


Abstract
Organic anion transporter 3 [Oat3(Slc22a8)] plays an important role in the renal handling of organic compounds. The substrate specificity of rat Oat3 and human Oat3 has been elucidated; information on mouse Oat3 (mOat3) is less defined. The aim of this study was to extend the substrate selectivity of mOat3. When expressed in Xenopus laevis oocytes, mOat3 mediated the uptake of p-aminohippuric acid and estron sulfate (ES). In addition to these substrates, we found that several organic compounds such as prostaglandin E2, prostaglandin F2α, allopurinol, 6-mercaptopurine (6-MP), 5-fluorouracil (5-FU), and l-carnitine are substrates of mOat3, compounds identified for the first time. The apparent Km values for the uptake of mOat3 that mediated the transport of 6-MP, 5-FU, and l-carnitine were 4.01 ± 0.7 μM, 53.9 ± 8.9 nM, and 61.9 ± 1.1 nM, respectively. Northern blot analysis revealed that gene coding for mOat3 is predominant in the kidney and, to a lesser extent, in the brain and the eye. Our findings thus provide further insights into the role of Oat3 in renal drug transport.
The kidney provides a pivotal route for the elimination and excretion of a wide range of structurally dissimilar compounds (Moller and Sheikh, 1983; Pritchard and Miller, 1993; Ullrich and Rumrich, 1993; Ullrich 1997). Renal excretion of organic compounds involves three steps: glomerular filtration, secretion, and reabsorption. The transepithelial active transport of ionic substrates in the kidney proximal tubules from the blood stream is carried out via several different types of multispecific organic ion transporters such as OATP/oatp1 (organic anion-transporting polypeptides), multidrug resistance-associated proteins, OCT (organic cation transporter) and Oat (organic anion transporter) (Koepsell, 1998; Russel et al., 2002; van Montfoort et al., 2003). These transporters are considered to be primarily involved in the transepithelial active transport of the kidney. Because the kidney is a major organ for the preservation of homeostasis and protects the body from toxic compounds, investigation of the substrate specificity of renal transporters is important in the drug discovery and clinical setting.
Regarding the Oat family, several isoforms, such as Oat1 (Slc22a6), Oat2 (Slc22a7), Oat3 (Slc22a8), Oat4 (Slc22a11), Oat5 (Slc22a10), and urate transporter 1 [URAT1 (Slc22a12)], which belong to the solute carrier superfamily 22a, have been isolated from different species (Simonson et al., 1994; Sekine et al., 1997; Sweet et al., 1997; Kusuhara et al., 1999; Race et al., 1999; Cha et al., 2000, 2001; Sun et al., 2001; Enomoto et al., 2002b). Functional analyses have revealed that the uptake of organic anions via Oat1 and Oat3 are trans-stimulated by glutarate, indicating that these two isoforms are an organic anion/dicarboxylate exchanger (Sekine et al., 1997; Sweet et al., 2003). In contrast, Oat2 and Oat4 do not exhibit such trans-stimulation (Sekine et al., 1998a; Cha et al., 2000). Thus, the functions of Oat are different among isoforms.
Oat3 was initially isolated from the rat and mouse kidney cDNA libraries (Brady et al., 1999; Kusuhara et al., 1999). Subsequently, hOat3 cDNA was isolated from the human kidney cDNA library (Cha et al., 2001). With regard to the substrate selectivity of rOat3, many investigators have reported that rOat3 mediates the transport of PAH, ES, ochratoxin A, cimetidine, benzylpenicillin, pravastatin, and uremic toxin indoxyl sulfate (Brady et al., 1999; Kusuhara et al., 1999; Deguchi et al., 2002; Hasegawa et al., 2002). Cha et al. (2001) have revealed that hOat3 mediates the transport of PAH, ES, ochratoxin A, methotrexate, cimetidine, dehydroepiandrosterone sulfate, glutarate, PGE2, PGF2α, cAMP, salicylate, and urate. All these findings indicate that Oat3 is an important molecule for the renal elimination of organic substrates from the body. However, only a few compounds such as PAH, ES, and taurocholate have been known to be substrates of mOat3 (Sweet et al., 2002).
To extend further the substrate specificity of mOat3, in the present study, we studied transport experiments using a Xenopus laevis oocyte expression system. We found that several organic compounds are novel substrates of mOat3. Our results provide further insights into the clinical and pharmacological relevance of Oat3 in renal drug transport.
Materials and Methods
Chemicals. [14C]PAH (1.50 GBq/mmol) and [3H]l-carnitine (2.22 TBq/mmol) were purchased from American Radiolabeled Chemicals (St. Louis, MO). [3H]PGE2 (7.429 TBq/mmol), [3H]PGF2α (7.943 TBq/mmol), [3H]5-FU (0.318 TBq/mmol), and [3H]ES (1.702 TBq/mmol) were purchased from PerkinElmer Life and Analytical Sciences (Boston, MA). [3H]Allopurinol (51 GBq/mmol) and [14C]6-MP (1.96 GBq/mmol) were purchased from Moravek Biochemicals (Brea, CA). Deoxycytidine [5′-α-32P]triphosphate (dCTP) (111 TBq/mmol) was obtained from Muromachi Yakuhin Kaisha, LTD (Tokyo, Japan). All other chemicals were of the highest grade commercially available.
Xenopus laevis Oocyte Preparation, cRNA Synthesis, and Transport Assays. Stage V and VI defolliculated oocytes were selected throughout this experiment. To remove the follicular layer from Xenopus oocytes, collagenase A (Roche Applied Science, Mannheim, Germany) was used at a final concentration of 2.0 mg/ml in OR2 (83 mM NaCl, 2 mM KCl, 1 mM MgCl2, 5 mM HEPES, pH 7.5) and slowly shaken for 1 h at room temperature. The mOat3 cDNA was linearized with BamHI, and the capped cRNA was transcribed in vitro by T7 RNA polymerase. Defolliculated oocytes were microinjected with 50 ng of in vitro transcribed cRNA under a stereomicroscope using a microdispenser (Drummond Scientific, Broomall, PA) and incubated for 2 days in a modified Barth's solution containing gentamicin (50 μg/ml) at 18°C. Uptake experiments of radiolabeled substrates, as indicated in each experiment, were performed in ND96 solution (96 mM NaCl, 2 mM KCl, 1.8 mM CaCl2, 1 mM MgCl2, 5 mM HEPES, pH 7.4) at room temperature. Oocytes were incubated in ND96 solution containing radiolabeled substrates for 1 h at room temperature. The uptake was terminated by the addition of 2 ml of ice-cold ND96 solution, and the oocytes were washed with the same solution at least five times. The oocytes were solubilized with 250 μl of 10% SDS, and accumulated radioactivity was determined with a liquid scintillation counter.
Kinetic Study. Concentration-dependent uptake experiments of [14C]6-MP, [3H]5-FU, and [3H]l-carnitine via mOat3 were performed with each compound at a final concentration range of 1, 5, 10, 20, and 60 μM; 1, 10, 20, 50, 75, and 100 nM; and 1, 10, 20, 50, 75, and 100 nM, respectively. The compounds were incubated with oocytes expressing mOAT3 for 1 h at room temperature, stopped with ice-cold ND96 solution, and washed five times as described above. Individual oocytes were transferred to scintillation vials and dissolved on 250 μl of 10% SDS. A scintillation cocktail was added, and radioactivity was counted. Counts in control (water-injected) oocytes were subtracted from the counts in cRNA-injected oocytes. Data are presented as mean ± S.E.M., except for kinetic constants for which the error represents the error of the fit. Km indicates the Michaelis-Menten constant (nano- or micromolar).
Inhibition Study. For the inhibition experiments, oocytes expressing mOat3 were incubated for 1 h in ND96 solution containing [14C]PAH in the presence or absence of various inhibitors at a final concentration of 1 mM. PAH, diclofenac, famotidine, ibuprofen, cimetidine, amantadine, enalapril, and tenoxicam were directly dissolved in ND96 solution from stock solution. These stock solutions of the inhibitors were prepared in dimethyl sulfoxide and diluted to a final concentration as described above. The final concentration of dimethyl sulfoxide in the assay medium did not exceed 1.0%.
Northern Blot Analysis. Total RNA was isolated from various male mouse tissues using the acid guanidine thiocyanate-phenol-chloroform extraction method (Chomczynski and Sacchi, 1987). Two micrograms of poly(A)+ RNA were purified from total RNA and loaded onto 1.0% agarose/formaldehyde gel. After electrophoresis, the nucleic acids were transferred onto a nylon membrane (Hybond N+; Amersham Biosciences AB, Uppsala, Sweden). The filter was hybridized at 42°C overnight in a hybridization solution (50% formamide) with a full-length cDNA of mOat3, which was randomly labeled with [32P]dCTP (Feinberg and Vogelstein, 1983). The filter was finally washed in 0.1× standard saline citrate/0.1% SDS at 42°C.
Statistical Analysis. Kinetic data from experiments measuring the uptake of radiolabeled substrates were fit to the Michaelis-Menten equation by nonlinear least-squares regression analysis with standard errors derived from these curves. Comparisons of data measuring initial rates of uptake of radiolabeled substrates in the presence and absence of inhibitors were performed by the unpaired Student's t test.
Results
Sweet et al. (2002) have demonstrated that mOat3 transports PAH, ES, and taurocholate. In accordance with their report, oocyte expressing mOat3 mediated the uptake of PAH and ES to about 16- and 5-fold greater than the control oocyte, respectively (Fig. 1). In addition to these observations, we found that mOat3 also mediated the high transport of [3H]PGE2, [3H]PGF2α, [3H]allopurinol, [14C]6-MP, [3H]5-FU, and [3H]l-carnitine. These findings indicate that these compounds are novel substrates for mOat3.

Uptake of various 14C- or 3H-labeled compounds by mOat3-expressing oocytes.
The uptake rates of radiolabeled compounds ([14C]PAH, 10 μM; [3H]ES, 50 nM; [3H]PGE2, 10 nM; [3H]PGF2α, 10 nM; [3H]allopurinol, 10 μM; [14C]6-MP, 5 μM; [3H]5-FU, 20 nM; [3H] l-carnitine, 20 nM) by the control or mOat3 expressed oocytes were measured for 1 h. Values are mean ± S.E.M. of 9∼22-oocyte determinations. The significance between control (water-injected) and mOat3-cRNA-injected oocytes was determined by the unpaired t test (*, p < 0.01). Other experimental conditions and details are described under Materials and Methods.
Based on these findings, the concentration dependence of the uptake of [14C]6-MP, [3H]5-FU, and [3H]l-carnitine via mOat3 was subsequently examined. As shown in Fig. 2, the mOat3-mediated uptake of 6-MP, 5-FU, and l-carnitine showed saturable kinetics and could be modeled by the Michaelis-Menten equation. Nonlinear regression analyses yielded Km values of 4.01 ± 0.7 μM, 53.9 ± 8.9 nM, and 61.9 ± 1.1 nM for 6-MP, 5-FU, and l-carnitine, respectively.

Concentration-dependent uptake of [14C]6-MP (A), [3H]5-FU (B), and [3H]l-carnitine (C) mediated by mOAT3.
The uptake experiments of 6-MP, 5-FU, and l-carnitine via mOat3 were performed with these compounds at final concentration ranges of 1, 5, 10, 20, and 60 μM; 1, 10, 20, 50, 75, and 100 nM; and 1, 10, 20, 50, 75, and 100 nM, respectively. The uptake rates of 6-MP, 5-FU, and l-carnitine by the control (water-injected) or mOat3-epressing oocytes for 1 h were measured at variable concentrations. The uptake was saturable with Km values of 4.01 ± 0.7 μM, 53.9 ± 8.9 nM, and 61.9 ± 1.1 nM for 6-MP, 5-FU, and l-carnitine, respectively, and fit to the Michaelis-Menten curve. Values are mean ± S.E.M. of 4∼11-oocyte determinations. The mOat3-mediated transport was determined by subtracting the transport velocity in control (water-injected) oocytes from that in mOat3-expressing oocytes. Other experimental conditions and details are described under Materials and Methods.
To examine further substrate specificity of mOat3, the inhibition of mOat3-mediated [14C]PAH uptake was next examined. The result is shown in Fig. 3. The inhibition of the mOat3-mediated transport of [14C]PAH exhibited a rank order of enalapril > tenoxicam > cimetidine > ibuprofen = famotidine > diclofenac = amantadine. Thus, mOat3, as well as rat and human Oat3, has a broad substrate specificity.
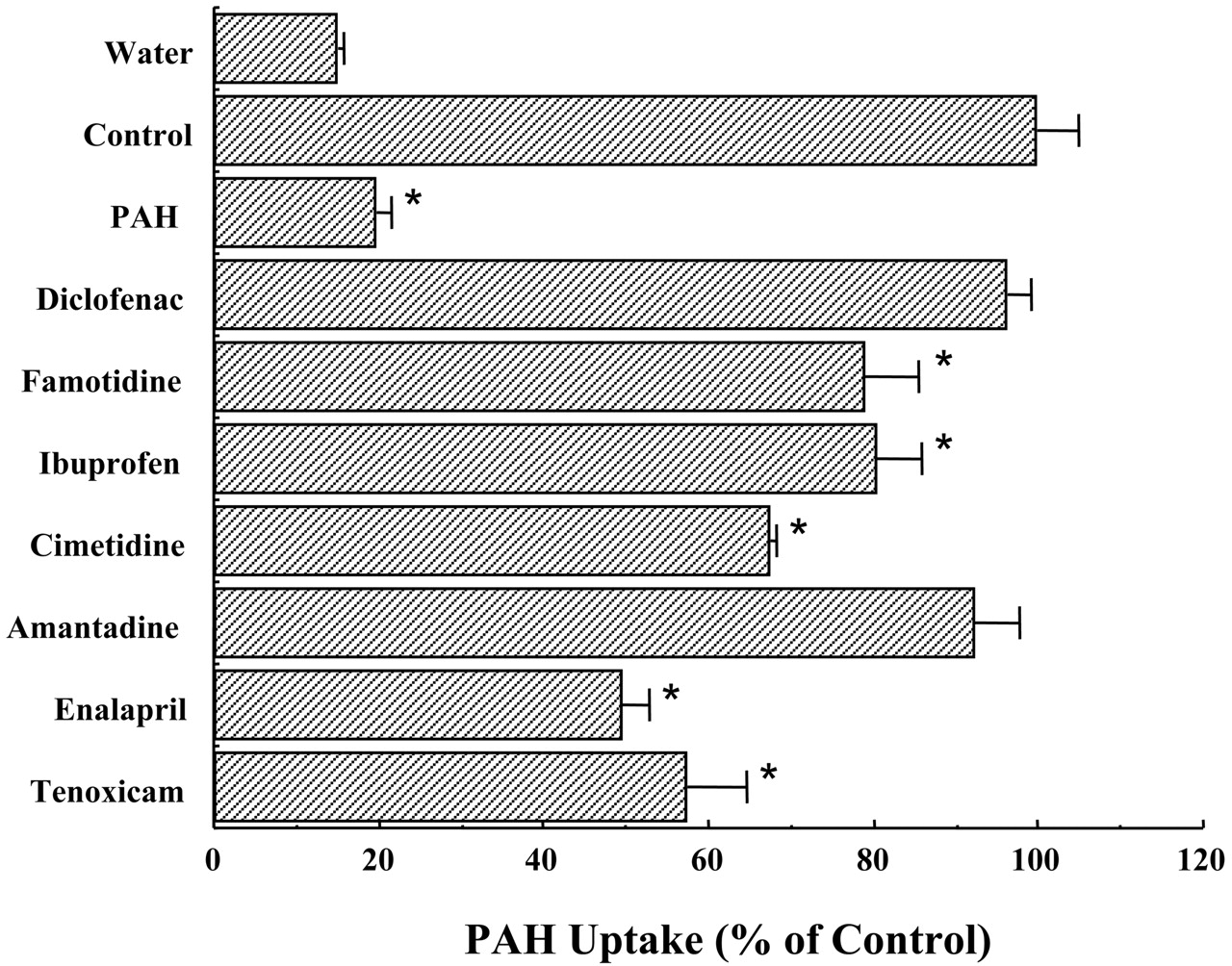
Inhibition of mOat3-mediated [14C]PAH uptake by various compounds.
The uptake rate of [14C]PAH by mOat3-expressing oocytes or control (water-injected) oocytes was determined in the presence or absence of 1 mM. The concentration of [14C]PAH was 10 μM. The values were expressed as a percentage of [14C]PAH uptake in mOat3-expressing oocytes in the absence of the inhibitor. Values are mean ± S.E.M. of 8∼15-oocyte determinations. The significance between control (water-injected) and mOat3-cRNA-injected oocytes was determined by the unpaired t test (*, p < 0.01). Other experimental conditions and details are described under Materials and Methods.
Brady et al. (1999) and Sweet et al. (2002) have shown that the mOat3 gene is expressed in the kidney. To elucidate further the distribution of the mOat3 gene, Northern blot analysis was examined. As shown in Fig. 4, a single mRNA band (2.7 kilobases) was pre-dominant in the kidney and weakly expressed in the brain and the eye. Hybridization signals could not be detected in mRNAs isolated from other tissues, such as the skeletal muscle, testis, pancreas, liver, heart, and lung. Thus, the expression of the mOat3 gene is tissue-specific.

Localization and tissue distribution of mOat3 mRNA by Northern blot analysis.
A high-stringency Northern blot analysis was made of poly(A)+ RNA from various tissues of the male mouse probed with 32P-labeled mOat3 cDNA. poly(A)+ RNA (2 μg) was loaded onto 1.0% agarose/formamide gel and transferred to a nylon membrane. A highly expressed 2.7-kilobase transcript was predominantly detected in the kidney and weakly expressed in the brain and the eye. Hybridization signals could not be detected in mRNA isolated from other tissues, such as the skeletal muscle, testis, pancreas, liver, heart, and lung. Other experimental conditions and details are described under Materials and Methods. kb, kilobase pairs.
Discussion
In the present study, we report the substrate specificity and tissue distribution of mOat3. Elucidation of the substrate specificity of drug transporters in the kidney is important to improve drug therapy and development. To date, several distinct drug transporter families such as the OATPs, OCTs, multidrug resistance-associated proteins, and Oats have been isolated. Among the Oat family, Oat3, as well as Oat1, is known to be a kidney predominant transporter (Brady et al., 1999; Kusuhara et al., 1999; Cha et al., 2001; Sweet et al., 2002). Functional analyses have revealed that rOat3 mediates the transport of PAH, ochratoxin A, cimetidine, benzylpenicillin, indoxyl sulfate, and ES (Kusuhara et al., 1999; Deguchi et al., 2002; Nagata et al., 2002). hOat3 mediates the transport of a wide range of organic compounds such as PAH, ochratoxin A, cimetidine, ES, taurocholate, methotrexate, estradiol-17β-glucuronide, dehydroepiandrosterone sulfate, glutarate, cAMP, salicylate, urate, and PGE2 (Cha et al., 2001). Thus, substrates for the Oat3 homolog partially overlap across the species. When expressed in Xenopus oocytes, mOat3 mediates the transport of PAH, PGE2, and ES, indicating that PAH, PGE2, and ES are conserved substrates of Oat3 across species.
In addition to these conserved substrates, we found several compounds (allopurinol, 6-MP, 5-FU, and l-carnitine) as mOat3 substrates. Allopurinol is known to be an inhibitor of xanthine oxidase and is useful in reducing the renal uric acid level in patients with recurrent or threatened renal deposition of urates, which occurs in gouty over-excretors of uric acid (Elion et al., 1966). On the other hand, 6-MP is converted to 6-thiourate by xanthine oxidase. It is well known that pharmacologic and toxic effects will occur with concomitant administration of allopurinol and 6-MP by inhibiting xanthine oxidase and resulting in an increase of the blood concentration of 6-MP (Elion et al., 1966). To elucidate a possible mOat3-mediated drug interaction between allopurinol and 6-MP, based on the uptake experiments, we performed the cis-inhibition experiments. Interestingly, the transport of 6-MP via mOat3 was completely inhibited by the addition of allopurinol (data not shown). Considering the membrane localization of mOat3 in the kidney proximal tubular cells (Sweet et al., 2002) and our uptake experiments, Oat3 may take part in the drug interaction between allopurinol and 6-MP and result in an increase of the blood concentration of 6-MP. Further studies (i.e., in vivo pharmacokinetic study) are required.
The antineoplastic 5-FU has been used for the treatment of several different types of malignancies, such as esophageal, gastrointestinal, ovarian, and breast carcinoma (Bonadonna and Valagussa, 1988; Jeung et al., 2001; Colleoni et al., 2002). The main mode of elimination of 5-FU is via renal glomerular filtration, but it is also eliminated via proximal tubule cells (Diasio and Harris, 1989). It has been reported that treatment with 5-FU results in the induction of proximal tubular cell necrosis (Kuriyama et al., 1984). However, as far as we know, there is no report concerning the molecular mechanism of 5-FU-induced nephrotoxicity. Although 5-FU possesses no anionic moieties, mOat3 mediates the transport of 5-FU, suggesting that Oat3 may, at least partly, participate in the development of 5-FU-induced nephrotoxicity.
l-Carnitine is a water-soluble vitamin and an essential component for mitochondrial β-oxidation of long-chain fatty acids (Bremer, 1983). Novel organic cation transporter 2/carnitine transporters 1 and 2 are localized at the luminal site of the renal proximal tubules and the transport of l-carnitine with high affinity (Sekine et al., 1998b; Tamai et al., 1998; Enomoto et al., 2002a). Taking the membrane localization and the transport experiments into consideration, mOat3 may play important roles in the uptake of l-carnitine to maintain the physiological concentration. Further detailed studies are needed.
Kusuhara et al. (1999) and Cha et al. (2001) have reported that transport of ES mediated by rOat3 and hOat3 is inhibited by chemically dissimilar compounds. Based on their findings, we further studied the inhibitory effect of the mOat3-mediated uptake by various organic compounds to obtain more detailed substrate selectivity. Because PAH is a conserved substrate across species (Kusuhara et al., 1999; Cha et al., 2001; Sweet et al., 2002), we chose PAH for this experiment. Although diclofenac is a potent inhibitor for the hOat3-mediated uptake of PAH, this compound had no inhibitory effects, indicating that these closely related proteins have functional differences. Thus, diclofenac may be a good tool for elucidating the differential substrate recognition between human and mouse Oat3.
It has been revealed that the gene coding for mOat3 is expressed in the kidney and the brain (Brady et al., 1999; Sweet et al., 2002). Kusuhara et al. (1999) and Cha et al. (2001) have demonstrated that rOat3 mRNA is expressed in the kidney, liver, eye, and brain, whereas hOat3 mRNA is localized in the kidney, skeletal muscle, and brain. Thus, tissue distribution of the Oat3 gene differs among species. To elucidate further a detailed distribution of the mOat3 gene, the expression and localization of mOat3 mRNA in various mouse tissues was studied with the use of Northern blot analysis. Interestingly, mOat3, as well as rOat3, was also weakly detected in the eye. Accordingly, our result indicates that expression of the Oat3 gene in the eye appears to be rodent-specific.
In conclusion, we demonstrate the substrate specificity and tissue distribution of mOat3. Our uptake experiments revealed that PAH, ES, and PGE2 are conserved substrates of rat, human, and mouse Oat3. Specifically, 6-MP, 5-FU, and l-carnitine were identified for the first time as mOat3 substrates. Furthermore, we found some differences in substrate specificity between mOat3 and hOat3. Our present findings provide further insight into the clinical and pharmacological role of Oat3 in renal drug transport.
Acknowledgments
We express special thanks to Dr. Ana M. Pajor, Department of Physiology and Biophysics, The University ofTexas Medical Branch at Galveston, Galveston, TX, for helpful suggestions for the manuscript.
Footnotes
-
↵1 Abbreviations used are: OATP, organic anion-transporting polypeptide; dCTP, deoxycytidine [5′-α-32P]triphosphate; ES, estron sulfate; 5-FU, 5-fluorouracil; hOat, human organic anion transporter; 6-MP, 6-mercaptopurine; mOat, mouse organic anion transporter; Oat, organic anion transporter; OCT, organic cation transporter; PAH, p-aminohippuric acid; PG, prostaglandin; rOat, rat organic anion transporter.
-
This work was supported in part by Grants-in-Aid from the Nakatomi Foundation (H13).
- Received November 21, 2003.
- Accepted January 16, 2004.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}