


Abstract
Concentrations of nicotine, cotinine, and nornicotine in brain and blood following both intermittent and continuous administration of [2′-14C]nicotine to rats were determined to assess nicotine metabolite accumulation in brain following repeated nicotine administration. For intermittent studies, rats were administered s.c. 1 to 10 doses of nicotine (0.3 mg/kg, 15 or 25 μCi of [2′-14C]nicotine; 30-min interinjection interval). For continuous administration studies, rats were implanted s.c. with an osmotic minipump delivering nicotine (0.8 mg/kg/day, 25 or 50 μCi of [2′-14C]nicotine for 1–21 days). Whole brain and trunk blood was collected. The concentration of [2′-14C]nicotine and its metabolites was determined via high-pressure liquid radiochromatography. Brain concentrations of nicotine, cotinine, and nornicotine increased 2-, 12-, and 9-fold, respectively, following 10 injections, reaching a plateau following the fifth injection. Brain blood ratios indicate an enhanced preferential distribution of nornicotine to brain with increasing numbers of injections. Across the 21-day period of continuous infusion, blood nicotine and nornicotine concentrations remained relatively constant, whereas concentrations in brain increased ∼4-fold. Generally, cotinine concentrations in brain and blood did not change across the infusion period. Brain/blood ratios indicate an increase in nicotine distribution into brain across days of nicotine infusion. Results demonstrate that both nicotine and its metabolites accumulate in brain following repeated nicotine administration, and indicate that brain nicotine concentration can not be extrapolated from plasma cotinine or nicotine concentrations. Thus, nornicotine accumulation following repeated nicotine administration suggests that this metabolite plays a contributory role in the neuropharmacological effects of nicotine.
Little is known about the metabolism of nicotine in human brain, although a plethora of information on peripheral nicotine metabolism exists (Kyerematen and Vesell, 1991; Gorrod and Wahren, 1993; Gorrod and Jacobs, 1999). Pharmacokinetic studies have determined the amount of nicotine in brain after various routes of administration in several animal species (Nordberg et al., 1989; Plowchalk et al., 1992). Nicotine metabolites have also been detected in the brain of animals peripherally administered radiolabeled nicotine (Appelgren et al., 1962; Schmiterlow et al., 1967; Stalhandske, 1970; Petersen et al., 1984). It is important to note that in the latter studies, N-demethylated nicotine metabolites such as nornicotine and norcotinine could not be detected by radiochromatography because the form of nicotine used contained a radiolabel in the N-methyl group.
Recently, extensive studies have been performed to identify and quantitate nicotine, cotinine, and N-demethylated nicotine metabolites in brain to determine their brain residence times following acute peripheral administration of [2′-14C]nicotine to rats (Crooks et al., 1995,1997; Crooks, 1999; Ghosheh et al., 1999). These studies have shown that, in addition to nicotine, the biotransformation products cotinine, nornicotine, norcotinine, and two minor unidentified metabolites appear in brain after a single dose of nicotine. Brain half-lives for cotinine, nornicotine, and norcotinine were 6, 3, and 4 times longer, respectively, than the half-life of 52 min determined for nicotine (Ghosheh et al., 1999), demonstrating that these biotransformation products have longer brain residence times compared with nicotine.
Intermittent inhalation during smoking affords peaks of nicotine superimposed upon a constant nicotine plasma level (Benowitz et al., 1991; Jacob and Benowitz, 1993). Considering the chronicity of nicotine exposure during tobacco smoking and the relatively longer brain residence time of the nicotine biotransformation products (Ghosheh et al., 1999), the possibility exists that after repeated peripheral nicotine administration, the metabolites of nicotine may accumulate in brain and reach pharmacologically significant concentrations. In this respect, cotinine and nornicotine have been demonstrated to have pharmacological activity in behavioral and neurochemical studies (Stolerman et al., 1995; Crooks and Dwoskin, 1997; Teng et al., 1997;Bardo et al., 1999; Dwoskin et al., 1999a,b). Thus, the accumulation in brain of these two slowly effluxing nicotine metabolites following repeated nicotine administration may contribute to the pharmacological effects resulting from nicotine exposure and possibly tobacco smoking.
Continuous drug administration affords equilibrium conditions between central and peripheral compartments and eliminates the peaks and troughs in circulating blood levels that occur with intermittent injections. This method of delivery has been used in animal models of chronic nicotine exposure and avoids stress as well as potential effects of conditioning due to repetitive injections (e.g., Marks et al., 1983; Sanderson et al., 1993; Rowell and Li, 1997). However, continuous nicotine infusion differs from the intermittent pattern of nicotine delivery during smoking (Benowitz et al., 1994).
The current study determined the distribution profile of nicotine, cotinine, and nornicotine in brain and blood following both intermittent and continuous administration of [2′-14C]nicotine to rats in order to test the hypothesis that these metabolites accumulate in brain following repeated nicotine administration.
Materials and Methods
Compounds.
(±)[2′-14C]Nicotine (free base, specific activity: 55 mCi/mmol; radiochemical purity was > 99%) was purchased from Moravek Biochemicals (Brea, CA). Altima AP scintillation cocktail was purchased from Packard BioSciences (Groningen, The Netherlands). S(−)-Nicotine tartrate was purchased from RBI/Sigma (Natick, MA). S(−)-Cotinine was purchased from Sigma Chemical Company (St. Louis, MO). HPLC1 grade triethylamine, acetonitrile, methanol, phosphoric acid, and sodium phosphate (monobasic and dibasic) were purchased from Fisher Scientific (Pittsburgh, PA). S(−)-Nornicotine and S(−)-norcotinine were prepared in our laboratory by previously described methods (Ravard and Crooks, 1996; Swango et al., 1998). Log P values for nicotine, nornicotine, and cotinine were calculated using CS ChemDraw7, version 4.5 (CambridgeSoft Corp., Cambridge, MA).
In Vivo Metabolic Experiments.
Male Sprague-Dawley rats (250–350 g) were obtained from Harlan Laboratories (Indianapolis, IN) and were housed two rats per cage with free access to food and water in the Division of Laboratory Animal Resources at the College of Pharmacy, University of Kentucky. Experimental protocols involving animals were approved by the Institutional Animal Care and Use Committee at the University of Kentucky. There was no attrition of subjects in any experimental condition.
Intermittent Nicotine Administration.
Groups of rats were administered either 1, 2, 5, or 10 doses of nicotine (0.3 mg/kg; free base equivalent containing either 15 or 25 μCi of [2′-14C]nicotine in saline; 30-min interinjection interval; 0.1 ml/100 g of body weight). The lesser amount of radiolabel was injected when either 5 or 10 doses were administered. The dose of nicotine was chosen as representative of that shown to be behaviorally active in rats (Clarke and Kumar, 1983; Ksir et al., 1985). The 30-min interval between nicotine injections was chosen to approximate the intermittent exposure to nicotine that the tobacco smoker experiences (Jacob and Benowitz, 1993). Rats were killed by rapid decapitation 30 min after the last injection. Whole brain was obtained within 2 min of decapitation. At least 3 ml of trunk blood was collected in centrifuge tubes containing 1 ml of an EDTA solution (10 mg/ml).
Continuous Nicotine Administration.
Nicotine was administered continuously by osmotic minipump to assess the kinetic parameters of nicotine uptake, accumulation into brain, and metabolism under equilibrium conditions. Continuous administration also approximates delivery via transdermal patch, i.v. infusion, or other continuous delivery devices. Rats were implanted s.c. with an osmotic minipump (Alza Corporation, Palo Alto, CA) delivering nicotine (0.8 mg/kg/day, free base equivalent; 50 μCi of [2′-14C]nicotine in 1-, 3-, and 7-day pumps, models 2001D, 2003D, 2007D, respectively; 25 μCi of [2′-14C]nicotine in 14- and 21-day pumps, models 2002 and 2004, respectively). Pumps were placed in phosphate buffer in a 37°C water bath for 4 h to equilibrate nicotine release before s.c. implantation. Before implantation, the rats were anesthetized with diethyl ether. Subsequently, an incision was made between the shoulder blades, the pump inserted, and the incision closed. Rats were killed by decapitation 1, 3, 7, 14, or 21 days after implantation of the appropriate pump, and whole brain and trunk blood collected. After removal of the pump, residual nicotine content was determined.
Preparation of Biological Samples.
The concentration of radiolabeled nicotine and its metabolites in brain was determined by homogenization of the entire brain in 3 volumes of ice-cold 1.15% w/v KCl using an Ultra-Turrax homogenizer (Janke & Kunkle, Staufen, Germany). The resulting homogenate was then centrifuged at 3000g for 30 min at 4°C (Sorvall RC 28S centrifuge, DuPont Company, Wilmington, DE). The supernatant was separated from the pellet and treated with 1 ml of 2% w/v ZnSO4 and incubated for 1 h at 34°C. The precipitated protein was centrifuged at 30,000g for 60 min at 4°C. The resulting supernatant was then analyzed by HPLRC.
The concentration of radiolabeled nicotine and its metabolites in blood was determined by initial centrifugation of collected blood samples at 1200g for 15 min at 4°C. The resulting supernatant (plasma) was separated and treated with 1 ml of 2% w/v ZnSO4. The resulting mixture was incubated for 1 h at 34°C. The precipitated protein was then centrifuged at 30,000g for 60 min at 4°C. The supernatant was collected and analyzed by HPLRC.
HPLRC Systems.
The HPLRC system comprised a Packard Series 1100b Radiomatic flow-through detector (Packard Instruments, Meriden, CT) connected to a Spectroflow 783 UV detector (ABI Analytical, Ramsey, NJ) operating at a fixed wavelength of 254 nm. A Beckman System Gold PSM 116 pump (Fullerton, CA) was used for solvent delivery. Column effluent was mixed in a 1:3 v/v with Altima flow AP scintillation cocktail before entering the Radiomatic detector. Samples were applied to the column (see below for column characteristics) using a Rheodyne model 7125 syringe loading injector via a 2-ml Rheodyne loop (Rheodyne Inc., Berkeley, CA). The outputs from the two detectors were recorded simultaneously on different channels of an Epson LX 810 printer (Torrance, CA), and were corrected for lag time.
A reversed phase HPLC system was used to quantify nornicotine, nicotine, and cotinine in rat brain supernatants. A cationic exchange HPLC system was used to quantify nicotine, cotinine, and nornicotine in rat blood supernatants as well as in brain supernatant. The reversed phase HPLC system used a Partisil-10 C8analytical column (4.6 × 250 mm, Whatman, Clifton, NJ) linked to a Partisil-10 C8 security-cartridge column (Phenomenex Co., Torrance, CA). The cationic exchange HPLC system used a Partisil-10 SCX analytical column (4.6 × 250 mm, Whatman) linked to a Partisil-10 SCX security-cartridge column (Phenomenex). Mobile phase buffers were prepared using HPLC grade reagents, and before use were degassed by filtering with a 0.45-mm × 47-mm filter (MSI, Westboro, MA). For both procedures, the flow rate was 1.5 ml/min. The mobile phase for the reversed phase system was 0.1 M sodium phosphate/methanol (92:8, v/v), containing 14 ml/l triethylamine (pH 4.8). The mobile phase for the cationic exchange system was 0.1 M sodium phosphate/acetonitrile (95:5, v/v), containing 30 ml/l triethylamine and adjusted to pH 7.0 with phosphoric acid.
Brain supernatants were coinjected with 100 μl of a standard solution containing 1.5 μg of authentic standards of S(−)-nicotine, S(−)-cotinine, S(−)-norcotinine, and S(−)-nornicotine. Radioactive metabolites eluting from the HPLC column were identified by comparing retention times with those of the UV-active authentic standards. Identities of the radiolabeled components were also confirmed using gas chromatography-mass spectroscopy and dual labeled experiments as described (Ghosheh et al., 1999).
Statistical Analysis.
Two-way mixed-factor ANOVAs were performed with number of injections and duration of administration as between-groups factors, and biological sample (brain and blood) as a within-subject factor. Where appropriate, Tukey's post hoc tests and simple main effect analyses were performed (p < 0.05).
Results
The concentration profiles of nicotine, cotinine, and nornicotine in brain and blood after 1 to 10 intermittent peripheral injections of nicotine (0.3 mg/kg, s.c., 30-min interinjection interval) are illustrated in Fig. 1. The results of the statistical analyses for the intermittent injection experiments are presented in Table 1. Profiles for nicotine and nornicotine in brain and blood were generally similar, such that brain concentrations increased with increasing number of nicotine doses, whereas blood concentrations appeared to remain relatively constant. In contrast, the profile for cotinine showed that both brain and blood concentrations increased with an increasing number of nicotine doses. Thus, the distribution profile for nicotine and its metabolites in brain and blood depended upon the number of nicotine injections administered.
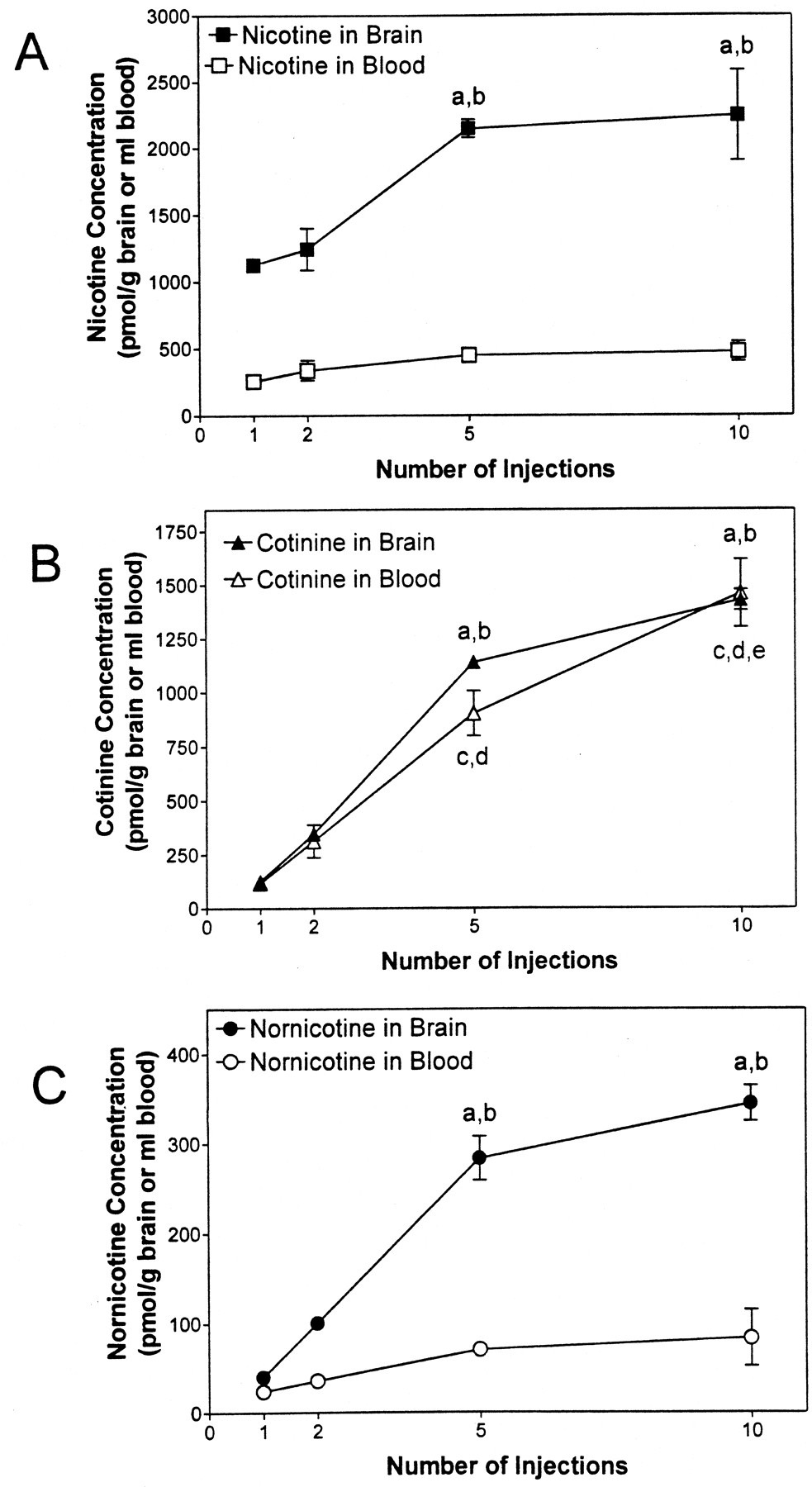
Concentrations of nicotine (A), cotinine (B), and nornicotine (C) in brain and blood after 1 to 10 peripheral injections of nicotine (0.3 mg/kg, s.c., 30-min interinjection interval).
a, p < 0.05, different from one injection for respective compound in brain; b, p < 0.05, different from two injections for respective compound in brain; c,p < 0.05, different from one injection for cotinine in blood; d, p < 0.05, different from two injections for cotinine in blood; e, p < 0.05, different from five injections for cotinine in blood.n = 3 rats/condition.
Summary of statistical analysis of concentrations of nicotine, cotinine, and nornicotine in brain and blood across 1 to 10 intermittent, peripheral [2′-14C]nicotine injections
Analysis of nicotine concentration in brain and blood revealed that nicotine distributed to brain and blood differently with increasing numbers of injections (Fig. 1A; Table 1). Across the injection sequence, nicotine concentration in brain was greater than that in blood (marginal means: 1692 ± 174 pmol/g of brain and 381 ± 36.0 pmol/ml of blood). Although there was a ∼2-fold increase in nicotine concentration in both brain and blood across the 10 nicotine injections, statistical significance was not obtained for blood, whereas a significant increase in brain nicotine concentration was found across the intermittent injection sequence (Fig. 1A; Table 1). Specifically, brain nicotine concentrations after 5 and 10 nicotine injections were greater than that after 1 and 2 injections. Importantly, the concentration of nicotine in brain after five injections was not different from that after 10 injections, indicating that nicotine concentration in brain reached a plateau. Thus, these results demonstrate that after intermittent injection of nicotine, the concentration of nicotine in brain is not reflected by its concentration in blood.
In contrast to nicotine, the results illustrated in Fig. 1B demonstrate that following intermittent nicotine injection, cotinine concentrations in blood accurately reflect cotinine concentrations in brain. Concentrations of cotinine in brain and blood after 5 and 10 nicotine injections were significantly greater (∼12-fold) than those following 1 and 2 injections (Fig. 1B; Table 1). Cotinine concentrations in blood following 10 injections of nicotine were significantly greater than those following 5 injections. In contrast, brain concentrations of cotinine following 5 and 10 injections were not different. Thus, although a similar cotinine distribution pattern is observed in brain and blood, the latter results indicate that a plateau was observed in brain, but not in blood.
The nornicotine distribution pattern in brain and blood was similar to that observed for nicotine. Nornicotine distributed to brain and blood differently with increasing numbers of injections (Fig. 1C; Table 1). Importantly, the concentration of nornicotine was greater in brain than in blood (marginal means: 192 ± 38.5 pmol/g of brain and 53.5 ± 10.1 pmol/ml of blood). Although the nornicotine concentration in blood increased ∼4-fold across the series of injections, a significant change was not found. In contrast to blood, the brain nornicotine concentration was greater (∼9-fold) after 5 and 10 nicotine injections than after 1 and 2 injections. Importantly, the concentration of nornicotine in brain after 5 injections was not different from that after 10 injections, indicating that a plateau was obtained. Thus, after intermittent nicotine administration, the concentration of nornicotine in brain is higher than its concentration in blood.
The brain/blood ratios for nicotine, cotinine, and nornicotine following 10 peripheral injections of nicotine are illustrated in Fig.2 (also see Table 1). The brain/blood ratio of nicotine (marginal mean: 4.6 ± 0.4) was different from that for cotinine (1.2 ± 0.1), but not different from that for nornicotine (4.0 ± 0.7). Thus, nicotine and nornicotine concentrations were ∼4 times greater in brain than blood, whereas brain and blood cotinine concentrations were not significantly different. These results indicate a preferential partitioning of nicotine and nornicotine from periphery into brain, whereas cotinine showed no preferential partitioning (Fig. 2). Although the brain/blood ratio for nicotine did not change across 10 nicotine injections, brain/blood ratios for nornicotine significantly increased (Fig. 2, Table 1), indicating enhanced distribution from the periphery into brain with increasing number of injections. Thus, there were no differences in brain/blood ratio across the number of injections for both nicotine and cotinine; however, there were differences in brain/blood ratio for nornicotine across the number of injections.
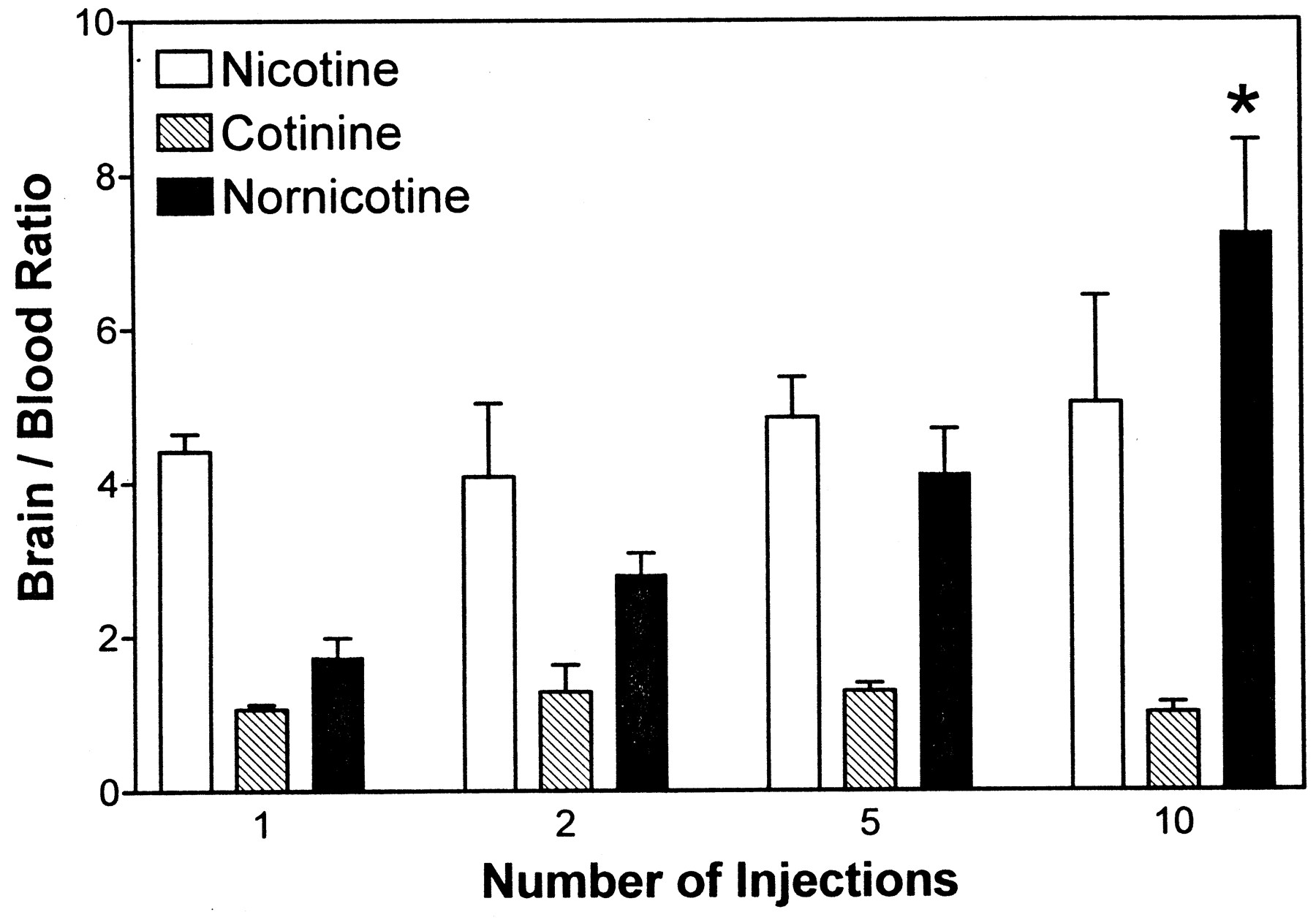
Brain/blood ratios for nicotine, cotinine, and nornicotine after 1 to 10 peripheral injections of nicotine (0.3 mg/kg, s.c., 30-min interinjection interval).
*p < 0.05, different from one and two injections for nornicotine. n = 3 rats/condition.
In a separate series of experiments, rats were implanted s.c. with an osmotic minipump, which continuously delivered nicotine at a dose of 0.8 mg/kg/day for up to 21 days. Figure 3illustrates the concentration versus time profiles for nicotine, cotinine, and nornicotine in brain and blood across the 21-day period of continuous nicotine infusion. The results of the statistical analyses for the continuous administration experiments are presented in Table 2. Generally, a greater concentration of nicotine and nornicotine was found in brain than in blood, whereas no difference between brain and blood concentration was apparent for cotinine.

Concentrations of nicotine (A), cotinine (B), and nornicotine (C) in brain and blood across a 21-day period of continuous infusion of nicotine (0.8 mg/kg/day via osmotic minipump).
a, p < 0.05, different from 1 day of infusion for respective compound in brain; b, p < 0.05, different from 3 days of infusion for respective compound in brain; c, p < 0.05, different from 7 days of infusion for respective compound in brain; *p < 0.05, different from 1, 3, 7, and 21 days of infusion for cotinine in respective biological sample; +p < 0.05 difference between brain and blood at 14-day infusion.n = 3 to 5 rats/condition.
Summary of statistical analysis of concentrations of nicotine, cotinine, and nornicotine in brain and blood across 1 to 21 days of continuous peripheral [2′-14C]nicotine administration
Surprisingly, nicotine concentrations in brain increased 4-fold across the 21 days of nicotine infusion, whereas blood nicotine concentrations did not change (Fig. 3A; Table 2). Although equilibrium was obtained for nicotine in blood, the concentration of nicotine in brain was directly dependent on the duration of continuous nicotine infusion. The brain nicotine concentration was significantly greater at 21 days than at 1 to 7 days, greater at 14 days than at 1 to 3 days, and greater at 7 days than at 1 to 3 days. Thus, brain concentrations of nicotine are higher than blood nicotine concentrations across the 21-day duration of continuous nicotine infusion.
Following continuous infusion of nicotine, cotinine concentrations in brain accurately reflected cotinine blood concentrations (Fig. 3B; Table 2). Although both cotinine brain and blood concentrations increased less than 2-fold at 14 days of continuous infusion compared with all other time points, brain cotinine was greater than blood cotinine only at the 14-day time point. Concentrations of cotinine in brain and blood were not different across the majority of the 21-day period of nicotine infusion. Thus, cotinine concentrations in blood are not different from those in brain, and equilibrium for cotinine between brain and blood was attained after a single day of continuous nicotine administration.
The concentration-time profile for nornicotine in brain and blood was similar to that for nicotine (Table 2). Figure 3C illustrates that nornicotine blood concentrations were not different across the 21-day period of infusion; however, brain concentrations of nornicotine increased 4-fold across the 21-day period. The brain nornicotine concentration was significantly greater at 21 days than at 1 to 7 days, and greater at 14 days than at 1 day of continuous administration. Thus, nornicotine brain concentration reached a plateau by 14 days, and the concentration of nornicotine in brain was higher than that in blood.
The brain/blood ratios for nicotine, cotinine, and nornicotine following 21 days of continuous nicotine infusion are illustrated in Fig. 4, and the results of the statistical analyses are presented in Table 2. The brain/blood ratio for nicotine was significantly greater than that for nornicotine and cotinine (marginal means: 5.2 ± 0.5, 2.1 ± 0.3, and 1.1 ± 0.1, respectively). The brain/blood ratios for nicotine and nornicotine increased ∼3-fold across the period of infusion, although statistical significance was only found for nicotine. The increase in the ratio primarily reflects an increase in concentration in the central compartment, since nicotine concentrations in blood were relatively constant across the infusion period (Fig. 2A; Table 2). In contrast, the cotinine brain/blood ratio (value of ∼1) was relatively constant across the 21-day infusion period. The latter results indicate that cotinine attained equilibrium by day 1, was equally distributed across central and peripheral compartments, and behaved differently from either nicotine or nornicotine.

Brain/blood ratios for nicotine, cotinine, and nornicotine across a 21-day period of continuous infusion with nicotine (0.8 mg/kg/day via osmotic minipump).
*p < 0.05, different from 1 day of infusion for nicotine. n = 3 to 5 rats/condition.
Discussion
An important pharmacokinetic characteristic of nicotine metabolites is that their brain residence times are significantly longer than that of nicotine (Ghosheh et al., 1999). The latter study reported the half-lives of cotinine and nornicotine to be 333 and 166 min, respectively, compared with 52 min for nicotine. Consequently, these metabolites have the potential to accumulate in rat brain to a larger extent than nicotine following repeated nicotine administration. Thus, prolonged, intermittent administration of nicotine, as is the case with chronic tobacco use, may result in pharmacologically relevant brain concentrations of these slow-effluxing metabolites. Considering the chronicity of nicotine exposure during tobacco smoking, the potential for accumulation of pharmacologically active nicotine metabolites is of importance for explaining the prolonged effects of tobacco smoking (e.g., tolerance and dependence). Metabolite accumulation could contribute to the central effects that result from peripheral nicotine exposure. Typically, a chronic smoker uses tobacco every 20 to 30 min with intermittent inhalation of mainstream smoke. Thus, delivery of nicotine to the brain during tobacco smoking constitutes intermittent peaks, superimposed on a constant plasma nicotine level (Benowitz et al., 1991; Jacob and Benowitz, 1993).
The current study determined the brain and blood concentrations of nicotine, cotinine, and nornicotine following acute, intermittent, and continuous peripheral administration of nicotine using an animal model. Results demonstrate that in addition to nicotine, cotinine and nornicotine accumulate in brain, and thus have the potential to contribute to the neuropharmacological effects observed after nicotine exposure.
Nicotine has been reported to accumulate in various rat tissues to afford concentrations 2-to 5-fold higher than those in plasma (Schmiterlow et al., 1967; Stalhandske, 1967; Tsujimoto et al., 1975). In the present study, peripheral intermittent injection of nicotine resulted in accumulation of nicotine, cotinine, and nornicotine in brain, which was evident after five injections. Brain concentrations of nicotine, cotinine, and nornicotine increased from 1.1, 0.1, and 0.04 μM, respectively, following acute nicotine administration to 2.2, 1.4, and 0.34 μM, respectively, after 10 injections, reaching a plateau following the fifth injection.
Importantly, in the intermittent nicotine injection study, cotinine concentrations in blood are an accurate reflection of cotinine concentrations in brain. Clinical studies have utilized cotinine plasma levels to indicate exposure to environmental tobacco smoke and prevalence of tobacco smoking (Wagenknecht et al., 1992; Benowitz, 1999). However, cotinine plasma levels only provide information on brain cotinine levels, and they provide only limited information concerning nicotine and its metabolites in peripheral and/or central compartments. The present results indicate that nicotine and nornicotine blood concentrations are clearly not related to their concentrations in brain after intermittent nicotine administration. In this respect, the minor peripheral metabolite, nornicotine, has been reported to constitute only ∼0.8% of peripherally metabolized nicotine in smokers (Benowitz et al., 1994). The relatively higher brain concentration of nornicotine is probably the result of a number of factors, including its longer brain residence time compared with that of nicotine, the number of nicotine injections, the interinjection interval, the preferential partitioning into brain (log P = 0.5), the potential involvement of an active transport process, and/or local metabolism from nicotine in brain.
Brain/blood ratios from the intermittent injection study reveal that the ratio for nicotine and cotinine is relatively constant over the injection schedule. Nicotine concentrations plateaued in both blood and brain after the fifth dose, indicating that equilibrium was attained between nicotine elimination and brain/blood distribution. Importantly, the brain/blood ratio for nicotine was ∼4, indicating that nicotine preferentially distributes from the periphery to the CNS compartment. A nicotine brain/blood ratio of ∼3 has been reported by Rowell and Li (1997) and Benowitz (1990), consistent with the present results. Cotinine, at a value of ∼1, was equally distributed to CNS and peripheral compartments. Importantly, the brain/blood ratio for nornicotine increased by ∼4-fold with increasing numbers of nicotine injections. Thus, the preferential distribution of nornicotine to the central compartment is accentuated with increasing numbers of doses, which may be due to involvement of active transport processes or metabolic factors. For example, as nicotine accumulates and increases in concentration in brain following repeated intermittent nicotine administration, it is possible that oxidativeN-demethylation to nornicotine occurs locally in brain in addition to nornicotine distribution to brain from the periphery. Compared with the liver, the brain is a poor metabolizing organ for nicotine biotransformation. However, cytochrome P450s 2El and 2B1 have been reported to increase in brain after 7 to 10 days of intermittent nicotine administration to rats (Anandatheerthavarada et al., 1993; Miksys et al., 2000). Thus, induction of nicotine N-demethylation in brain and liver and formation of nornicotine locally in brain would be expected to increase the total concentration of nornicotine in brain.
Continuous delivery of nicotine afforded cotinine brain and blood concentrations that were relatively constant across the entire 21-day period of administration. Although blood nicotine and nornicotine concentrations remained relatively constant, their concentrations in brain significantly increased (∼4-fold) across the period of infusion. Concentrations of nicotine, cotinine, and nornicotine following 1 day of continuous nicotine administration were 75, 137, and 10 nM, respectively, increasing to 274, 146, and 40 nM, respectively, after 21 days of continuous administration. Thus, in this steady-state condition wherein blood levels of all three components are relatively constant, nicotine and nornicotine continue to accumulate in brain with time.
Expression of the results as a brain/blood ratio revealed that cotinine had a value of ∼1, clearly indicating that this metabolite distributes equally in brain and periphery throughout the time course of continuous administration. Furthermore, this result is in agreement with previous work showing that after nicotine administration, cotinine had a partition coefficient of 1.28 between brain and arterial blood in the male rat (Gabrielsson and Gumbleton, 1993).
The brain/blood ratio for nicotine significantly increased from a value of 3 on day 1 to 7.5 on day 21. This is surprising, considering the equilibrium conditions and the short CNS half-life for nicotine compared with its metabolites. The brain/blood ratio for nornicotine also had a tendency to increase across the infusion period. One explanation for this result is recruitment or induction of active transport processes, leading to increases in nicotine and nornicotine in the CNS compartment. Another explanation is the presence of a barrier that prevents the exit of nornicotine and nicotine from brain. Furthermore, the physicochemical characteristics of nicotine and nornicotine, i.e., their basicity (pKa = 9.13 and 9.83, respectively) and lipophilicity (log P = 0.95 and 0.5, respectively) (Crooks, 1999) should also be considered. Alkaloids such as nicotine and nornicotine will likely be avidly sequestered into lipid-rich glia within the CNS compartment. Once compartmentalized, the slightly acidic pH (6.5–7.0) of the glial cytosol would lead to increased protonation of these molecules within this compartment compared with that in the extracellular compartment (pH 7.4). The increase in brain/blood ratio of either nicotine or nornicotine with time cannot be explained by this increase in protonation within the glial compartment, since this is an extremely rapid process. However, it is possible that partitioning of the neutral alkaloid species between extracellular fluid and glial cytosol is a time-dependent equilibrium process that can account for the increase in brain/blood ratio with time. In this respect, an equilibrium will be established between the protonated and unprotonated forms of the alkaloid in the extracellular fluid, and only the unprotonated species will permeate the glial membrane. Once the free base is present in glial cytosol, another equilibrium will be established between the protonated and unprotonated forms in this compartment. If the rate of permeation of the unprotonated species either into or out of the glia is a slow process (i.e., time-dependent), then the unprotonated form will accumulate in brain and an increase in brain/blood ratio of these two alkaloids will occur over time, constituting a barrier that impedes the exit of the alkaloid from brain into periphery.
The lack of change in the brain/blood ratio for cotinine across the 21-day period of continuous infusion is likely due to its lower basicity (pKa 4.27) and more polar nature (log P 0.04) compared with nicotine or nornicotine. Thus, distribution into the lipid-rich glial compartment will be reduced for cotinine, and protonation will not occur.
A less likely contributing factor may be up-regulation of high-affinity nicotine binding sites in brain following both intermittent and continuous nicotine administration (Marks et al., 1983; Rowell and Li, 1997). The doses used in the present studies were within the dose range reported to produce receptor up-regulation. However, since only a 20 to 30% increase in binding has been observed in the previous studies, up-regulation is probably only a minor contributor to nicotine and nornicotine accumulation in brain.
Although it is tempting to make direct comparisons between continuous infusion experiments and the intermittent injection experiments, such direct comparisons would be inappropriate because different single doses of nicotine were administered in the two series of experiments. To make such direct comparisons, a full nicotine dose response would be required using both types of administration protocols.
Of the two metabolites found in brain, nornicotine may contribute to the neuropharmacology of tobacco smoking, since it has the potential to accumulate in the CNS after both intermittent and continuous nicotine administration and because it possesses a pharmacology similar to that of nicotine. Nornicotine and cotinine evoke a concentration-dependent increase in dopamine release from rat striatal slices (EC50 ∼ 1 and 30 μM, respectively; Dwoskin et al., 1993, 1999b; Crooks et al., 1995; Teng et al., 1997). Similar to nicotine, nornicotine has reinforcing properties, as demonstrated by its maintenance of i.v. self-administration in rats (Bardo et al., 1999). Our study demonstrates that concentrations of nornicotine, but not cotinine, found in rat brain following intermittent administration of nicotine are within the range previously found to increase dopamine release in vitro, and this response may be responsible for the reinforcing properties of this alkaloid.
Both cotinine and nornicotine are major alkaloid components of tobacco (up to 20% of total alkaloidal content; Kisaki and Tamaki, 1961; Zhang et al., 1990). Therefore, chronic tobacco use may afford brain concentrations of these two metabolites/alkaloids that may be higher than those observed in the current experiments examining intermittent and continuous nicotine administration. Thus, a combination of alkaloidal nornicotine and metabolic nornicotine may be sufficient to produce brain concentrations of nornicotine that contribute to the neuropharmacological effects of tobacco smoking.
Acknowledgment
We gratefully acknowledge useful discussion with Dr. Patrick McNamara.
Footnotes
-
Send reprint requests to: Peter A. Crooks, Ph.D., College of Pharmacy, University of Kentucky, Lexington, KY 40536-0082. E-mail: pcrooks{at}pop.uky.edu
-
This research was supported by National Institutes of Health Grants DA 00399, DA 08656, and DA 06043 and a grant from the Tobacco and Health Research Institute, Lexington, KY.
- Abbreviations used are::
- HPLC
- high-performance liquid chromatography
- ANOVA
- analysis of variance
- CNS
- central nervous system
- HPLRC
- high-pressure liquid radiochromatography
- UV
- ultraviolet
- Received October 5, 2000.
- Accepted January 10, 2001.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}