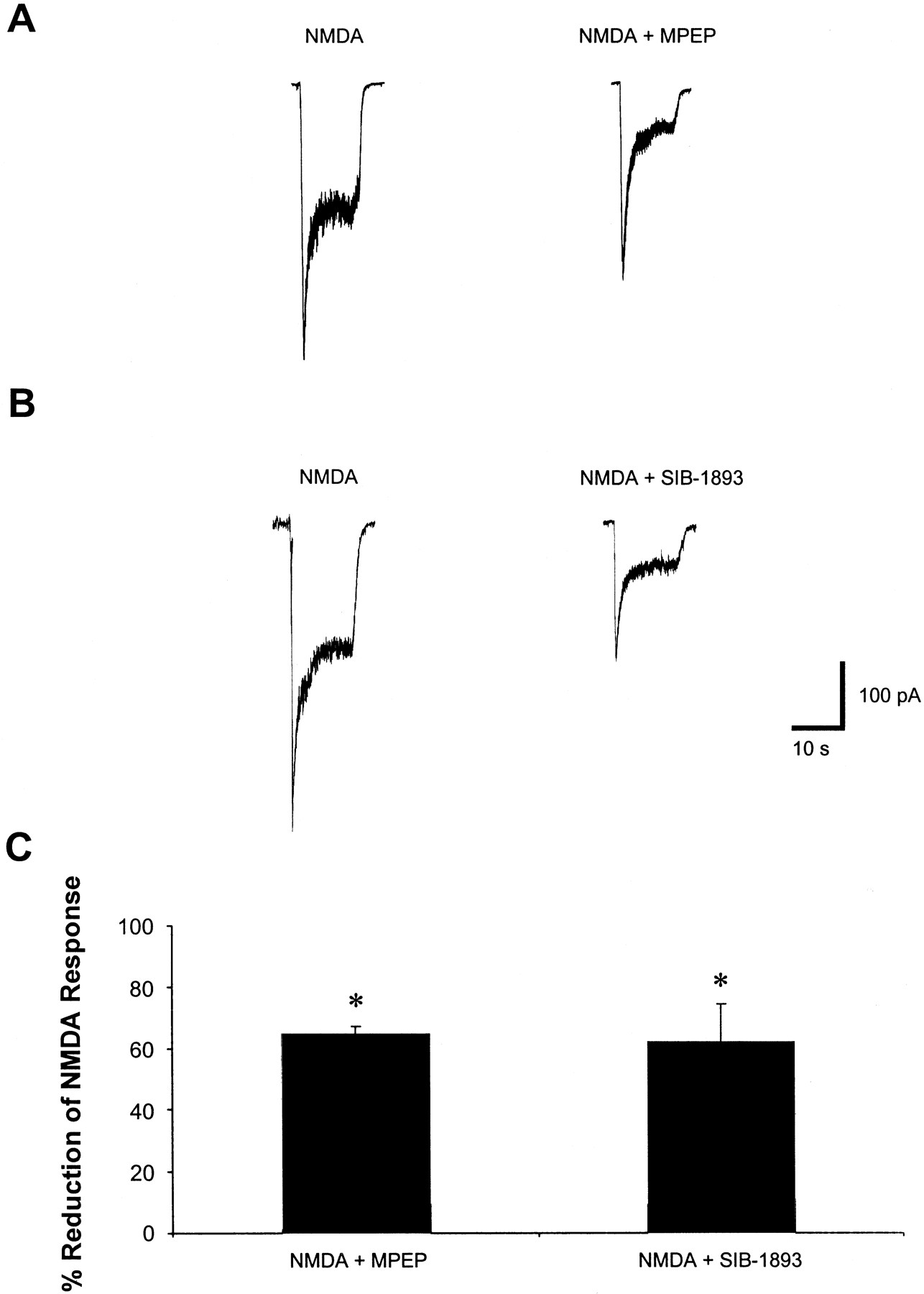
MPEP and SIB-1893 significantly reduce the whole-cell NMDA-evoked current response. A, representative traces of NMDA-evoked whole-cell current in the presence or absence of 200 μM MPEP. B, representative traces of NMDA-evoked whole-cell current in the presence or absence of 200 μM SIB-1893. C, summary of the effects of MPEP and SIB-1893 on NMDA-evoked current. Histograms represent percentage of NMDA-evoked current reduction ± S.E.M., n = 4. *P < 0.01 versus NMDA alone compared by Student's two-tailed t test.
To further investigate whether the neuroprotection caused by MPEP and SIB-1893 reflect action on NMDA receptor pathways, we examined the effects of these antagonists on traumatized neuronal cultures, with or without coadministration with the NMDA receptor antagonist MK801 (Bagetta et al., 1990; Ueda et al., 1995; Haghighi et al., 1996). Cotreatment of injured cultures with MK801 and MPEP or SIB-1893 showed no additional effects to those provided by MK801 alone (Fig.7A). We also examined the ability of the specific mGluR5 agonist CHPG to modulate the effects of MPEP and SIB-1893 on cell survival. Administration of CHPG (1 mM) to cortical neuronal cultures before treatment with the mGluR5 antagonists had no effect on MPEP or SIB-1893-mediated protection from glutamate-induced toxicity (Fig. 7B).

Coadministration of NMDA antagonist MK801 with MPEP or SIB-1893 have no additive effect on MK801-mediated reduction of injury-induced LDH release in rat cortical neuronal cultures. Cotreatment with the mGluR5 agonist CHPG did not affect the neuroprotection provided by MPEP. A, at 7 DIV, 20 μM MK801 together with 20 μM MPEP or SIB-1893 was added to cultures 30 min before mechanical injury. B, at 7 DIV, cultures were incubated 15 min with MPEP at indicated concentrations followed by 15 min of pretreatment with 1 mM CHPG before administration of 150 μM sodium glutamate. LDH release was measured after 24 h of treatment. Histograms represent LDH release as percentage of injured control ± S.D.,n = 8 to 16 cultures/condition. *P < 0.05 versus injured controls compared by ANOVA, followed by the Student-Newman-Keuls test. ▪, injured control; ■, CHPG; ▨, MPEP; ░, MPEP + CHPG.
Discussion
In the present study, we found that administration of mGluR5 antagonists MPEP or SIB-1893 significantly increased cell survival in cortical neuronal cultures after controlled mechanical trauma. The antagonists were effective in this model when given either as pretreatment or postinjury application. MPEP and SIB-1893 also showed marked protective effects against glutamate- or NMDA-induced toxicity in vitro. Given these results, we examined the effects of treatment with the mGluR5 antagonists in a well characterized model of TBI in rats. Traumatic injury in this system causes both necrotic and apoptotic cell death in ipsilateral cortex and apoptosis in ipsilateral hippocampus (Rink et al., 1995; Yakovlev et al., 1997). The model also shows well defined behavioral effects, both motor and cognitive, which can be modified by treatments that modulate either necrosis (Faden et al., 1989) or apoptosis (Yakovlev et al., 1997). Because MPEP and SIB-1893 share close structural homology (Gasparini et al., 1999) and were found to act similarly in the cell culture model, we used only MPEP for our in vivo study. Treatment with MPEP substantially attenuated motor dysfunction and significantly improved cognitive recovery after TBI. MPEP-treated animals also showed markedly reduced lesion volumes by MRI compared with vehicle-treated controls. MRI measurement of lesion volumes has been shown to reliably correlate with histological assessment of lesion size in other models of central nervous system injury (Hockings et al., 1995; Loubinoux et al., 1997), without the disadvantages of post-mortem artifacts caused by histological processing.
The ability of MPEP and SIB-1893 to function as mGluR5 antagonists via regulation of PI hydrolysis has been shown previously using mGluR5-transfected non-neuronal cell lines and rat brain tissue slices (Gasparini et al., 1999; Varney et al., 1999). We also examined the ability of MPEP to modulate PI hydrolysis in our cell culture system. Our results indicate that MPEP acts as an effective mGluR5 antagonist in rat cortical neuronal cultures, as shown by the complete inhibition of the phosphoinositide hydrolysis induced by the specific mGluR5 agonist CHPG. It should be noted that MPEP blocked agonist-induced PI hydrolysis at concentrations 100 times lower than that required for neuroprotection in cortical neuronal cultures. This finding suggests that neuroprotection provided by MPEP may not be related to mGluR5 modulation.
Moreover, other data from our study suggest that the neuroprotection observed with MPEP and SIB-1893 may have resulted from their ability to act as antagonists at NMDA receptors. As shown by our electrophysiological findings, both compounds significantly reduced the steady-state NMDA evoked whole-cell current at concentrations that provide neuroprotection in cultured cortical neuronal cells. These results were supported by cell viability assays that did not reveal additive neuroprotection when injured cells had been treated with the NMDA antagonist MK801 in combination with MPEP or SIB-1893. Such additive neuroprotective effects have been shown by us using the mGluR1 antagonist (RS)-1-aminoindan-1,5-dicarboxylic acid (AIDA) in combination with MK801 (J. Allen, S. Vicini, and A. I. Faden, unpublished observation). Moreover, pretreatment with the selective mGluR5 agonist CHPG did not reverse the neuroprotection provided by MPEP or SIB-1893 in cell cultures subjected to glutamate-induced toxicity. These findings are consistent with our previous observation that the reduction of neuronal cell death in injured cultures can be achieved by modulation of mGluR1, but not mGluR5 activity, using antisense oligonucleotides (Mukhin et al., 1996). Others, using the selective mGluR1 antagonists recently reported similar neuroprotective effects (Strasser et al., 1998;Pellegrini-Giampietro et al., 1999). Taken together, the presented data suggest that MPEP and SIB-1893 may have therapeutic potential in the treatment of TBI, although the mechanism of neuroprotective action for these drugs may to be linked to their modulation of NMDA receptor activity rather than to the inhibition of mGluR5. NMDA antagonists show considerable neuroprotective activity after traumatic injury in vivo and in vitro (Faden et al., 1989; Mukhin et al., 1997b). However, concerns have been raised about the possible side effects with such compounds, including neuronal vacuolization (Olney et al., 1989) and psychotropic action (Kornhuber and Weller, 1997). If compounds such as MPEP or SIB-1893 do not show such side effects, they may present distinct advantages as potential therapeutic agents in central nervous system injury.
Footnotes
-
Send reprint requests to: Alan I. Faden, M.D., Georgetown Institute for Cognitive and Computational Sciences, Department of Neuroscience, Georgetown University Medical Center, 3900 Reservoir Rd. N.W., Research Bldg., Rm. EP12, Washington, DC 20007. E-mail:fadena{at}giccs.georgetown.edu
-
This study was supported by grants from the National Institutes of Health (RO1NS37313) and the Department of Defense (DAMD-17-93-V-3018).
- Abbreviations:
- mGluR
- metabotropic glutamate receptor
- IP
- inositol phosphate(s)
- MPEP
- 2-methyl-6-(phenylethynyl)-pyridine
- SIB-1893
- (E)-2-methyl-6-(2-phenylethenyl)-pyridine
- TBI
- traumatic brain injury
- NMDA
- N-methyl-d-aspartate
- DIV
- day in vitro
- CHPG
- (R,S)-2-chloro-5-hydroxyphenylglycine
- LDH
- lactate dehydrogenase
- MRI
- magnetic resonance imaging
- PI
- phosphoinositide
- MK801
- (5R,10S)-(+)-5-methyl-10,11-dihydro-5H-dibenzo[a,d]cyclohepten-5,10-imine
- TR/TE
- repetition time/echo time
- RARE
- rapid acquisition with relaxation enhancement
- Received June 13, 2000.
- Accepted September 14, 2000.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}