


Abstract
The zonal uptake of estrone sulfate (E1S; 1 to 400 μM) was investigated in periportal and perivenous rat hepatocytes and cells isolated from whole liver (regular hepatocytes). Transport of E1S by periportal, perivenous, and regular hepatocytes was described by saturable (Kms of 24 to 26 μM and Vmaxs of 1.8 nmol/min/mg protein) and nonsaturable components (2.5 to 3.2 μl/min/mg protein) that were not different among the zonal regions (p > .05, ANOVA). These kinetic constants represented pooled values for the entire complement of transporters for E1S, including two known transporters of E1S: Ntcp, Na+-taurocholate cotransporting polypeptide, and oatp1, the organic anion transporting polypeptide cloned from rat liver. Uptake of E1S was significantly reduced by estradiol 17β-glucuronide (50 μM) and bumetanide (200 μM), and was inhibited strongly and competitively by pregnenolone sulfate with an inhibition constant of 6.7 μM. Further segregation of the kinetic constants as the sodium-dependent and -independent systems was achieved through simultaneous fitting of data obtained in the presence and absence of sodium from parallel hepatocytic uptake studies. For the periportal, perivenous, and regular hepatocytes, two saturable systems: a sodium-dependent transport system, characterized by similarVmaxs (1.1 to 1.4 nmol/min/mg protein) andKms (49 to 55 μM), a sodium-independent transport system of comparable Vmaxs (0.70 to 0.84 nmol/min/mg protein) and Kms (16 to 22 μM), and a linear clearance of 1.7 to 2.7 μl/min/mg protein (ANOVA, p > .05) were obtained. The data suggest that hepatic uptake of E1S involved sodium-dependent and -independent transporter systems. No heterogeneity in transport was observed.
Estrone sulfate (E1S)1serves as a storage form of estrogens in the human circulation and is used in hormone replacement therapy. It is metabolized primarily in liver to estrogens such as estrone, estradiol, and estriol. The influx of E1S into the liver could influence levels of estrogens in the body. Moreover, the presence of zonal distribution of transporters in the liver would affect the cellular concentration and processing of E1S by desulfation or biliary excretion among zonal cells, thus altering the overall hepatic clearance. This aspect has been shown to impact drug levels in simulation studies (Sato et al., 1986; Hansel and Morris, 1996; Kwon and Morris, 1997).
The hepatic transport of E1S is mediated by the organic anion transporting polypeptide (oatp1 and oatp2) and the sodium-dependent taurocholate cotransporting polypeptide (Ntcp), two recently cloned sinusoidal transporters (Hagenbuch et al., 1991, 1994;Jacquemin et al., 1994; Noé et al. 1997). oatp1 and oatp2, glycoproteins with 12 putative transmembrane domains, exist on the rat liver sinusoidal membrane (Jacquemin et al., 1994), the apical membrane of the kidney (Bergwerk et al., 1996), and the choroid plexus of the brain (Noé et al., 1997). The proteins are noted for their transport of bile acids and derivatives, the anionic dye bromosulfophthalein, ouabain, and anionic estrogen conjugates (Kullack-Ublick et al., 1994; Shi et al., 1995; Bossuyt et al., 1996;Kanai et al., 1996; Noé et al., 1997; Pang et al., 1998). oatp1-mediated transport is sodium independent, bidirectional (Shi et al., 1995), energy dependent (Shi et al., 1995), and possibly involves bicarbonate (Satlin et al., 1997) and glutathione (Li et al., 1998) as the counter ion. Ntcp, a glycoprotein with seven transmembrane domains, exists on the basolateral membrane of the liver. Ntcp preferentially mediates not only the hepatic uptake of bile acids and derivatives, but also anions such as E1S and bromosulfophthalein (Hagenbuch et al., 1991; Meier et al., 1997; Schroeder et al., 1998). The transport of anions such as the bile acid taurocholate by Ntcp is electrogenic and is driven by the physiological sodium gradient (Hagenbuch et al., 1991) with an apparent Na+-taurocholate stoichiometry of 2:1 (Weinman, 1997).
Although the nature of transport proteins is known for the uptake of E1S, there is virtually no information on its zonal uptake within the liver acinus. Heterogeneity in transport has been found for various endogenous and exogenous compounds. For example, the uptake of glutamate (Burger et al., 1989; Stoll et al., 1991), taurocholate (Stacey and Klaassen, 1981), ouabain (Stacey and Klaassen, 1981), and cysteine (Saiki et al., 1992) was reported to be greater in perivenous than in periportal hepatocytes, whereas the Na+/K+ ATPase activity was implicated to be lower in the perivenous region (Sillau et al., 1996). The intrinsic transport functions of oatp1 and Ntcp for E1S uptake has not been compared among zonal hepatocytes, although existing immunohistochemical evidence showed a lack of liver heterogeneity for Ntcp (Stieger et al., 1994) and for the mRNA of oatp1 in rat liver (Dubuisson et al., 1996). The objective of this study was to examine the heterogeneity in the uptake of E1S by investigating the intrinsic difference in transport velocity of E1S among periportal and perivenous hepatocytes, namely, for the sodium-dependent and -independent uptake of E1S. The uptake of E1S in the presence of pregnenolone sulfate, a structural analog, was appraised to study the possible interactions between the hormonal sulfate conjugates.
Experimental Procedures
Materials.
[3H]E1S [ammonium salt, specific activity (S.A.), 50 Ci/mmol], [14C]L-glutamic acid (S.A., 250 mCi/mmol), [14C]sucrose (S.A., 6.4 mCi/mmol), and [3H]sucrose (S.A., 11.9 Ci/mmol) were purchased from NEN Life Science Products (Boston, MA). Unlabeled E1S, glutamic acid, bumetanide, estradiol 17β-glucuronide, choline, BSA (Fraction V), and pregnenolone sulfate were obtained from Sigma Chemical Co. (St. Louis, MO). Collagenase A (Clostridium histolyticum) was purchased from Boehringer Mannheim (Darmstadt, Germany). Digitonin was obtained from Fluka Chemie (Buchs, Switzerland). Silicone oil (Fluids 510 and 550) was purchased from Dow Corning (Mississauga, ON). All other reagents and solvents were of HPLC grade.
Isolation of Rat Hepatocytes.
Male Sprague-Dawley rats (275–375 g, Charles River Canada, St. Constant, QC) were used for the preparation of isolated (regular) hepatocytes from whole liver by a previously established method (Hassen et al., 1996). Enriched periportal and perivenous hepatocytes were isolated by the digitonin/collagenase perfusion technique of Lindros and Pentı̈lla (1985), with modifications. After anesthesia (50 mg pentobarbital/kg, i.p.), the rat liver was perfused via the portal vein in a single- pass fashion at 25 ml/min for 10 min with a Ca2+- free buffer [consisting of Hanks buffer, 10 mM HEPES, 0.5 mM EGTA, 4.2 mM NaHCO3, 5 mM glucose, 0.65% BSA, pregassed with carbogen (95% O2, 5% CO2), and buffered to pH 7.2]. The medium was then changed to the digitonin solution (3.25 mM digitonin, 150 mM NaCl, 6.7 mM KCl, and 50 mM HEPES) as described by Tosh et al. (1996). The digitonin solution was delivered at a lower perfusion rate of 5.6 ml/min for approximately 35 ± 9 s (n = 8) progradely (flowing into the portal vein and exiting at the hepatic vein) or 77 ± 13 sec (n = 8) retrogradely (flowing into the hepatic vein and exiting at the portal vein). The infusion with digitonin was stopped immediately when maximal destruction of selective zonal cells was visually detected on the surface of the liver: appearance of white specks for destruction of perivenous cells and rings for destruction of periportal cells. The digitonin solution remaining in the liver was flushed out by perfusion with calcium-free buffer in the opposite direction (12 ml/min) for 2 min, then with collagenase buffer (Hanks buffer plus 4 mM CaCl2 and 0.06% w/w collagenase) for 8 min. All other subsequent steps involved were carried out as described previously (Hassen et al., 1996). Viability of the regular and zonal hepatocytes obtained was greater than 90%, as assessed by trypan blue exclusion.
Zonal Cells.
The enrichment of periportal and perivenous cells by the digitonin/collagenase perfusion method was monitored with alanine aminotransferase with a Sigma diagnostics kit and with glutamine synthetase assayed by a standard UV method (Meister, 1985); the protein content was measured by the method of Lowry et al. (1951). Because the uptake of glutamate from blood was shown to be predominantly perivenous (Burger et al., 1989; Stoll et al., 1991), the periportal and perivenous cells were further differentiated by their ability to transport glutamate. After preincubation at 37°C for 10 min, hepatocyte suspensions were added to a mixture of unlabeled glutamate, [14C]glutamate and [3H]sucrose (an extracellular marker) to result in glutamate concentrations of 1 to 200 μM in 1.6 × 106 cells/ml. Samples (100 μl) retrieved at 30, 60, 90, and 120 s were placed into 300-μl polyethylene microfuge tubes containing silicone oil (100 μl of density 1.02 g/ml) atop 50 μl of 3 N NaOH. The tubes were centrifuged immediately for rapid filtration of the cells through the silicone oil layer to sediment into the lower alkaline layer. The radioactivities of the incubation mixture, the supernatant (top layer), and the hepatocytes (residue) were quantified by liquid scintillation counting (model 6800, Beckman Canada, Mississauga, ON).
Uptake of E1S.
For all studies, cell suspensions of 2 × 106 cells/ml of the regular, periportal, or perivenous hepatocytes, preincubated at 37°C for 10 min, were added mixtures of E1S, [3H]E1S and [14C]sucrose (an extracellular marker) to result in E1S concentrations of 1 to 400 μM in 1.6 × 106 cells/ml. Samples (100 μl) were retrieved at 15, 30, 45, and 60 s for centrifugation as described above. The effect of estradiol 17β-glucuronide (50 μM) and the diuretic bumetanide (200 μM) on E1S (50 μM) uptake was also studied. The inhibitory effect of pregnenolone sulfate (0, 10, 25, and 100 μM) on the uptake of E1S (1 to 200 μM) was investigated in parallel incubations. Because 1% ethanol was used for dissolution of pregnenolone sulfate in these studies, cell viability and uptake of estrone sulfate in 0% and 1% ethanol were first compared. Cell viability in 1% ethanol remained greater than 90% for at least 15 min, as determined by trypan blue exclusion. The preservation of hepatocytic viability in 1% ethanol was also observed by Sawyer et al. (1994) who demonstrated that both the liver cell viability and respiratory function were not adversely affected by 1% ethanol over a period of 6 h. In another set of studies, parallel incubations were conducted in the presence or absence of sodium—choline chloride was substituted for sodium chloride and potassium bicarbonate for sodium bicarbonate.
Kinetic Analysis of E1S Uptake.
The linear portion of the plot of accumulated amount versus time yielded a positive intercept and a slope on regression of the data. The positive intercept that represents rapid and saturable binding of E1S to hepatocyte membrane had been described byHassen et al. (1996). The slope furnished the initial velocity of uptake (v).
Various mechanisms, ranging from simple saturable Michaelis-Menten (eq.1) to multiple components (eqs. 3 or 5) and presence of a nonsaturable system, denoted by Pdiff or the first-order clearance (see eqs. 2 and 4), were used to describe the uptake of estrone sulfate by rat hepatocytes.
For the inhibition study with pregnenolone sulfate, data on E1S transport in the presence of pregnenolone sulfate (0, 10, 25, and 100 μM) were force fit to eq. 6 or 7 which denoted competitive inhibition, or eq. 8 or 9, which defined noncompetitive inhibition.
Fitting.
The data were fitted with use of the software package SCIENTIST (version 2; MicroMath Scientific Software, Salt Lake City, UT) with the least-squares method. The optimized parameters are summarized in Tables1 to 4. The selection of the weighting scheme and goodness of fit were based on the coefficients of variation of the estimated parameters and residual plots.
Biochemical characterization of zonal hepatocytes with marker enzymes, alanine aminotransferase, and glutamine synthetase
Refinement of kinetic parameters for E1S uptake in presence and absence of sodium in parallel incubation studies in isolated regular, periportal, and perivenous rat hepatocytes, at 37°C (n = 4)4-a
Statistical Analysis.
All data were presented as the mean ± S.D. and the means were compared by use of ANOVA, with the level of significance set at .05. The Model Selection Criterion and the Akaike Information Criteria (Akaike, 1974; Ludden et al., 1994) were used to select the appropriate model equation(s).
Results
Biochemical Characterization of Zonal Hepatocytes.
The activities of the two marker enzymes, alanine aminotransferase and glutamine synthetase, in the periportal and perivenous hepatocytes are summarized in Table 1. Significant difference (p < .05, ANOVA) was observed among the periportal and perivenous hepatocytes.
The accumulation of [14C]glutamate was linear with time up to 2 min among all of the concentrations investigated. The initial velocities of glutamate uptake (v), calculated from regression of data between 0.5 to 2 min, were best described by eq. 2and were statistically different among periportal and perivenous hepatocytes (Fig. 1). Saturable uptake of different Kms (59 ± 24 and 136 ± 23 μM) and Vmaxs (9.4 ± 5.6 and 47 ± 5.7 nmol/min/mg protein) were obtained for [14C]glutamate uptake by periportal and perivenous hepatocytes (ANOVA, p < .05) and the rate of [14C]glutamate uptake was significantly greater in perivenous cells at all concentrations. However, the nonlinear component was similar (Pdiff of 0.02 ± 0.01 μl/min/mg versus 0.03 ± 0.003 μl/min/mg, ANOVA,p > .05).

Glutamate transport by isolated rat periportal and perivenous hepatocytes at 37°C (n = 3).
Initial uptake rate of glutamate, obtained from the slope upon linear regression of data, was linear for all concentrations over 2 min.
Uptake Kinetics of E1S.
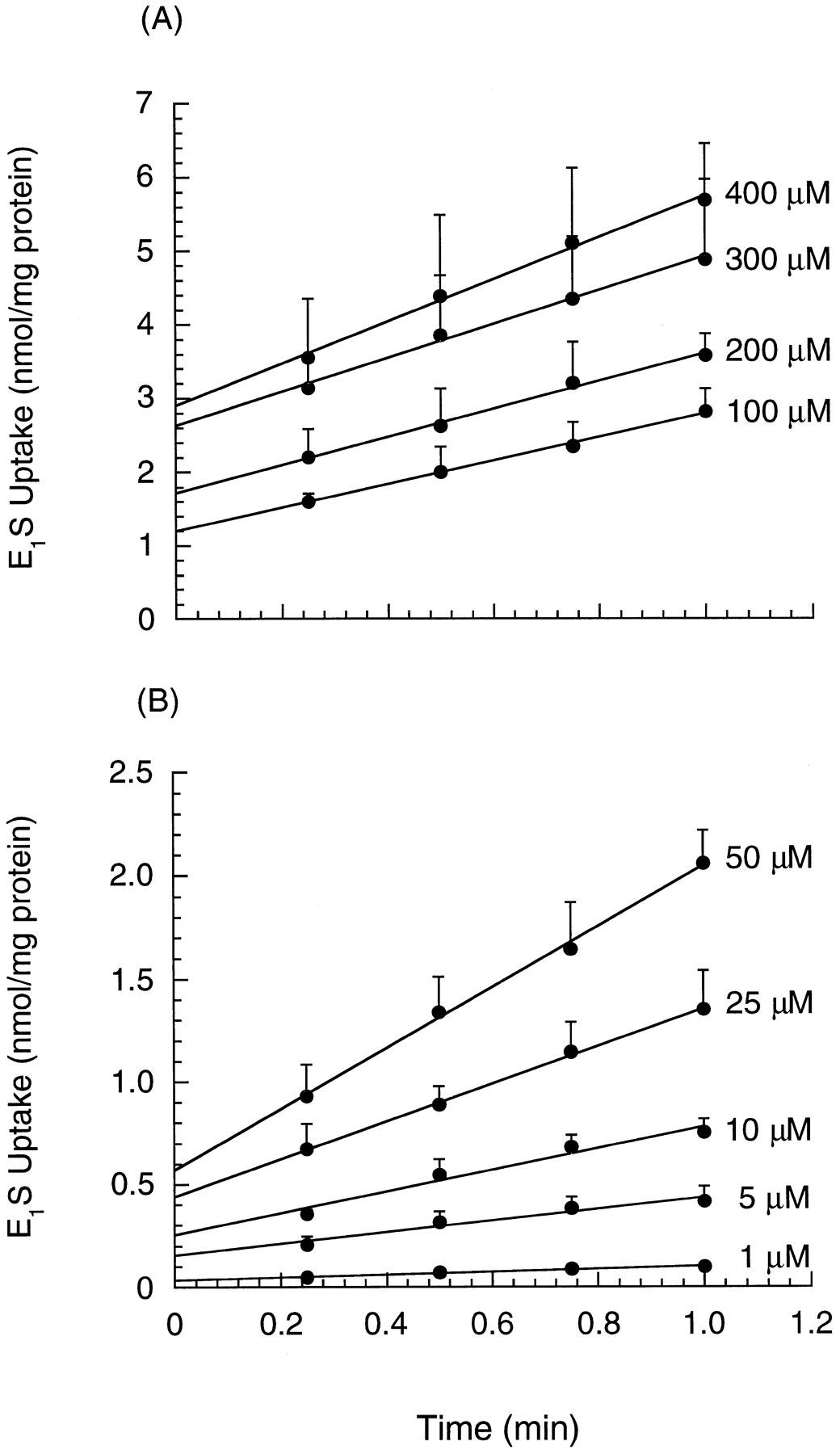
The time course for the uptake of E1S was shown in Fig. 2. Accumulation of E1S in hepatocytes remained linear with time within 1 min for all of the concentrations of E1S (1 to 400 μM). When the velocity was plotted against the E1S concentration, concentration-dependent kinetics became evident (Fig. 3A). Moreover, transport was temperature dependent and was reduced to a constant value (clearance of 2.1 ± 0.2 μl/min/mg protein) at 4°C. The data were best described by eq. 2, as found by Hassen et al. (1996), and a weighting scheme of 1/observation was optimal for fitting. The uptake of E1S was not significantly different among regular, periportal, and perivenous hepatocytes (Fig.3B; ANOVA, p > .05, table2). The Kms and Vmaxs varied from 24 to 26 μM and 1.8 nmol/min/mg protein, respectively, and the linear clearance,Pdiff, was 2.5 to 3.2 μl/min/mg protein. These kinetic constants were generally similar to those found by Hassen et al. (1996), but differed from those of Schwenk and del Pino (1980), who examined a much lower concentration range (0.1 to 10 μM) to arrive at lower estimates (Km of 0.8 μM and Vmax of 0.3 nmol/min/mg protein).

Time course of E1S uptake: 100 to 400 μM (A) and 1 to 50 μM (B) by isolated rat hepatocytes (regular or prepared from whole liver in absence of digitonin).
Points represent mean ± S.D. (n = 6).

Uptake rates (mean ± S.D.) for E1S by isolated rat hepatocytes: at 4°C and 37°C by hepatocytes isolated from whole liver (regular hepatocytes) (n = 6) (A), and in hepatocytes isolated from whole liver (n = 6), as well as in periportal (n = 8), and perivenous (n = 8) hepatocytes at 37°C (B).
Lines represent fitted lines based on eq. 2 with an optimal weighting scheme of 1/prediction.
Uptake kinetic parameters of E1S for isolated regular, periportal, and perivenous rat hepatocytes at 37°C2-a
Uptake of E1S in Presence of Inhibitors.
Previous studies had shown inhibition of E1S uptake by ouabain, energy depletors, and other sulfate conjugates (Hassen et al., 1996), as well as by taurocholate (Schwenk and del Pino, 1980). Presently, the coadministration of estradiol 17β-glucuronide (50 μM) and bumetanide (200 μM) was found to reduce E1S (50 μM) uptake significantly (to 66 ± 5.2% and 64 ± 3.5% of control, respectively,n = 4, p < .05). The uptake of E1S in the presence and absence of 1% ethanol was best described by eq. 2. Upon comparison, the saturable component of E1S transport in the presence and absence of 1% ethanol was similar, but the nonsaturable component of the uptake of E1S was slightly higher in the presence of ethanol (Fig. 4A and Table3). The reason for the difference was not apparent. When the effect of pregnenolone sulfate (0, 10, 25, and 100 μM dissolved in 1% ethanol) on E1S uptake was examined in parallel incubations, the extent of inhibition intensified with increasing concentration of pregnenolone sulfate. The data were best fitted to the competitive inhibition equation (eq. 5, Table4); the overall inhibition constant was 6.7 μM, a value that is less than the overallKm for E1S uptake (cf. values in Tables 2 and 3).

Uptake of E1S by isolated rat hepatocytes: in 0% (n = 6)and 1% ethanol (n = 4) in absence of pregnenolone sulfate (PS) (A), and in presence of 0, 10, 25, and 100 μM pregnenolone sulfate (PS) in 1% ethanol (n = 4), at 37°C (B).
Cell viability in 1% ethanol remained greater than 90% for at least 15 min. Lines in Fig. 4A represent best-fitted lines based on eq. 2, with an optimal weighting scheme of 1/prediction. All data points in Fig. 4B were simultaneously fitted to eq. 6 (competitive inhibition model) and an optimal weighting scheme of 1/prediction.
Uptake of E1S in presence of 0, 10, 25, and 100 μM pregnenolone sulfate (in 1% ethanol) by isolated rat hepatocytes
Uptake of E1S in Presence and Absence of Sodium.
The uptake parameters for E1S by rat hepatocytes were significantly reduced in absence of sodium (Fig.5A and Table 4, p < .05, ANOVA). Similar values as well as trends were observed for the data for periportal and perivenous hepatocytes in absence of sodium (Fig. 5B). The data were best fitted simultaneously to eqs. 4 and 2, which described two saturable systems and a nonsaturable component, instead of eqs. 5 and 3 due to the greater Model Selection Criterion, lower Akaike Information Criteria values, and the smaller coefficient of variation. These “refined” uptake parameters, summarized in Table4, revealed a slightly higher affinity (Kmof 16 to 22 μM) and slightly lower capacity (Vmax of 0.70 to 0.84 nmol/min/mg protein) system for the sodium-independent component, a lower affinity (Km of 49 to 55 μM) and slightly higher capacity (Vmax of 1.1 to 1.4 nmol/min/mg protein) system for the sodium-dependent component, and a linear component (Pdiff of 1.7 to 2.7 μl/min/mg protein).

Parallel incubations for study of uptake of E1S in presence and absence of sodium (choline and potassium substitution for sodium) in isolated rat hepatocytes: prepared from whole liver (regular hepatocytes) (n = 4) (A), and periportal and perivenous hepatocytes (n = 4), at 37°C (B).
Data obtained in presence and absence of sodium were force fit to eqs.4 and 2, respectively; a weighting of unity was optimal.
Discussion
The marker enzymes, alanine aminotransferase and glutamine synthetase, verified that enriched periportal and perivenous hepatocytes were isolated from different lobular origin within the rat liver. Our finding that alanine aminotransferase activity was higher in periportal hepatocytes (with a ratio of 1.9 for periportal to perivenous activities, Table 1) was in agreement with histochemical evidence on the enzyme in rat liver (Gorgens et al., 1988) and what was observed in other studies (Sillau et al., 1996; Tosh et al., 1996). The dramatic difference in glutamine synthetase activity between the perivenous and periportal regions (Table 1) was also shown by Stoll et al. (1991) and provided evidence on the successful preparation of enriched populations of periportal and perivenous cells. In addition, glutamate uptake, mediated by the sodium-dependent transporter, System G, in the perivenous region (Stoll et al., 1991) as well as the sodium-independent system in both periportal and perivenous hepatocytes (Burger et al., 1989; Stoll et al., 1991) was higher in perivenous hepatocytes (Fig. 1).
Although heterogeneity in transport was found for many endogenous and exogenous compounds: taurocholate (Stacey and Klaassen, 1981), ouabain (Stacey and Klaassen, 1981), and cysteine (Saiki et al., 1992), no significant difference in the uptake of E1S was found among the regular, periportal and perivenous hepatocytes. Among these systems, E1S uptake is described by a saturable and nonsaturable system of similar magnitudes (Table 2). The constancy in the values of Pdiff among the data sets in both the absence and presence of sodium suggests thatPdiff is most likely passive diffusion, because a relatively high diffusive component could be predicted for E1S in view of its high (1.4) octanol/water partition at pH 7.4 (our unpublished data). However, the discrepancy in the nonsaturable component of E1S uptake, Pdiff, in the presence of 1% ethanol may due to the fitting anomaly; the concentration used for this study was lower (up to 200 μM) and might not be high enough to fully reveal the linear component. In addition to the previously found inhibitors (Schwenk and del Pino, 1980; Hassen et al., 1996), the transport of E1S was reduced by estradiol 17β-glucuronide, a high-affinity substrate for oatp1 (Kanai et al., 1996), and bumetanide, a substrate for an organic anion transporter distinct from oatp1 and Ntcp (Horz et al., 1996), and was strongly inhibited by pregnenolone sulfate in a competitive fashion (Table 3 and Fig. 4).
Because oatp1, oatp2, and Ntcp mediate the transport of E1S in rat liver, it is reasonable to assume that data obtained with sodium-free media represent uptake by oatp1 and/or oatp2, the sodium-independent transporter, whereas data obtained in the presence of sodium encompass the entire spectrum of transporters in liver, including Ntcp. Thus, simultaneous fitting of data obtained in the presence and absence of sodium to eqs. 4 and 2, respectively, yielded the best model. The hepatocellular entry of E1S was best described by three components: a sodium-dependent component (saturable component), a sodium-independent component (saturable component), and a nonsaturable or linear component (Fig. 5 and Table 4). Furthermore, uptake of E1S by both sodium-dependent or -independent systems was not significantly different among the periportal and perivenous hepatocytes (Table 4). This finding was in agreement with immunohistochemical evidence for Ntcp (Stieger et al., 1994) and for oatp1 in isolated, zonal hepatocytes (T. N. Abu-Zahra, A.W. Wolkoff, and K. S. Pang, unpublished observations).
In this investigation, the Kms of the E1S transporters in rat hepatocytes were 14 and 47 μM, respectively, for the sodium-independent and -dependent system, suggesting that the sodium-independent pathway is of higher affinity. Indeed, transporters found in the rat liver, namely, oatp1 (Bossuyt et al., 1996), oatp2 (Noé et al., 1997), and Ntcp (Schroeder et al., 1998) expressed in the Xenopus laevisoocytes mediate the transport of E1S with corresponding Kms of 4.5, 11, and 27 μM, respectively, whereas the Km for the uptake of E1S uptake in COS cells expressing the human liver NTCP is 60 μM (Craddock et al., 1998). A lowKm for E1S was inferred in the inhibition of studies of E1S on estradiol 17β-glucuronide transport (Kanai et al., 1996). It appears that the Kms obtained from rat hepatocytes in this study correlate well with those obtained in other expression systems and that separation of the contributions of oatp1 and oatp2 for the transport of E1S was not feasible, because the Kms are close in values. The observation supports the view that, in absence of perturbation of the system and proper modeling, it becomes extremely difficult to separate the respective saturable components when theKms are similar (Sedman and Wagner, 1974). It is further interesting to note that the sum of theVmaxs of the two saturable systems (Table4) was similar to the overall Vmax obtained when data were treated as if it were a single saturable system (Table2). Analogously, the Kms of the two saturable systems (Table 4), when averaged, matched the overallKm obtained when data were treated as if it were a single saturable system (Table 2).
In conclusion, two saturable systems, a sodium-dependent and a sodium-independent system that most likely represent Ntcp and oatp, respectively, and a linear system, were found to mediate the transport of estrone sulfate. There was no difference in the uptake of E1S among regular, periportal, and perivenous hepatocytes. Pregnenolone sulfate was found to competitively inhibit the transport of E1S in rat liver.
Footnotes
-
Send reprint requests to: Dr. K.S. Pang, Faculty of Pharmacy, University of Toronto, 19 Russell St., Toronto, Ontario, Canada M5S 2S2. E-mail: pang{at}phm.utoronto.ca
-
This work was supported by the National Institutes of Health (Grant GM-38250) and Medical Research Council of Canada (MA-9104). Eugene Tan was a recipient of the Medical Research Council of Canada graduate fellowship.
- Abbreviations used are::
- E1S
- estrone sulfate
- oatp1 and oatp2
- organic anion transporting polypeptide 1 and 2
- Ntcp
- sodium-dependent taurocholate cotransporting polypeptide
- Received October 15, 1998.
- Accepted December 16, 1998.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}