


Abstract
The aim of this study was to quantitatively determine the constitutive expression levels of various transporter mRNAs in rat choroid plexus. To provide a reference for the relative expression levels, the expression of various transporter mRNAs in choroid plexus were compared with that in liver, kidney, and ileum. The mRNA levels of multidrug resistance protein (Mrp)1, 2, 3, 4, 5, and 6; multidrug resistance (Mdr)1a, 1b, and 2; organic anion transporting polypeptide (Oatp)1, 2, 3, 4, 5, 9, 12, and Oat-K (1/2); organic anion transporter (Oat)1, 2, and 3; organic cation transporter (Oct)1, 2, 3, N1, and N2; bile acid transporters sodium taurocholate cotransporting polypeptide (Ntcp), bile salt excretory protein (Bsep), and ileal bile acid transporter (Ibat); divalent metal transporter 1 (DMT1), Menke's and Wilson's metal transporters; equilibrative nucleotide transporters (Ent) 1 and 2, and constitutive nucleotide transporters (Cnt)1 and 2; peptide transporters (Pept)1 and 2; as well as ATP-binding cassette (Abc)G5 and 8 were measured in choroid plexus by the branched DNA signal amplification method. Mrp1, 4, and 5, Oatp3, Menke's transporter, DMT1, Ent1, and Pept2 mRNAs were expressed in choroid plexus at higher levels than in liver, kidney, or ileum. OctN1 and N2, Oatp2, Oat2 and 3, and Cnt1 and 2 mRNAs expressions were detectable in choroid plexus, but the levels were lower compared with that in liver, kidney, or ileum. The remaining transporters [Mrp2, Mrp3, Oct1, Oct2, Oatp1, Oatp4, Oatp5, Oatp12, Oat-K (1/2), Ntcp, Bsep, Ibat, Mdr1a, Mdr1b, Mdr2, Oat1, Ent2, Pept1, AbcG5, AbcG8] were expressed at very low levels in choroid plexus. The constitutive expression levels of different transporters in choroid plexus may provide an insight into the range of xenobiotics that can potentially be transported by the choroid plexus, thereby providing a means of xenobiotic detoxification in the brain.
The target-organ toxicity of any xenobiotic is dependent on its absorption, distribution, metabolism and excretion (ADME). The topic of xenobiotic metabolism has been the subject of intensive research for over 40 years, but research on various transport proteins that can mediate absorption, distribution, and excretion is a relatively recent undertaking.
Studies using rodent and human tissues have reported the existence of specific proteins that transport a variety of xenobiotics and endobiotics, such as drugs, flavonoids, metals, nucleotides and nucleosides, di- and tripeptides, and bile acids (Saito et al., 1995; Lomri et al., 1996; Miyamoto et al., 1996; Yao et al., 1997; Suzuki and Sugiyama, 1998; Cousins and McMahon, 2000; Ferguson et al., 2001; Li et al., 2001). The total number of transporters in rodents reported in the literature independent of the discoveries through the genome sequencing projects is about 50, and many more seem to await characterization. There are different classes of these transporters, such as organic anion transporters, organic cation transporters, ABC3 (ATP-binding cassette) transporters, metal transporters, peptide transporters, and nucleotide and nucleoside transporters, to name a few. Transporters of a particular class may have overlapping as well as unique substrate specificity. Each transporter is expressed in a tissue-specific and developmental stage-specific manner, and such expression may show a coordinate temporal regulation with that of various drug-metabolizing enzymes (Cherrington et al., 2002; Guo et al., 2002). A temporal integration of expression of specific transporters and drug-metabolizing enzymes that have similar substrate specificity can optimize the body's ability to cope with physiological and environmental chemicals.
The expression of various transporters in major tissues, such as liver, kidney, and intestine, has been studied by a number of investigators. However, comprehensive studies on the constitutive expression levels of various transporters in choroid plexus have not been undertaken previously. The choroid plexus is a highly vascularized tissue that lies in the ventricles of the brain. It consists of a single layer of cuboidal epithelial cells derived from the ependymal lining of the ventricles and overlying a vascular central core. The blood vessels and the connective tissue core of the choroid plexus are derived from a vascular fold of the pia mater termed the tela choroidea. The choroid plexus produces and secretes cerebrospinal fluid. The layer of cells abutting the capillary wall has complete access to the blood and its contents. Using materials brought into the circulation, the cells of the choroid plexus synthesize the components of the cerebrospinal fluid and secrete it into the lumen of the ventricles. The lining of the brain's ventricles and of the spinal cord's central canal is composed of ciliated cells, which circulate the cerebrospinal fluid. Choroid plexus plays an important role in regulating the exchange of various materials including heavy metals and small molecules, as well as many peptides, between the blood and the cerebrospinal fluid. This transport ability not only plays a crucial role in protecting the brain from various potential toxicological insults, but is also important in regulating the distribution of various physiologically and pharmacologically important compounds between the blood and the cerebrospinal fluid. Thus, choroid plexus is arguably one of the most toxicologically important tissues of the central nervous system. Therefore, various proteins and enzymes that play important roles in the detoxification of xenobiotics or protection from oxidative stress may be expressed at high levels in choroid plexus. Indeed, various antioxidant enzymes and enzymes involved in glutathione metabolism are known to be highly expressed in the choroid plexus. Additionally, high levels of glutathione and metallothioneins are also present in choroid plexus (Tayarani et al., 1989; Choudhuri et al., 1995; Cooper and Kristal, 1997).
The constitutive expression levels of different transporter mRNAs in choroid plexus may provide an insight into the range of xenobiotics that can potentially move in and out of the central nervous system and particularly in the brain of adult rats. Additionally, a detailed knowledge of the expression of various transporters in the choroid plexus may help identify previously unknown substrates that may reach the central nervous system. Therefore, the present work was undertaken in an effort to study the constitutive expression levels of various transporter mRNAs in rat choroid plexus in relation to the liver, kidney, and ileum. The expression of various transporter mRNAs has been determined using the highly specific and sensitive branched DNA (bDNA) signal amplification technique.
Throughout the text, the original nomenclature of transporters has been maintained, but in Table 1, the revised nomenclature has also been presented along with the original nomenclature to put the new transporter classification scheme in perspective.
Original and revised nomenclature of various transporters investigated in the present study
Materials and Methods
Isolation of Choroid Plexus. Brains from untreated male and female Sprague-Dawley rats (150-200 g; Charles River Laboratories, Inc., Wilmington, MA) were received from XenoTech LLC (Kansas City, KS). After decapitation, intact brains were rapidly collected, wrapped in aluminum foil, and kept in prechilled 50-ml tubes, which were flash-frozen in liquid nitrogen. This method of collection prevented shattering of the brains. Brains were transferred to -80°C until further use. For choroid plexus isolation, each brain was warmed under flowing water to rapidly thaw, quickly unwrapped from the aluminum foil, and sliced in an anteroposterior direction along the mid-region to obtain two intact cerebral hemispheres. Each cerebral hemisphere was immediately placed under a magnifying lens on a glass plate with its sliced side up. The choroid plexus was then quickly pulled out from the lateral ventricle like a thin thread with the help of a forcep having a nonserrated tip (to prevent tearing of the choroid plexus). The isolated choroid plexus was immediately flash-frozen in liquid nitrogen. In pilot studies, the identity and purity of choroid plexus had been confirmed histologically (data not shown).
Total RNA Isolation. Total RNA was isolated using RNAzol B reagent (Tel-Test Inc., Friendswood, TX) according to the manufacturer's protocol. Each RNA pellet was resuspended in 0.2 ml of 10 mM Tris-HCl buffer, pH 8.0. The concentration of total RNA in each sample was quantified spectrophotometrically at 260 nm. RNA integrity and quality were analyzed by formaldehyde-agarose (1.2% agarose, 2.1 M formaldehyde in 1× MOPS, ethidium bromide 0.5 μg/mg) gel electrophoresis. The quality of RNA samples was judged by the integrity and relative ratio of 28S and 18S rRNA bands.
Development of Specific Oligonucleotide Probe Sets for bDNA Analysis. The following probe sets were used: Mdr1a and 1b (Brady et al., 2002); Mdr 2 (Leazer and Klaassen, 2003); Mrp1, 2, and 3 (Cherrington et al., 2002); Mrp4, 5, and 6 (Leazer and Klaassen, 2003); Oatp1, 2, 3, 4, and 5 (Li et al., 2002); Oat-K(1/2), Oatp9 and 12 (Leazer and Klaassen, 2003); Oct1, 2, 3, N1, and N2 (Slitt et al., 2002); Oat1, 2, and 3 (Buist et al., 2002); divalent metal transporter 1 (DMT1) (Park et al., 2002); Wilson's and Menke's (Leazer and Klaassen, 2003); Ntcp and Bsep (Leazer and Klaassen, 2003); concentrative nucleoside transporter (Cnt)1 and 2 as well as Ent1 and 2 (Leazer and Klaassen, 2003); Pept1 and 2 (Leazer and Klaassen, 2003); and AbcG5 and 8 (Leazer and Klaassen, 2003). The remaining sequence [ileal bile acid transporter (Ibat)] was obtained from GenBank (Table 1). These target sequences were analyzed by ProbeDesigner software version 1.0 (Genospectra, Fremont, CA). All oligonucleotide probes were designed with a melting temperature (Tm) of approximately 63°C, enabling hybridization conditions to be held constant; that is, hybridization at 53°C for each oligonucleotide probe set. Every probe developed through the ProbeDesigner software was BLAST-searched against the nucleotide database to ensure minimal or no cross-reactivity with other known rat sequences and expressed sequence tags (ESTs).
bDNA Signal Amplification Assay. Reagents required for RNA analysis (i.e., lysis buffer, amplifier/label probe buffer, and substrate solution) were supplied in the HV-QuantiGene bDNA signal amplification kit (Bayer Corp.-Diagnostics Division, Tarrytown, NY). Expression levels of transporter mRNAs were analyzed as described by Hartley and Klaassen (2000). Briefly, specific oligonucleotide probe sets were diluted in lysis buffer. Total RNA (1 μg/μl; 3 μl total volume) was added to each well of a 96-well plate containing 50 μl of capture hybridization buffer [0.05 M HEPES sodium salt, 0.05 M HEPES free acid, 0.037 M lithium lauryl sulfate, 0.5% (v/v) Micr-O-protect, 8 mM EDTA, 0.3% (w/v) nucleic acid blocking agent] and 50 μl of diluted probe set. Total RNA was allowed to hybridize to each probe set containing all probes for a given transcript (blocker probes, capture probes, and label probes) overnight at 53°C. Subsequent hybridization and posthybridization wash steps were carried out according to the manufacturer's directions, and luminescence was measured with the Quantiplex 320 bDNA Luminometer (Bayer Corp.-Diagnostics Division) interfaced with Quantiplex Data Management software version 5.02 (Bayer Corp.-Diagnostics Division) for analysis of luminescence from 96-well plates.
Statistical Analysis. All values are expressed as relative luminescence units (RLU) per 3 μg of total RNA with mean ± S.E.M. for an n of 4 pools, each from approximately 60 rats. Two choroid plexus pools were taken from female tissues only, whereas the third pool was from male choroid plexus. The fourth pool was a mixture of male and female tissues. Liver, kidney, and ileum samples were taken from individual male rats.
Results
Pilot studies were conducted using total RNA from choroid plexus (CP) of male and female rats. These studies did not indicate any major gender difference in the expression of transporters (data not shown). Thus, the results of the final study presented here represent an average of four independent experiments conducted using total RNA from the CP of males as well as females (see statistical analysis for details). The constitutive expression levels of various transporter mRNAs in the CP have been compared with that in liver, kidney, and ileum.
Figure 1 shows that mRNAs for multidrug resistance proteins 1, 4, and 5 (Mrp1, 4, and 5) had the highest constitutive expression in CP, followed by that in kidney, ileum and liver, respectively. Quantitatively, the expression of Mrp1, Mrp4, and Mrp5 in CP was about 11-, 1.3- and 4-fold greater than in kidney, but it was 107-, 35-, and 13-fold greater, respectively, than in liver. In contrast, Mrp6 mRNA expression in CP was about 30% of that in liver, where its expression was the highest. The expression of Mrp2 and 3 mRNAs in CP was very low; it was the highest in liver, followed by ileum. Quantitatively, the expression of Mrp 2 in CP was about 7% of that in liver, and the expression of Mrp3 was about 8.5% of that in ileum.

Expression of different forms of Mrp mRNA in CP, liver, kidney, and ileum.
Mrp1, Mrp4, and Mrp5 showed the highest expression in CP.
The mRNA expression of multidrug resistance 1a, 1b, and 2 (Mdr1a, 1b, 2) was very low in CP. Mdr1a and Mdr1b expressions were highest in ileum, and Mdr2 expression was highest in liver. Mdr1a and Mdr1b expressions in CP were about 1.8% and 4.4%, respectively, of that in ileum, whereas Mdr2 expression in CP was about 6.7% of that in liver (Fig. 2).
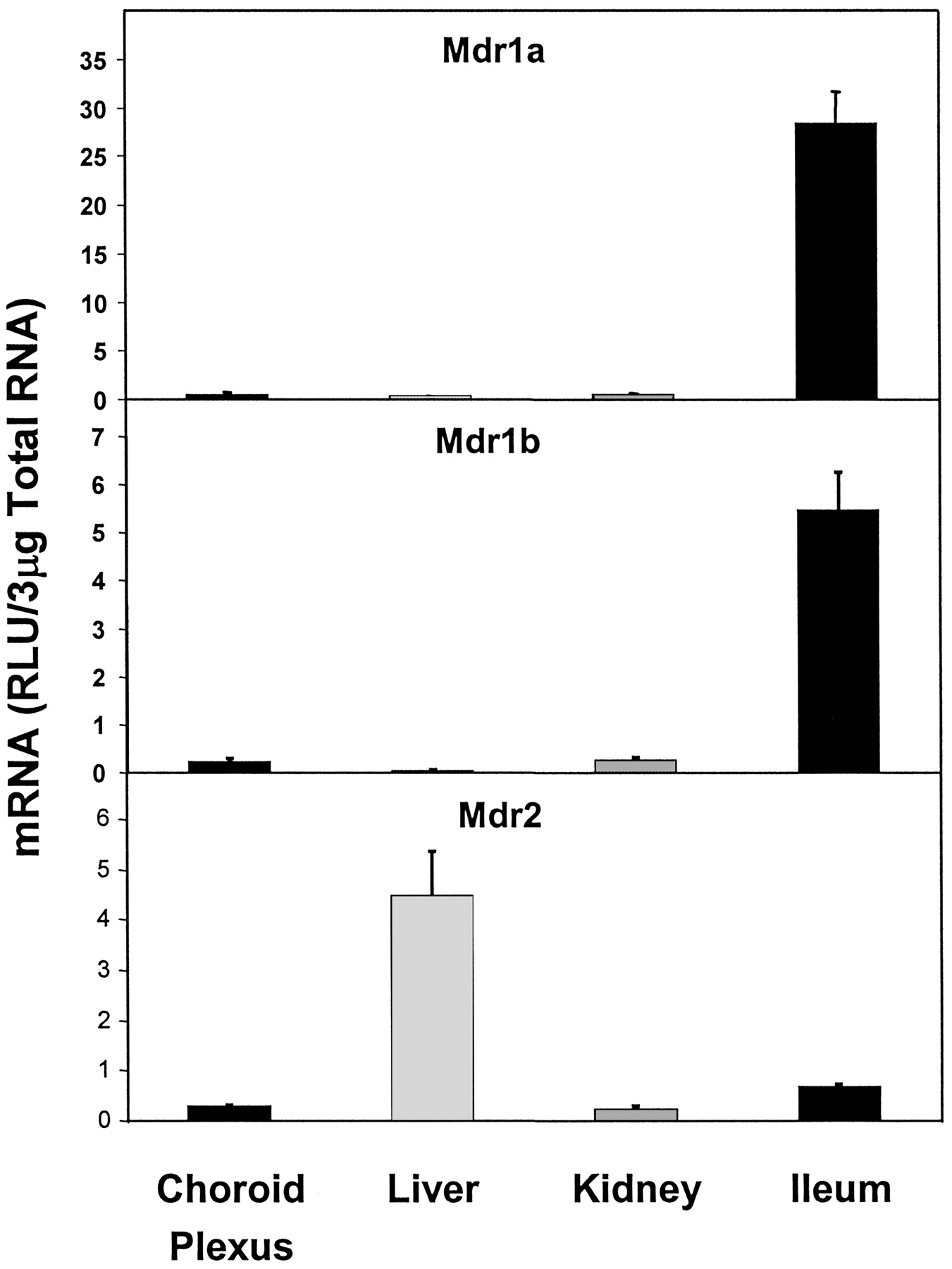
Expression of different forms of Mdr mRNA in CP, liver, kidney, and ileum.
Mdr1a, Mdr1b, and Mdr2 all showed very low-level expression in CP.
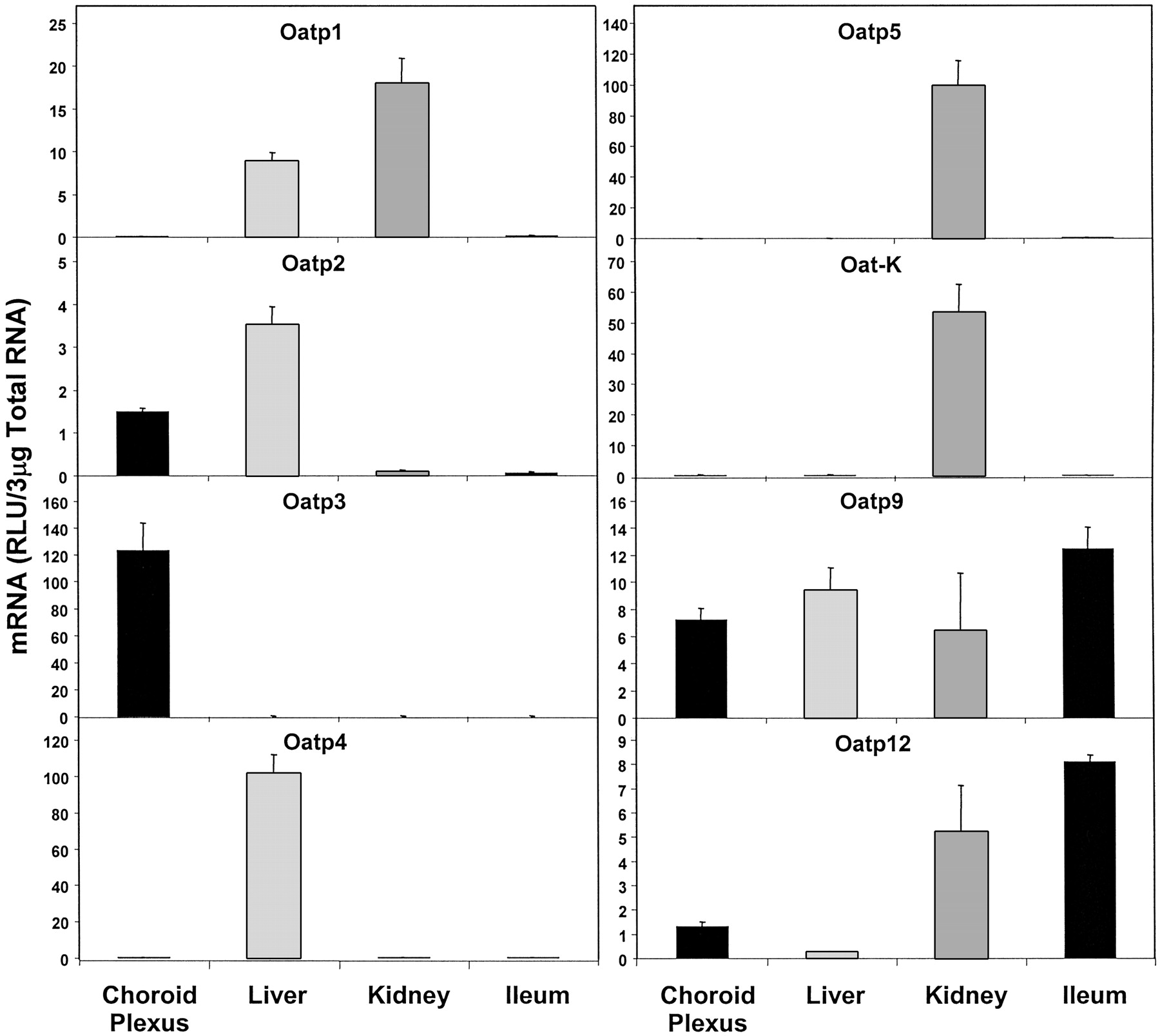
Figure 3 shows the expression of organic anion-transporting polypeptide (Oatp) mRNAs. Oatp3 mRNA was found to be highly expressed in CP and was quantitatively more than 500-fold greater than in liver, kidney, and ileum (mean RLU in choroid plexus was about 123; mean RLU in liver, kidney, and ileum was about 0.22). Constitutive expression of Oatp2 mRNA in CP was about 43% of that in liver, where its expression was the highest. The expression of Oatp2 in kidney and ileum was almost negligible compared with that in liver or CP. The expression of Oatp9 mRNA in CP was similar to that in liver. The expression of Oatp12 mRNA was about 15% of that of ileum, the organ with highest expression of Oatp12. In contrast, the expression of Oatp1, 4, 5, and Oat-K mRNAs was virtually nonexistent in CP as evident from the RLU values (less than 1% in CP as compared with that in the organ with the highest expression).

Expression of different forms of Oatp mRNA and Oat-K (1/2) in CP, liver, kidney and ileum.
Oatp3 was found to express almost exclusively in CP, whereas Oatp4, Oatp5 and Oat-K (1/2) showed no expression in CP.
Figure 4 illustrates the expression of organic anion transporter (Oat) mRNAs. Both Oat2 and Oat3 were highly expressed in CP, in relation to other tissues examined. Oat2 mRNA expression in CP was about 53% of that in liver, the organ with highest Oat2 expression. Oat3 mRNA expression in CP was about 50% of that in kidney, the organ with the highest Oat3 expression. Oat1 mRNA expression in CP, however, was virtually nonexistent, being about 1.8% of that in kidney, where its expression was the highest.
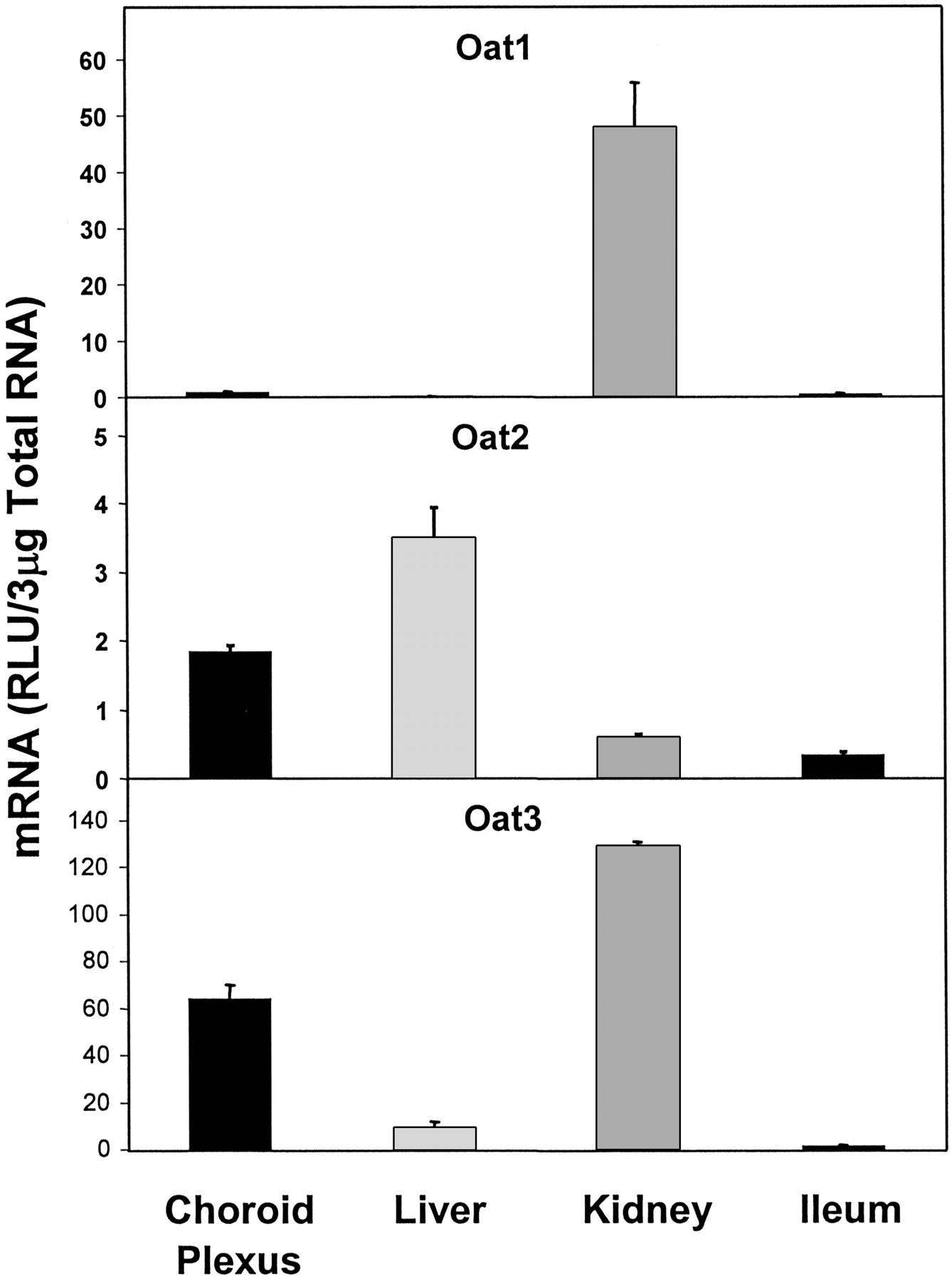
Expression of different forms of organic anion transporter (Oat1, Oat2, Oat3) mRNA in CP, liver, kidney, and ileum.
Oat1 had almost no expression in CP.
Organic cation transporter 3, N1, and N2 (Oct3, N1, N2) mRNAs were all expressed in CP. Oct3 expression was highest in ileum, and in CP it was about 41% of that in ileum. The expressions of OctN1 and OctN2 were highest in kidney, and in CP they were about 67% and 31% of that in kidney, respectively. The expression of Oct1 in CP was about 3.5% of that in kidney, the organ with the highest Oct1 expression. Oct2 expression was virtually nonexistent in CP, as evident from the RLU values (Fig. 5).

Expression of different forms of organic cation transporter (Oct1, Oct2, Oct3, OctN1, OctN2) mRNA in CP, liver, kidney and ileum.
No detectable expression of Oct2 in CP was found.
With the exception of Wilson's transporter mRNA, which is expressed at a low level in CP, the expression of all metal transporters was high in CP. The expression of Menke's transporter mRNA in CP was more than 15-fold higher than in liver (the organ with lowest expression) and about 2-fold higher than in either kidney or ileum. The expression of DMT1 mRNA in CP was about 1.5-fold greater than in kidney and about 19-fold greater than in liver. Both Menke's transporter and DMT1 mRNA expression was the lowest in liver. The expression of Wilson's transporter mRNA, however, was the highest in ileum and lowest in CP, the expression in CP being about 14% of that in ileum (Fig. 6).

Expression of different metal transporters, DMT1, Menke's, and Wilson's, in CP, liver, kidney, and ileum.
Both DMT1 and Menke's were expressed highly in CP.
Transporters involved in bile acid transport were found to have almost no mRNA expression in CP. Ileal bile acid transporter (Ibat) expression in CP was almost non-existent compared to that in ileum. Sodium taurocholate cotransporting polypeptide (Ntcp) and bile salt excretory protein (Bsep) mRNA expression was about 1.8% and 2.75% of that in liver, respectively, liver being the organ with the highest expression of both Ntcp and Bsep (Fig. 7).

Expression of Ntcp, Bsep, and Ibat mRNA in CP, liver, kidney, and ileum.
None of these bile acid transporters were expressed significantly in CP.
Figure 8 shows the expression of various nucleoside transporters. Constitutive expression of equilibrative nucleoside transporter 1 (Ent1) mRNA in CP was twice as high as that in kidney, the organ with the next highest level of expression, and about 12-fold higher than in ileum, the organ with the lowest level of expression. Expression of Ent2 was high in ileum but was low in CP (about 9% of that in ileum), liver, and kidney. Constitutive expression of both Cnt1 and Cnt2 was highest in ileum. Cnt1 and Cnt2 mRNA expressions in CP was about 85% and 39% of that in ileum, respectively.

Expression of nucleoside transporters in CP, liver, kidney, and ileum.
Both Cnt1 and Cnt2 as well as Ent1 were expressed in CP, whereas Ent2 expression in CP was very low.
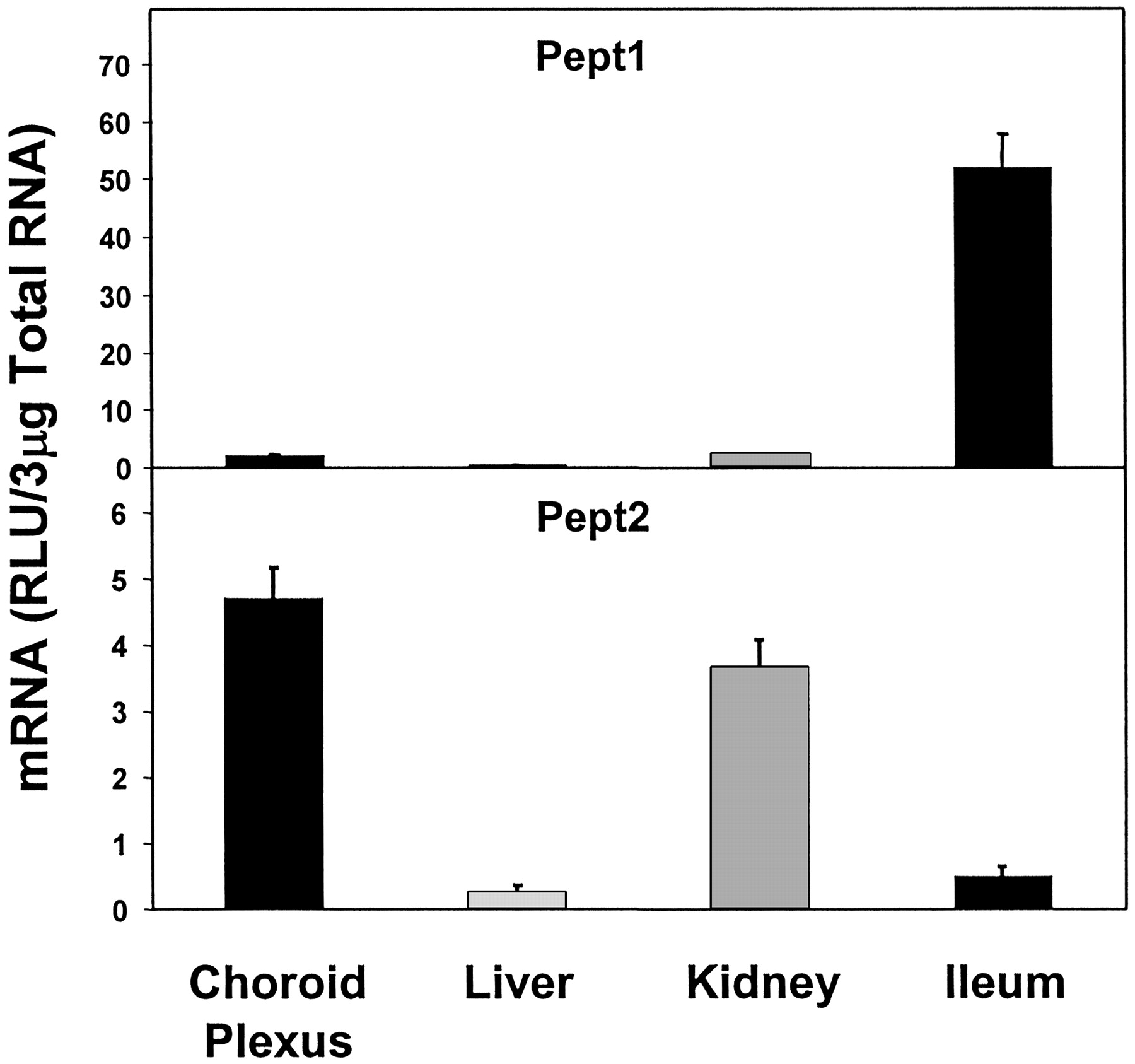
Peptide transporter 2 (Pept2) mRNA expression was highest in CP and was slightly lower in kidney (78% of that of CP). Liver and ileum both had very low-level of Pept2 mRNA, the expression in liver being the lowest, about 5.5% of that in CP (Fig. 9). Peptide transporter 1 (Pept1) expression was very low in CP, liver and kidney, but was high in ileum. The expression of Pept1 in CP was about 4% of that in ileum (Fig. 9).
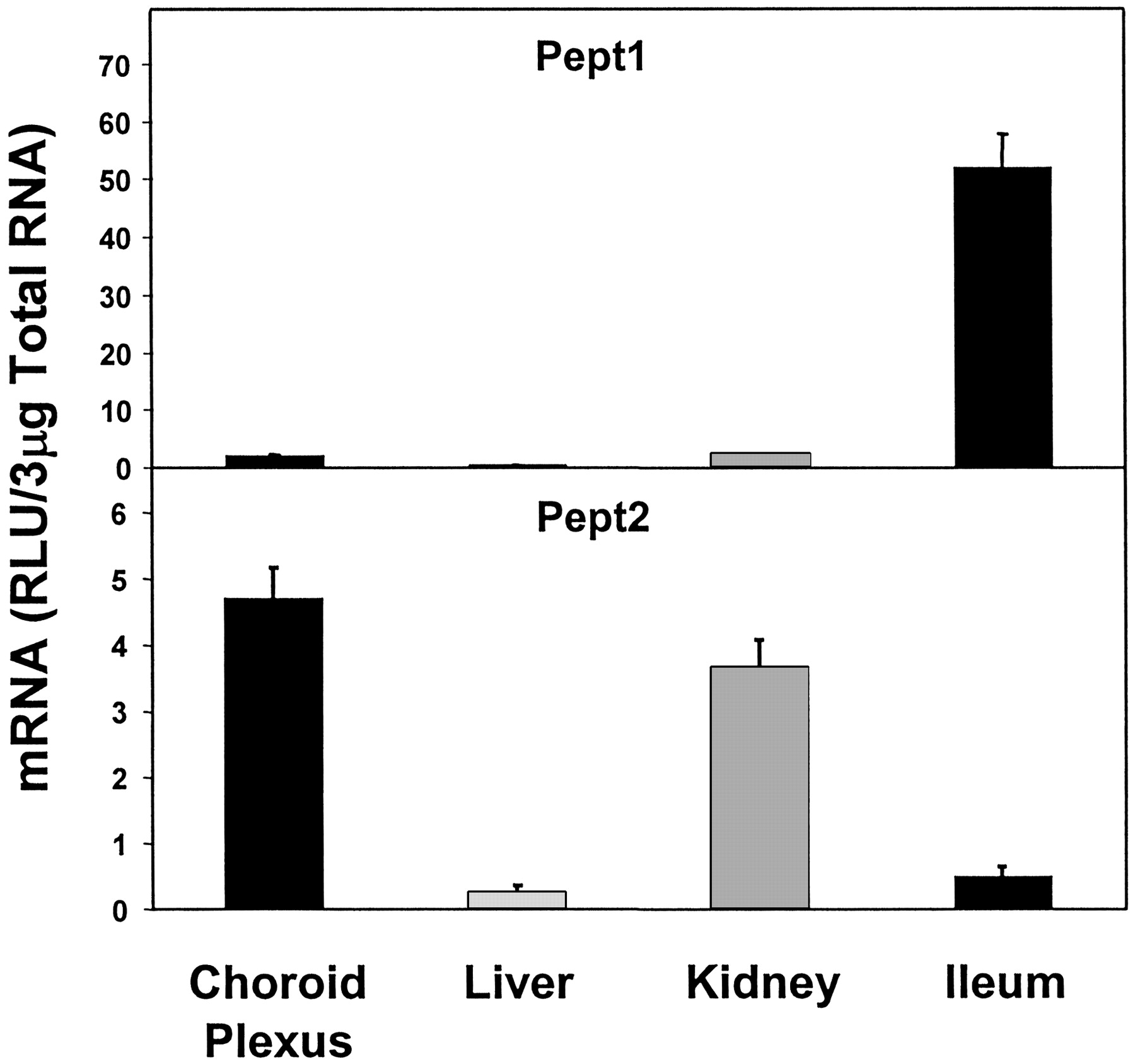
Expression of peptide transporters in CP, liver, kidney, and ileum.
Pept2 expression was very high, but Pept1 expression was very low in CP.
The expression of AbcG5 and AbcG8 mRNAs was very low in CP as compared with ileum, where their expression was the highest, being about 4% and 14%, respectively, of that in ileum (Fig. 10).

Expression of ABC transporters AbcG5 and AbcG8 in CP, liver, kidney, and ileum.
AbcG5 and AbcG8 were both expressed at low levels in CP but at very high levels in ileum.
Discussion
In the present study, the constitutive mRNA expression level of 39 different transporters has been determined in the choroid plexus (CP) of adult Sprague-Dawley rats, and the expression has been compared with that in liver, kidney, and ileum. The overall results show a differential constitutive expression pattern of transporters in the CP. Such differential expression attests to the selective transport activity of the CP, which in turn is responsible, at least in part, for the selective distribution of xenobiotics and endobiotics in the central nervous system (CNS).
The role of multidrug resistance proteins (Mrps) in the transport of xenobiotics from CSF to blood has been reported by Rao et al. (1999) and more recently by Wijnholds et al. (2000). Using a combination of Mrp1/Mdr1a/Mdr1b triple-knockout mice, along with Mdr1a/Mdr1b double-knockout mice, Wijnholds et al. (2000) observed a 10-fold increase in the etoposide levels in CSF in the triple-knockout mice compared with the double-knockout mice, after i.v. administration of the drug. This strongly suggests a role of Mrp1 in etoposide transport from CSF to blood. Studies comparing the role of P-glycoprotein (P-gp) and MRP1 in human CP epithelial transport found that MRP1 played a major role in the transport of xenobiotics from CSF to blood. In contrast, P-gp seemed to prevent movement of certain substances from CSF (Nishino et al., 1999; Rao et al., 1999).
Of the three multidrug resistance genes examined here, that is, Mdr1a, Mdr1b, and Mdr2, none is highly expressed in CP. Brady et al. (2002) showed that the constitutive expression of Mdr1a and Mdr1b was low in both cerebral cortex and cerebellum, but it was slightly higher than in liver. However, Mdr2 was expressed at a higher level in liver than in CP. Although Mdr1a, 1b, and 2 were expressed in CP, the high expression of Mdr1a and 1b in ileum indicates that intestinal P-gp is a major determinant of intestinal elimination of xenobiotics that are P-gp substrates. However, the low-level expression of Mdr1a and Mdr1b mRNAs in CP can still contribute to efflux transport across the CP epithelium (Rao et al., 1999). Gene knockout studies have provided compelling evidence for a role of these transporters in efflux transport across the blood-brain and blood-CSF barrier (reviewed by Schinkel, 1999; Wijnholds et al., 2000). In contrast to Mdr1a and 1b, the very high expression of Mdr2 in liver was expected because of its specific role as a “flippase” in transporting phospholipids.
Among the organic anion-transporting polypeptides (Oatp1 through Oatp5, Oatp9, and Oatp12) as well as organic anion transporter K (Oat-K), Oatp3 mRNA was very highly expressed in CP. Oatp2, albeit most highly expressed in liver, was also expressed in CP. Gao et al. (1999), using in situ hybridization histochemistry, showed that Oatp2 mRNA and protein are expressed on the basolateral membrane (blood side) of CP epithelium. Surprisingly, they also found brush-border membrane (CSF side) localization of Oatp1 in CP epithelium. In the present study, almost no Oatp1 mRNA expression was observed in CP. Our finding is supported by two recent publications that reported high Oatp3 expression along the brush-border membrane of the CP epithelial cells, but almost no Oatp1 expression in the CP (Kusuhara et al., 2003; Ohtsuki et al., 2003). Both these authors utilized RT-PCR amplification to study the expression of Oatp1 and Oatp3 mRNA. Many amphipathic compounds (drugs, drug conjugates, small peptides) could be substrates of rat Oatp2 as well as human OATP-A, such as, the cyclic opioid pentapeptide [d-penicillamine2,5]-enkephalin (Gao et al., 1999, 2000). This further underscores the importance of Oatp2 expression in the CP and its possible role in the transport of compounds from blood to CSF. The opposite polar localization of Oatp2 and Oatp3 in the CP epithelium may serve complementary functions in the transport of amphipathic organic anions into and out of CSF. Walters et al. (2000) showed that Oatp3 mRNA is expressed in rat brain. But the very high-level constitutive expression of Oatp3 mRNA in CP, as observed in the current study, was not exactly reported earlier. The abundant Oatp3 expression reported by Ohtsuki et al. (2003) and Kusuhara et al. (2003) was demonstrated using RT-PCR. Although their results support the finding of the present study, technically, RT-PCR involves amplification of the target. In contrast, the bDNA assay amplifies the signal intensity but not the target itself. Therefore, using the bDNA assay, we have been able to demonstrate the real-time existence (nonamplified) of the high constitutive level of Oatp3 mRNA in CP. It strongly suggests an important role of Oatp3 in the transport of xenobiotics/endobiotics between blood and CSF. In previous studies, such a high level of Oatp3 mRNA expression in CP was probably diluted in the context of expression in the whole brain. In the current study, the expression of Oatp3 mRNA in liver, kidney, and ileum was almost nonexistent compared with that in CP.
Among the organic anion transporters (Oats), Oat2 was found to exhibit a very significant gender difference in its expression in kidney (Buist et al., 2002). Oat3 was cloned from CP by Nagata et al. (2002). They found Oat3 mRNA expression on the CSF side of the CP epithelium. They also found that the Km of uptake of benzylpenicillin (PCG) by LLC-PK1 cells transfected with rat Oat3 was similar to that by intact CP, which strongly suggests that PCG is transported by Oat3 in adult CP. In addition to PCG, cimetidine and probenecid also showed Ki values for Oat3 almost an order of magnitude lower than that of Oat1, further suggesting an important primary role for Oat3 in their transport. Recently, Gibbs and Thomas (2002) reported that the limited distribution of the human immunodeficiency virus (HIV) reverse transcriptase inhibitor 2′3′-dideoxycytidine in CSF of guinea pigs was due to a probenecid-sensitive, Oat1/Oat3-like efflux transporter activity. Further evidence underscoring the importance of Oat3 in organic anion uptake in CP comes from Oat3 knockout mice (Sweet et al., 2002). These mice appear healthy, fertile, and without any gross morphological tissue abnormalities. However, the kidneys and the CP of these Oat3-/- mice show a substantial decrease in their ability to transport organic anions. In contrast, no discernible differences were observed in the uptake ability of hepatic slices from wild-type and Oat3-/- mice, suggesting that Oat3 does not play a major role in hepatic organic anion uptake, but it probably plays a major role in CP. In the present study, Oat2 and Oat3 were found to be expressed in CP and Oat3 mRNA was highly expressed, whereas Oat1 expression was insignificant. Oat1 mRNA expression was also found to be absent in adult mouse CP (Pavlova et al., 2000).
The expression of Octs, in general, was higher in kidney and ileum than in CP, but the expression of OctN1 and OctN2 in CP was much higher than in liver. Both rat OctN2 and human OCTN2 transport carnitine and tetraethylammonium. Human OCTN1 and OCTN2 can transport antiarrhythmic drugs and β-lactam antibiotics, respectively. Sekine et al. (1998) and Slitt et al. (2002) have reported a very low-level expression of OctN2 in rat brain. It appears that the relatively high expression of OctN2 in CP is a very localized expression that is not detected when the expression of OctN2 mRNA is measured in the context of the whole brain. In contrast, OctN1 expression is detectable in cerebellum and cerebral cortex (Slitt et al., 2002). Thus, the role of OctN2 in transport across CP epithelium should be investigated in more detail.
Sweet et al. (2001) showed that rat Oct1 and Oct2 as well as human OCT2 expressed in Xenopus oocyte could transport choline, but Oct3 did not. The estimated Km of rat Oct1-mediated choline transport was 346 ± 50 μM and that of Oct2-mediated choline transport was 441 ± 67 μM. They also showed that in freshly isolated intact CP, choline was transported in a concentration-dependent manner with an estimated Km of 183 μM. The presence of Oct1, 2, and 3 mRNAs in CP was also demonstrated by RT-PCR analysis. In the present study, a low-level constitutive expression of Oct1 as well as Oct3 mRNA was detectable in CP, whereas that of Oct2 mRNA was almost nondetectable. This apparent discrepancy is most likely due to different techniques employed in the two studies. In the present study, constitutive expression was measured using a bDNA signal amplification technique, but in the study by Sweet et al. (2001), RT-PCR was used to detect expression. Even though the bDNA signal amplification technique is much more sensitive compared with other classical mRNA detection techniques, it is still less sensitive than PCR, which in theory can amplify the signal from a single molecule. Additionally, to show the subcellular localization of Oct2, Sweet et al. (2001) transfected CP with green fluorescent protein-tagged Oct2 and demonstrated its apical localization. This method, again, is supposed to be more sensitive than in situ hybridization or immunocytochemistry, which could have been used to detect constitutive expression and localization (apical versus basal) of Oct2 in CP. Thus, the study by Sweet et al. (2001) demonstrated that rat Oct2 could transport choline as a substrate, and that Oct2 mRNA expressed in CP would be localized to the CSF side of the CP epithelial cells. However, the Km of choline transport (183 μM) observed with intact CP was lower than that observed with the Xenopus oocyte system (441 ± 67 μM), suggesting that additional choline transport system(s) may exist in intact CP.
The role of divalent metal transporter1 (DMT1), also known as divalent cation transporter 1 (DCT1), in iron transport has been studied in detail, but it is also known to transport divalent metals including zinc and cadmium (Gunshin et al., 1997; Tallkvist et al., 2001; Park et al., 2002). Previous studies on the localization of DMT1/DCT1 revealed the existence of DMT1/DCT1 mRNA in CP epithelium, but not in ependymal cells (Qian and Wang, 1998). At present, the function of DMT1 is not fully understood, but its cellular localization does strongly suggest its important transport function in the nervous system. The expression of cytosolic metal binding proteins, such as metallothionein I and II, had also been reported to be very high in CP (Choudhuri et al., 1995). It appears that DMT1, in conjunction with cytosolic metal binding proteins, may play an important role in the homeostasis of divalent metals in the CNS. Such potential functional coordination between metal transporters and metal-binding proteins is conceivable and may be important in maintaining metal homeostasis in the CNS, because even “physiological metals” like zinc can be neurotoxic in the free state (Choudhuri et al., 1995). Like DMT1, the expression of Menke's transporter was also highest in CP compared with that in liver, kidney, and ileum, whereas the expression of Wilson's was low but detectable in CP and highest in ileum. Two inherited human diseases, X-linked Menke's disease and autosomal-recessive Wilson's disease, highlight the importance of intact cellular copper transport mechanism (Kuo et al., 2001). In humans, for example, mutation in ATP7A, a Cu-ATPase, causes the copper deficiency condition Menke's disease, which is fatal in early childhood. In contrast, mutation in ATP7B, another Cu-ATPase, results in genetic toxicosis, Wilson's disease. Therefore, it has been proposed that the physiological regulation of copper is accomplished mainly by the intracellular copper-regulated trafficking of the Cu-ATPases (Mercer and Llanos, 2003). Although it is not clear yet what role these transporters play in CP, it is likely that they might control the copper traffic between blood and CSF.
The significance of differential expression of equilibrative nucleoside transporters Ent1 and Ent2 in CP is not clear. The very high-level expression of Ent1, but a very low-level expression of Ent2 mRNA in CP, combined with their opposite expression pattern in ileum, suggests that distribution of nucleosides and nucleoside-analogs from the intestine to blood may be mostly mediated by Ent2, whereas distribution from blood to CSF may be mostly mediated by Ent1. Although the subcellular localization of Ent1 and Ent2 in CP has not been demonstrated yet, it is tempting to speculate that Ent1 and Ent2 may have opposite regional expression in CP epithelium; that is, one being expressed on the blood-side, whereas the other is expressed on the CSF side. If opposite polar localization of Ent1 and Ent2 in the CP epithelium exists, it may aid complementary functions of these two transporters in the transport of nucleosides and nucleoside-analog drugs into and out of the CSF. Although the expression of Ent1 was found to be much higher than Ent2 in CP in the present study, the overall expression levels of Ent2 mRNA in different rat brain areas may be much higher than that of Ent1 (Anderson et al., 1999).
Between the peptide transporters Pept1 and Pept2, the latter was found to be highly expressed in CP. Recently, Teuscher et al. (2001) demonstrated Pept2-mediated uptake of a number of small neuropeptides in CP and their subsequent transport to CSF. Thus, Pept2 may play an important role in regulating neuropeptides, peptide fragment, and peptidomimetics in CSF. Shu et al. (2002) studied the directionality of Pept2-mediated transport of glycylsarcosine (GlySar) in cultured CP epithelium and found that the Km of uptake of GlySar from the apical side was 59.6 μM, as opposed to the Km of uptake from the basal side, which was 1.4 mM. The uptake from the apical side was consistent with high-affinity Pept2-mediated GlySar uptake, indicating that Pept2 might be an important efflux transporter carrying peptides and/or mimetics from CSF to blood.
Transporters known to be important in bile acid transport, such as Ntcp, Bsep, or Ibat, and those responsible for sterol uptake, such as AbcG5 and AbcG8, showed very low-level expression in CP. The expression of Ntcp and Bsep was highest in liver, and that of Ibat, AbcG5, and AbcG8 was highest in ileum. The low expression level of transporters involved in sterol uptake in the CP suggests that these transporters probably do not play an important role in the CNS level of cholesterol. Recent evidence indicates that a high cholesterol level in the CNS may be an important risk factor for Alzheimer's disease, because hypercholesterolemia facilitates deposition of amyloid Aβ and formation of amyloid plaques in the CNS (Buxbaum et al., 2001).
In conclusion, in the present study the constitutive expression of 39 different transporters was investigated in CP, and the expression was compared with that in liver, kidney, and ileum. The present study, which was done by bDNA assay through use of oligoprobes, will serve as a useful reference of constitutive transporter expression in CP of adult rats. Among the transporters studied, Mrp1, Mrp4, Mrp5, Oatp3, Menke's, DMT1, Ent1, and Pept2 were found to have the highest level of constitutive mRNA expression in CP; that is, higher expression than in liver, kidney, and ileum. Mrp6, Oct3, OctN1, OctN2, Oatp2, Oatp9, Wilson's, Oat2, Oat3, Cnt1, and Cnt2 had high levels of mRNA expression in CP, but the expression was lower than that in liver, kidney, and ileum. In contrast, Mrp2, Mrp3, Oct1, Oct2, Oatp1, Oatp4, Oatp5, Oatp12, Oat-K (1/2), Ntcp, Bsep, Ibat, Mdr1a, Mdr1b, Mdr2, Oat1, Ent2, Pept1, AbcG5, and AbcG8 had the lowest or almost no constitutive mRNA expression in CP. Thus, the coordinated expression of a number of transporters in CP underscores its role in maintaining the delicate balance in homeostasis of different endo- and xenobiotics in the CNS.
Acknowledgments
We thank Xenotech, LLC (Kansas City, KS) for supplying all rat brains, and Dr. Nichole Vansell, Dr. Dylan Hartley, and Dr. Yaping Liu for assisting in the collection of all brains.
Footnotes
-
↵1 Present Address: U.S. Food and Drug Administration, CFSAN/OFAS/DBGNR, 5100 Paint Branch Parkway, HFS-255, College Park, MD 20740.
-
↵2 These authors contributed equally to this work
-
↵3 Abbreviations used are: ABC, ATP-binding cassette; bDNA, branched DNA; Mrp, multidrug resistance protein; Mdr, multiple drug resistance; Oatp, organic anion transporting polypeptide; Oat-K, organic anion transporter K; Oct, organic cation transporter; Ntcp, sodium taurocholate cotransporting polypeptide; DMT1, divalent metal transporter 1; Ent, equilibrative nucleoside transporter; Cnt, concentrative nucleoside transporter; Pept, peptide transporter; Ibat, ileal bile acid transporter; RLU, relative luminescence unit(s); CP, choroid plexus; CNS, central nervous system; P-gp, P-glycoprotein; CSF, cerebrospinal fluid; RT-PCR, reverse transcription-polymerase chain reaction; PCG, benzylpenicillin; DCT1, divalent cation transporter1.
-
This work was supported by National Institutes of Health Grant ES-09716. This work was conceived of and performed entirely at the University of Kansas Medical Center. Opinions expressed in this article are authors' personal opinion and they do not reflect U.S. Food and Drug Administration's opinion.
- Received May 12, 2003.
- Accepted August 12, 2003.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}