


Abstract
The bovine uterine is easily contaminated with bacteria during coitus or parturition. A previous study suggested that prostaglandin E2 (PGE2) promoted Escherichia coli–infected bovine endometrial tissue inflammatory damage via cyclooxygenase-2 (COX-2) and microsomal prostaglandin E synthase-1 (mPGES-1). However, it remains unclear which PGE2 receptors regulate the proinflammatory effect of PGE2. In this study, we evaluated the effect of PGE2 and its mediated receptors on E. coli–infected endometrium explants isolated from the bovine uterus. The E. coli–infected bovine endometrial explants were cultured in vitro, and the study used EP2/4 receptor agonists to investigate the responses of COX-2, mPGES-1, PGE2, proinflammatory factors, and damage-associated molecular patterns (DAMPs). The expression of COX-2, mPGES-1, PGE2, proinflammatory factors, and DAMPs was significantly increased after infection with E. coli; however, the high expression levels caused by E. coli were reduced following treatment with COX-2 and mPGES-1 inhibitors. In addition, the expression levels of COX-2, mPGES-1, PGE2, proinflammatory factors, and DAMPs were higher in treatment with EP2/4 receptor agonists in E. coli–infected endometrium explants, and their promotable effects were effectively blocked by EP2/4 receptor antagonists. These findings provide evidence that PGE2 may promote the progress of inflammation in endometrial explants infected with E. coli in bovines. Furthermore, EP2/4 may be involved in a positive feedback loop for COX-2 and mPGES-1 expression, and this may be responsible for the proinflammatory reaction of PGE2 in E. coli–infected uteri of bovines.
SIGNIFICANCE STATEMENT PGE2 promoted E. coli–infected bovine endometrial tissue damage via COX-2 and mPGES-1. However, this proinflammatory effect of PGE2 depends on which receptors are affected by PGE2, and this remains unclear. In this study, it was investigated that EP2 and EP4 may be involved in a positive feedback loop for COX-2 and mPGES-1 expression, and this may be responsible for the proinflammatory reaction of PGE2 in E. coli–infected uteri of bovines.
Introduction
During sexual intercourse or parturition in bovine, the uterus is easily contamination with bacteria (Herath et al., 2006, 2009a). Endometrial epithelial and stromal cells sense these pathogen-associated bacteria and release cytokines and chemokines, including interleukins (ILs) IL-1β, IL-6, and IL-8, and damage-associated molecular patterns (DAMPs) including high mobility group box-1 (HMGB-1) and hyaluronan, which causes endometrial inflammation and tissue damage, even infertility (Herath et al., 2009b; Sheldon, 2014; Sheldon et al., 2014). A recent study demonstrated that bovine endometrial explants infected with Escherichia coli accumulate prostaglandin E2 (PGE2), resulting in the expression of IL-1β, IL-6, IL-8, tumor necrosis factor-α (TNF-α), HMGB-1, and hyaluronan (Li et al., 2019a,c), indicating PGE2 may promote the progress of E. coli–infected endometrial inflammation via increasing the expression of proinflammatory factors and DAMPs. However, the receptors responsible for this effect of PGE2 remain unclear.
Among the five principal bioactive prostaglandins generated in vivo, PGE2 is involved in all of the processes leading to the classic signs of inflammation. Research has shown that PGE2 accumulated when E. coli was used to stimulate endometrial cells and explants in vitro (Herath et al., 2006), which was influenced by the degree of endometritis and concentrations of uterine endotoxins (Mateus et al., 2003). Among the synthases of PGE2, cyclooxygenase-2 (COX-2) and microsomal prostaglandin E synthase-1 (mPGES-1) are the more important sources of prostanoid formation in inflammation and proliferative diseases (Ricciotti and FitzGerald, 2011). Previous research has demonstrated a dramatic increase in COX-2 and mPGES-1 in response to various proinflammatory stimuli to generate a transient spike in PGE2 levels (Crofford et al., 1994, 1997; Roshak et al., 1996; Alaaeddine et al., 1999; Murakami et al., 2000), and the deletion of these two synthases suppressed synovial inflammation and joint destruction in a collagen-induced arthritis model (Myers et al., 2000; Ochi et al., 2003; Trebino et al., 2003), which indicated PGE2 plays an essential role in the inflammatory process via COX-2 and mPGES-1.
The complex biologic effects of PGE2 are mediated by four distinct G protein–coupled receptors, namely, the EP1, EP2, EP3, and EP4 receptors (Hata and Breyer, 2004). Among the four receptors, the EP2 and EP4 receptors distribute mainly in the uterus (Sugimoto et al., 1992; Honda et al., 1993; Watabe et al., 1993; Katsuyama et al., 1995) and bind to G proteins, and then they activate adenylate cyclase to produce cyclic 3,5-adenosine monophosphate (cAMP), finally resulting in a series of inflammatory events (Regan, 2003; Cheon et al., 2006; Yokoyama et al., 2013). However, EP2 and EP4 receptors also play distinct roles in other processes. For example, in human macrophages infected with Mycobacterium tuberculosis, the EP4 receptor is integral to a positive feedback loop for PGE2 production, while the EP2 receptor is only responsible for protection against necrosis (Nishimura et al., 2013). These differences between EP2 and EP4 receptors may be related to their respective signaling pathways. Both in EP2 and EP4 receptor signaling, increased cAMP levels cause the activation of protein kinase A (PKA) followed by a series of proinflammatory actions. However, the EP4 receptor is also coupled to phosphatidylinositol 3-kinase, which may be achieved by inhibiting Gi rather than Gs (Fujino et al., 2003; Cheon et al., 2006; Yokoyama et al., 2013), and is also involved in inflammation. Interestingly, the EP2/PKA and EP4/phosphatidylinositol 3-kinase pathways can act independently in the same cells, indicating that PGE2 evokes different functions by stimulating either the EP2 or EP4 receptor (Yokoyama et al., 2013). Given these aspects, either EP2 or EP4 or both receptors may be involved in the pathologic accumulation and proinflammatory action of PGE2 in E. coli–infected endometrial explants.
The present study applied an EP2 receptor-selective agonist (butaprost) and an EP4 receptor-selective agonist (CAY10598) to evaluate the expression or release of COX-2, mPGES-1, PGE2, IL-1β, IL-6, IL-8, TNF-α, HMGB-1, and hyaluronan in E. coli–infected bovine endometrial explants in vitro. The results will be helpful in elucidating the regulatory role of pathologically accumulated PGE2 and its receptors on inflammatory mediator expression and tissue damage, and further provide guidance for clinical medication, especially nonsteroidal anti-inflammatory drugs (NSAIDs) or prostaglandin preparations that can be used to treat bacterial endometritis of bovines.
Materials and Methods
Reagents, Chemicals, and Antibodies.
The following reagents, chemicals, and antibodies were used: FBS (ExCell Biology, Inc., China); Dulbecco’s modified Eagle’s medium/F-12 (Gibco); penicillin and streptomycin (Gibco); amphotericin B (GENERAY, China); six-well culture plates (Corning); T-PER Tissue Protein Extraction Regent (Thermo Fisher Scientific); Halt Protease Inhibitor (Thermo Fisher Scientific); SDS-PAGE Loading Buffer (TAKARA, Japan); Pierce BCA Protein Assay Kit (Thermo Fisher Scientific); Prestained Protein Ladder (Thermo Fisher Scientific); centrifugal filter units (Millipore); SDS-PAGE Kit (Solarbio, China); Trizma Base (Sigma); SDS (GENERAY); 10X Tris/glycine buffer (Bio-Rad); transfer membranes (Millipore); Starting Block T20 (Tris-buffered saline) Blocking Buffer (Thermo Fisher Scientific); antibody dilution (Beyotime, China); pPrimary antibody: COX-2 antibody (Abcam, Cambridge, UK), mPGES-1 antibody (CAYMAN), rabbit anti-HMGB-1 antibody (Novus Bio, CO), hyaluronan binding protein 1 (HABP1) antibody (Abbexa, UK); secondary antibody: goat anti-rabbit IgG horseradish peroxidase–linked and goat anti-mouse IgG horseradish peroxidase–linked (Cell Signaling Technology); goat-donkey anti-rabbit IgG H&L antibody (Alexa Fluor 647) (Abcam); AxyPrep Multisource Total mRNA Miniprep Kit (Axygen Scientific); Primer Script RT Master Mix (TAKARA); SYBR Green Master (Rox) (Roche, Germany); Luria-Bertani broth (OXOID, UK), and optimal cutting temperature compound (SAKURA). The primers were synthesized by Invitrogen (China). Table 1 lists the agonists and inhibitors used in the study.
Agonists and inhibitors used in this study
Collection and Cultivation of Endometrial Explants In Vitro.
All of the animal studies were conducted in accordance with the experimental practices and standards approved by the Animal Welfare and Research Ethics Committee of Inner Mongolia Agricultural University, and all efforts were made to minimize animal suffering. Fresh and healthy bilateral uterine horns near the ovaries were obtained from 40 healthy spawning Holstein cows (age: 24 months; weight: ∼600 kg) in a local slaughterhouse according to the protocols of Ireland et al. (1980) and were kept on ice until further analysis. First, the uterine horns were trimmed and opened longitudinally under aseptic conditions followed by washing three times with sterile PBS supplemented with 100 IU/ml penicillin and streptomycin and 2.5 μg/ml amphotericin B. The horns were then stored at 4°C for 1 hour. Next, endometrial tissues containing epithelium and stroma was removed from the uterine horns using curved scissors and ophthalmic tweezers, and the explants was trimmed into pieces approximately 2 mm long, 2 mm wide, and 1 mm thick. Finally, these pieces were randomly placed in six-well plates with culture medium (Dulbecco’s modified Eagle’s medium/F-12 with 20% FBS, 100 IU/ml penicillin and streptomycin, and 2.5 μg/ml amphotericin B at 5 ml/well) and incubated in a humidified environment (95% air with 5% CO2) at 37°C for 2 days (Zhang et al., 2017; Li et al., 2019a). The medium was replaced at 24-hour intervals until subsequent treatment.
Preparation of Escherichia coli Suspension.
To ensure the clinical pathogenicity of the E. coli strain used in the experiment, we selected bovines with clinical endometritis (identification certificate number: SYS110017) characterized chiefly by tissue damage, necrosis, and the accumulation of pus in the genital tract. The isolated endometrial pathogenic E. coli from the uteruses was cultured overnight in Luria-Bertani broth medium containing 5% FBS at 37°C and 200 rpm until the culture reached the log phase. The broth was then centrifuged at 6000g for 10 minutes at 4°C, and the pellet was washed in sterile PBS and resuspended in tissue culture medium for later testing (Li et al., 2019a).
Experimental Treatments.
After being cultured for 1 day, the endometrial explants, except those in the control groups, were treated with 1 × 106 colony forming units/ml of live E. coli following the replacement of the medium. The explants were divided into groups as follows: a control group, an endometrial pathogenic E. coli group, a EP2 receptor agonist treatment group (butaprost; 10−7 M), a EP4 receptor agonist treatment group (CAY10598; 10−9 M), COX-2 inhibitor treatment groups (CAY10404 and NS398; 10−6 M), mPGES-1 inhibitor treatment groups (MF63 and MK886; 10−6 M), EP2 receptor agonist plus COX-2 inhibitor treatment groups, EP2 receptor agonist plus mPGES-1 inhibitor treatment groups, EP4 receptor agonist plus COX-2 inhibitor treatment groups, and EP4 receptor agonist plus mPGES-1 inhibitor treatment groups.
A verification experiment was administrated as follows: a control group, an endometrial pathogenic E. coli group, EP2 receptor agonist treatment group, EP2 receptor antagonist (AH6809; 10−5 M) plus EP2 receptor agonist treatment group, and EP4 receptor antagonist (AH23848; 10−7 M) plus EP2 receptor agonist treatment group. For the experiment with the EP4 receptor, the EP4 receptor agonist was applied substituting for the EP2 receptor agonist. All of the groups except for the control group were treated with live E. coli before the reagent treatments, and the groups were incubated for 12 hours.
The collected explants were washed with PBS, snap-frozen in liquid nitrogen, stored at −80°C until mRNA and protein extraction, and soaked in optimal cutting temperature compound before −80°C storage for immunofluorescence analysis. In addition, a portion of explants were fixed in 4% paraformaldehyde, and paraffin sections were made for hematoxylin-eosin staining.
Real-Time Reverse-Transcription Polymerase Chain Reaction Analysis.
Total mRNA was isolated from endometrial explants using the AxyPrepTM Multisource Total RNA Miniprep Kit (Axygen Scientific Inc.). Real-time and reverse-transcription polymerase chain reaction (RT-PCR) analyses were conducted according to the manufacturer’s instructions (PrimeScript RT Master Mix from Takara Bio Inc. and Fast Start Universal SYBR Green Master from Roche Applied Science). cDNA was amplified as follows: denaturing at 50°C for 2 minutes and then at 95°C for 10 minutes; 40 cycles at 95°C for 15 seconds and at 60°C for 30 seconds (ABI Quantstudio 7). Table 2 lists the primers used for the quantitative polymerase chain reaction. The annealing temperature was 58°C, and the results are presented as 2−ΔΔct.
Primer sequences for quantitative polymerase chain reaction
ELISA Analysis.
Concentrations of PGE2, IL-1β, IL-6, IL-8, and TNF-α in explant culture supernatants were measured by ELISA according to the manufacturer’s instructions (Prostaglandin E2 ELISA Kit-Monoclonal 514010 from Cayman; Bovine IL-1β ELISA Reagent Kit ESS0027 from Kingfisher Biotech; Bovine IL-6 ELISA Reagent Kit DY8190 from R&D Systems, Bovine IL-8 ELISA Kit CSB-E130528 from CUSABIO; and Bovine TNF-α Duo Set DY2279 from R&D Systems Europe Ltd.).
Western Blot Analysis.
Total protein extraction, concentration measurements, and protein denaturation were conducted according to the manufacturers’ instructions (T-PERTM Tissue Protein Extraction Reagent; Thermo Fisher Scientific for protein extraction; PierceTM BCA protein Assay Kit; Thermo Fisher Scientific for protein concentration measurement; SDS-PAGE sample loading buffer; and Beyotime for protein denaturation). Denatured proteins samples (20 µg) were subjected to 12% SDS-PAGE at 100 V. After semidry blotting, the proteins were transferred onto a polyvinylidene fluoride membrane set at 40 V for 40 minutes, and then the membranes were subsequently blocked with 3% bovine serum albumin in Tris-buffered saline (0.1% Tween 20) for 3 hours at room temperature. The blocked membranes were incubated with the primary antibodies for 10 hours at 4°C followed by three washes with Tris-buffered saline and Tween. The membranes were then incubated with the secondary antibody (1:7500) for 40 minutes at room temperature prior to three additional washes, and then were finally exposed to enhanced chemiluminescence western blot detection reagents and enhanced chemiluminescence film. The primary antibody dilutions were as follows: COX-2, 1:1000; mPGES-1, 1:200; HMGB-1, 1:1000; and HABP1 1:200. The band density was quantified using the ImageJ software program. The density of the target protein band was normalized based on the density of β-actin in the same samples.
Double-Label Immunofluorescence Assays.
The frozen sections of treated endometrial explants (6 µm) were thawed at room temperature for 15 minutes and then fixed with cold acetone for 10 minutes. These sections were blocked for 1 hour in 3% bovine serum albumin at room temperature, followed by three washes in cold PBS with 0.25% Tween. The sections were subsequently incubated with primary antibodies (HMGB-1, 1:200; and HABP1, 1:20) overnight at 4°C in darkness. The slides were washed three times for 15 minutes using PBS with 0.25% Tween 20 prior to being incubated with donkey anti-rabbit IgG H&L secondary antibody (Alexa Fluor 647) at 1:1000 dilution for 1 hour at room temperature.
Statistical Analysis.
The results are reported as the arithmetic mean ± S.D. Statistical significance was analyzed using Student’s t test or one-way ANOVA, followed by post-test analysis (Dunnett’s test) when applicable. Statistical significance was ascribed for P ≤ 0.05. The same letters represent no significant difference (P > 0.05) and different letters indicate significantly different mean values (P ≤ 0.05), The ImageJ and GraphPad Prism 5 software programs were used in the analyses.
Results
EP2/4 Receptor Agonists Increased the Expression of COX-2 and mPGES-1 Promoting PGE2 Synthesis in E. coli–Infected Bovine Endometrial Explants.
To investigate the effects of EP2/4 receptor agonists on PGE2 production in E. coli–infected endometrial explants, PGE2 synthetase COX-2 and mPGES-1 expression were measured by RT-PCR and western blotting, and PGE2 secretion was examined by ELISA. As shown in Fig. 1, addition of pathogenic E. coli obviously induced COX-2 (Fig. 1, A and B) and mPGES-1 expression (Fig. 1, C and D) and PGE2 secretion (Fig. 1E) compared with that of control group (P < 0.05). Moreover, the expression of COX-2 and mPGES-1 and PGE2 secretion were further enhanced by EP2 receptor agonist (butaprost) or EP4 receptor agonist (CAY10598) treatment compared with the E. coli groups (P < 0.05). By contrast, when COX-2 inhibitors (CAY10404 and NS398) or mPGES-1 inhibitors (MF63 and MK886) were added, the expression and secretion levels of COX-2, mPGES-1 and PGE2 were significantly reduced compared with E. coli treatment alone (P < 0.05). In addition, for EP2/4 receptor agonists with COX-2 or mPGES-1 inhibitors treated together, EP2/4 receptor agonists increased the expression of COX-2 and mPGES-1 and further promoted PGE2 synthesis which reduced by COX-2 or mPGES-1 inhibitors (P < 0.05), indicating EP2/4 receptors were responsible for the pathologic accumulation loop of PGE2 via COX-2 and mPGES-1 in E. coli–infected endometrial explants, and the endogenous PGE2 promotes its own accumulation under pathologic stimulation in the bovine endometrium.
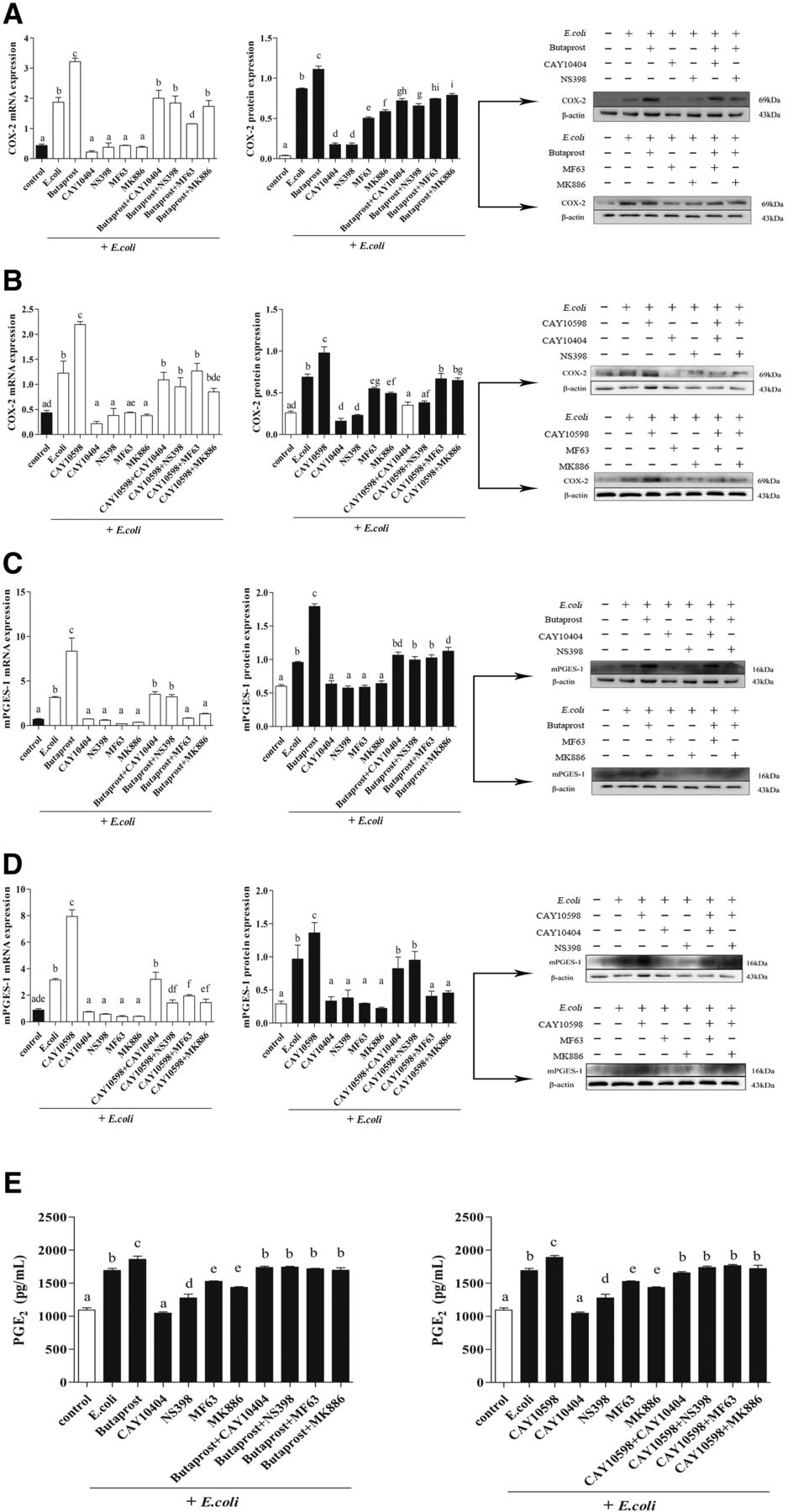
The effect of EP2/4 receptor agonists on COX-2 and mPGES-1 expression and PGE2 secretion in E. coli–infected endometrial epithelium. At 12 hours after treatment with EP2/4 receptor agonists (butaprost, 10−7 M; CAY10598, 10−9 M), COX-2 inhibitors (CAY10404 and NS398; 10−6 M), mPGES-1 inhibitors (MF63 and MK886; 10−6 M), and EP2/4 receptor agonists plus COX-2 and mPGES-1 inhibitors, the relative gene expression of COX-2 (A and B), and mPGES-1 (C and D) was analyzed by real-time polymerase chain reaction using β-actin as a reference gene, protein expression was measured by western blotting, and PGE2 secretion (E) was measured by ELISA [(E and F); n = 3; means ± S.D.]. The same letters represent no significant difference; different letters represent significant differences, which were analyzed by GraphPad Prism 5 software (P ≤ 0.05).
EP2/4 Receptor Agonists Upregulated the Expression of IL-1β, IL-6, IL-8, and TNF-α Reduced by COX-2 and mPGES-1 Inhibitors in E. coli–Infected Bovine Endometrial Explants.
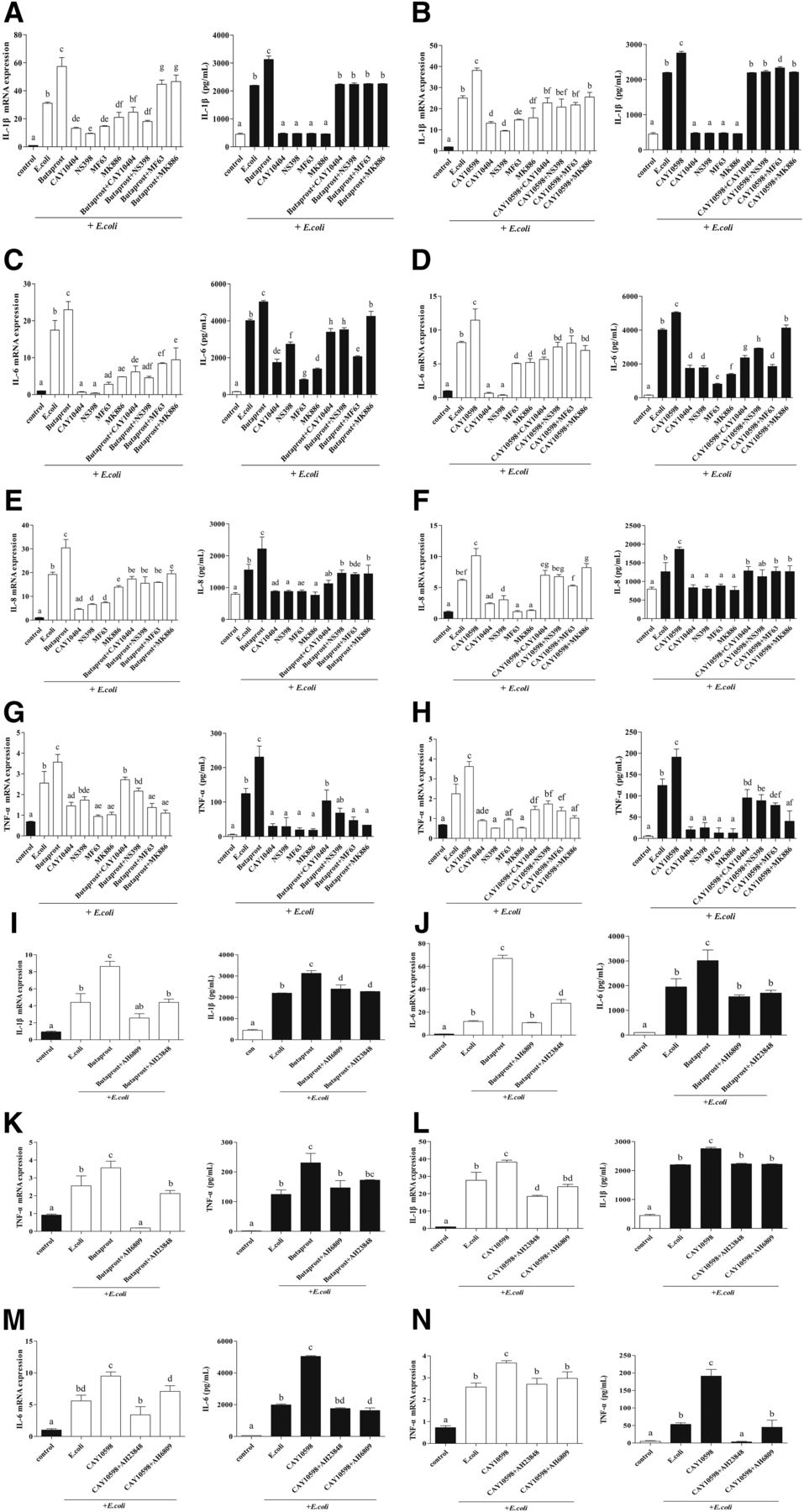
To investigate whether EP2 and EP4 mediate the proinflammatory actions of PGE2 in E. coli–infected endometrial explants, expression of IL-1β, IL-6, IL-8, and TNF-α were evaluated by RT-PCR and ELISA in E. coli–infected bovine endometrium. The production of these four proinflammatory factors was greatly increased by E. coli treatment, and further upregulated by EP2/4 receptor agonists. However, in the COX-2 and mPGES-1 inhibitor treatment groups, these four factor expressions were significantly reduced compared with the E. coli treatment group, which was upregulated by EP2/4 receptor agonists (P < 0.05) (Fig. 2, A–H).

Regulation of IL-1β, IL-6, IL-8, and TNF-α expression in E. coli–infected endometrial epithelium by EP2/4 receptors. The expression of IL-1β (A and B), IL-6 (C and D), IL-8 (E and F), and TNF-α (G and H) in endometrial explants treated with EP2/4 receptor agonists, COX-2 and mPGES-1 inhibitors, and EP2/4 receptor agonists plus COX-2 and mPGES-1 inhibitors for 12 hours was evaluated by RT-PCR and ELISA (n = 3; means ± S.D.). The same letters represent no significant difference; different letters represent significant differences analyzed by GraphPad Prism 5 software (P ≤ 0.05). The expression of IL-1β (I and J), IL-6 (K and L), and TNF-α (M and N) in explants treated with EP2/4 receptor agonists, and EP2/4 receptor agonists plus selective EP2/4 receptor antagonist (AH6809, 10−5 M; AH23848, 10−7 M; n = 3; means ± S.D.) is shown. The same letters represent no significant difference; different letters represent significant differences analyzed by GraphPad Prism 5 software (P ≤ 0.05).
Additionally, a selective EP2 receptor antagonist (AH6809), followed by EP2 receptor agonist in E. coli–infected explants, reduced the expression levels of IL-1β, IL-6, and TNF-α significantly compared with the EP2 receptor agonist alone. More interestingly, this phenomenon also occurred in the group with a selective EP4 receptor antagonist (AH23848) plus EP2 receptor agonist (P < 0.05) (Fig. 2, I–K), indicating that EP2 and EP4 work together to mediate the PGE2-induced inflammation in E. coli–infected endometrial epithelium explants. This was further confirmed by the results from EP4 receptor agonist plus EP4/2 receptor antagonists (P < 0.05) (Fig. 2, L–N). These results suggest that the pathologic accumulation of PGE2 through activating the EP2/4 receptors promotes the expression of proinflammatory factors, such as IL-1β, IL-6, IL-8, and TNF-α, when the bovine endometrium is infected with pathogenic E. coli.
EP2/4 Receptors Were Involved in the Expression of Damage-Associated Molecular Patterns and Tissue Damage in E. coli–Infected Bovine Endometrial Explants.
During inflammation caused by E. coli, endometrial explants not only secrete inflammatory factors but also cause the production of damage-associated molecular patterns, such as HMGB-1 and HABP1, thereby enhancing the tissue damage process (Scaffidi et al., 2002; Johnson et al., 2015; Komai et al., 2017). Accordingly, the present study evaluated HMGB-1 and HABP1 by RT-PCR and western blotting in E. coli–infected bovine endometrial explants. After E. coli treatment, the EP2/4 receptor agonists significantly increased the mRNA expression of HMGB-1 (Fig. 3, A and B) and HABP1 (Fig. 3, D and E) as well as protein production, but reduced the mRNA expression in the COX-2 and mPGES-1 inhibitor treatment groups. In contrast, when treated together with COX-2 and mPGES-1 inhibitors, the two receptor agonists induced promotion effects on HMGB-1 and HABP1 expression that were lower than those observed with agonists alone (P < 0.05). To more intuitively clarify the expression pattern of the two factors and the effects of EP2/4 receptor agonists in E. coli–infected endometrial explants, this study applied immunofluorescence staining for HMGB-1 and HABP1. The strongest fluorescence was found in the EP2/4 receptor agonists groups, and the COX-2 or mPGES-1 inhibitors obviously decreased the fluorescence signaling in the E. coli–infected explants, all of which was consistent with the expression levels of mRNA and protein in HMGB-1 and HABP1 (Fig. 3, C and F).
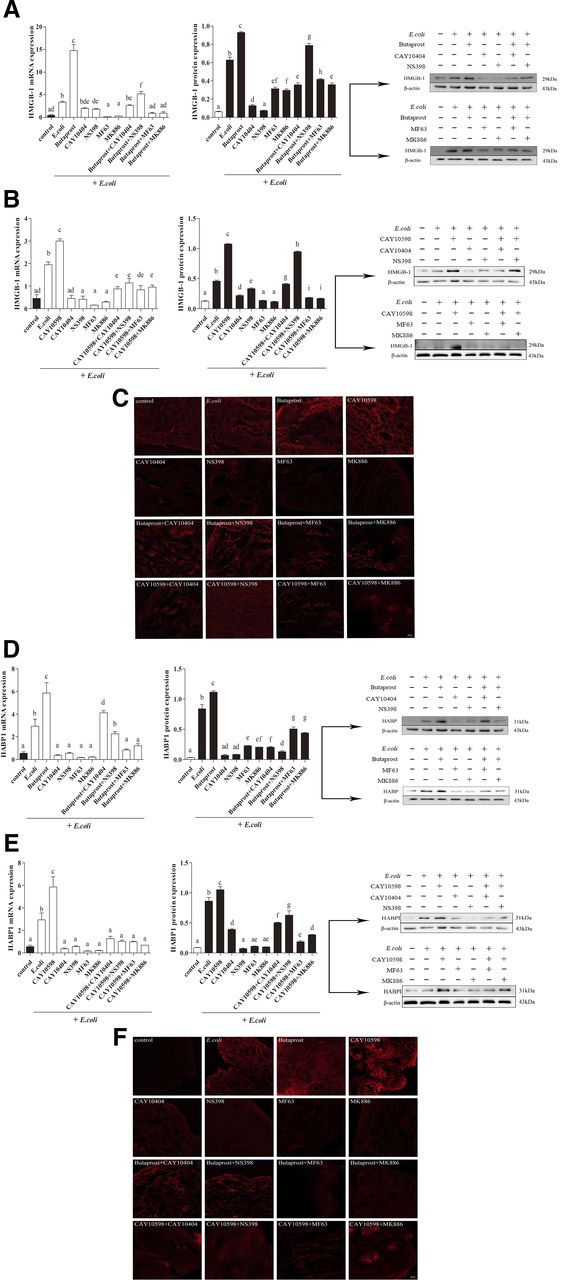
Release of HMGB-1 and HABP1 in bovine endometrial explants infected with E. coli via EP2/4 receptor agonists. The gene and protein expression of HMGB-1 (A and B) and HABP1 (D and E) in explants treated with EP2/4 receptor agonists, COX-2 and mPGES-1 inhibitors, and EP2/4 receptor agonists plus COX-2 and mPGES-1 inhibitors for 12 hours are shown (n = 3; mean ± S.D.). The same letters represent no significant difference; different letters represent significant differences analyzed by GraphPad Prism 5 software (P ≤ 0.05). Immunofluorescence staining of HMGB-1 (C) and HABP1 (F) from bovine endometrial explants are shown as well.
This study also observed pathologic damage of bovine endometrial explants infected with E. coli. In the control group, the structure of endometrium was intact, and the outline of endometrial epithelium, glands, and blood vessels was clear and penetrated the endometrial tissues. Also, the morphology of the cells was clear and the nucleus was round and intact. In contrast with the control group, the epithelial cells of endometrial explants infected with E. coli were partially exfoliated, the glandular cells disintegrated and necrotized, and the outline of the glands was not clear. However, after E. coli treatment, the EP2/4 receptor agonists further strengthened the inflammatory damage of endometrial explants, characterized by complete shedding of epithelial cells and more serious disintegration and fusion of glands. In the COX-2 and mPGES-1 inhibitor groups, the endometrial lesions were alleviated, the outlines of the glands and blood vessels were clear, and epithelial and glandular cells were relatively complete, but were aggravated by the cotreatment with EP2/4 receptor agonists, manifesting as the obscission of epithelial cells, disintegration of glandular cells, and fusion of glands (Fig. 4). These results indicate that the pathologic accumulation of PGE2, through activation of the EP2/4 receptors, increases the expression of damage-associated molecular patterns such as HMGB-1 and HABP1 and promotes inflammatory damage when the bovine endometrium is infected with pathogenic E. coli.

Morphologic observation of tissue injury by the hematoxylin and eosin (H&E) staining technique. Sections of bovine endometrial explants after treatment with EP2/4 receptor agonists, COX-2 and mPGES-1 inhibitors, and EP2/4 receptor agonists plus COX-2 and mPGES-1 inhibitors for 12 hours are shown via H&E staining (original magnification, 100×). Black arrows refer to the endometrial epithelial cells, blue arrows refer to the uterine gland, and red arrows refer to the vessel. All of the samples were infected with E. coli except for the control.
Discussion
Bovine endometritis therapy has made great progress since the arrival of antibiotics. The effects of antibiotics can be weakened by bacterial drug resistance (Zhao et al., 2014). As a result, it is vital to understand the pathogenesis of endometritis in order to search for more efficient NSAIDs and prostaglandin reagents to treat endometritis. Recently, NSAIDs targeting PGE2 synthetase COX-2 and mPGES-1 have been employed to treat bovine endometritis (Chen et al., 2015; Koeberle et al., 2016); finding the NSAIDs inhibiting these two synthases may indeed have potent effects in alleviating bovine endometritis. But unfortunately, these reagent may simultaneously interfere with the homeostatic effects of these mediators downstream from COX-2 and mPGES-1 and potentially block the synthesis of PGE2, a biomolecule that plays an important role in a series of physiologic activities (Ricciotti and FitzGerald, 2011; Nakanishi and Rosenberg, 2013).
The present research explored the inflammatory action of PGE2 in bovine endometrium infected by E. coli that can be signaled by one or more receptors of PGE2. EP2/4 receptor agonists were applied because these two receptors are closely related to inflammation, as initially characterized by coupling to Gαs and increased intracellular cAMP formation, thereby activating a series of downstream events (Regan, 2003; Cheon et al., 2006; Yokoyama et al., 2013). When EP2/4 receptor agonists were added to the explants infected with E. coli, the expression and secretion levels of COX-2, mPGES-1, PGE2, proinflammatory factors, and DAMPs increased enormously. This promotion effect was depressed by adding COX-2 and mPGES-1 inhibitors, indicating that the pathologic accumulation of PGE2 via activation of EP2 and EP4 receptors plays a key role in proinflammatory factors and DAMP expression in the bovine endometrium. In addition, not only the EP2 receptor antagonist but also the EP4 receptor antagonist treatment blocked the enhancement of the EP2 receptor agonist on the expression of IL-1β, IL-6, and TNF-α, which was also observed with the EP4 receptor agonist. Morphologic sections of the treated explants revealed the disappearance of endometrial epithelium and the blurring of glands and vessels when treated with these two agonists; the analysis also demonstrated the alleviation by COX-2 and mPGES-1 inhibitors. These findings jointly resolved the question of the receptors involved in bovine endometritis: both EP2 and EP4 mediate the inflammatory reaction induced by PGE2 in E. coli–infected uteri of bovines.
Under inflammatory conditions, PGE2 is of particular interest because it is involved in many processes leading to the classic signs of inflammation, and COX-2 and mPGES-1 are essential for pathologic biosynthesis of PGE2 (Regan, 2003; Trebino et al., 2003; Jiang and Dingledine, 2013; Kasimanickam et al., 2013; Peter et al., 2015). Thus, this study first examined the level of PGE2 in E. coli–infected bovine endometrial explants. We found that E. coli infection caused the production of PGE2 via COX-2 and mPGES-1, leading to the expression and secretion of inflammatory mediators and DAMPs. This was consistent with the results of other research studies, where IL-1β, IL-6, IL-8, and TNF-α expression was induced by E. coli and lipopolysaccharides in endometrial epithelial cells (Chapwanya et al., 2013; Yao et al., 2017; Li et al., 2019a,c).
After formation via COX-2 and mPGES-1, PGE2 acts locally through binding one or more cognate receptors of its own (Park et al., 2006). The EP1 receptor regulates Ca2+ channel gating via Gαq and the EP3 receptor inhibits adenylate cyclase via Gi. EP2 and EP4 receptors couple to Gs and mediate the increase of cAMP concentration in the cells (Namba et al., 1993; Tabata et al., 2002; Regan, 2003; Cheon et al., 2006; Sugimoto and Narumiya, 2007). In addition, EP2 and EP4 receptors are reportedly involved in a positive feedback loop for PGE2 production in microglia and endometrial tissues (Shie et al., 2005; Li et al., 2019a,b), which is consistent with our study showing that these two receptor agonists increased the expression and secretion of COX-2, mPGES-1, and PGE2 significantly in E. coli–infected endometrial explants. This result indicated that EP2/4 receptors are integral to the pathologic accumulation of PGE2 in E. coli–infected endometrial explants. However, these conclusions are partially contradicted by the results of another investigation, where EP4 was shown to be involved in the feedback loop for PGE2 production in human macrophages infected with M. tuberculosis but EP2 was apparently not involved (Nishimura et al., 2013). This different effect may be explained by the diverse signaling pathways between these two receptors, and may be also related to the selective expression of either of these two receptors in different cells, for example, the action of EP2 during cumulus expansion in ovulation and fertilization (Hizaki et al., 1999) and that of EP4 in closure of the ductus arteriosus (Segi et al., 1998).
Experiments involving colitis and collagen-induced arthritis demonstrated that EP2/4 receptor agonists, but not those for EP1/3 receptors, induced IL-6, IL-1β, and IL-23 expression and caused T cells to differentiate into T helper cell 17 effectors (Honda et al., 2006; Sheibanie et al., 2007a,b), suggesting that EP2/4 receptors play a key role in the release of proinflammatory factors. However, there is no evidence for the effect of EP2/4 receptors in E. coli–infected bovine endometrial explants, especially regarding the secretion of inflammatory factors. Our results were consistent with the aforementioned findings, showing that in the process of inflammation via E. coli infection, EP2/4 receptor agonists significantly induced the expression of proinflammatory factors, establishing the essential role of EP2/4 receptors in the endometrial inflammatory action caused by E. coli.
Another important feature of inflammation, besides the release of inflammatory factors, is tissue inflammatory damage. Our study evaluated the production of two DAMPs, HMGB-1 and HABP1. Previous studies have observed the redundant distribution of these two factors in the inflammation of human endometrial and intestinal tissues (de la Motte and Drazba, 2011; Yun et al., 2016). We found that EP2/4 receptors were responsible for this redundant distribution of HMGB-1 and HABP1 in E. coli–infected endometrial explants, indicating that the endometrial explant damage caused by E. coli is closely related to PGE2-EP2/4.
Tsuchiya et al. (2003) obtained several consensus sequences relevant to inflammatory stimuli, such as those for nuclear factor (NF) IL-6 (NF-IL6), NF-κB, and activator protein-2 (Tsuchiya et al., 2003). Among the mutual promoters of EP2 and EP4 receptors, NF-κB is known to be activated rapidly in response to proinflammatory cytokines, such as IL-1, resulting in regulation of immune responses including the upregulation of EP2 and EP4 receptors in inflammatory diseases (Li and Verma, 2002). Considering the mutual promoters of EP2 and EP4 receptors, we supposed that these two receptors may act together to mediate the inflammation resulting from E. coli. Interestingly, the results of the present study are consistent with our hypothesis, thus supporting the conclusion that the joint action of EP2/4 receptors promoted the inflammation of endometrial explants in dairy bovines infected with E. coli.
Our data have demonstrated the promoting effect of PGE2 through EP2/4 receptors in E. coli–infected endometrial inflammation of bovines. We believe that these findings are of significant therapeutic relevance, and that EP2/4 receptors could serve as better intervention targets than cyclooxygenase or prostaglandin E for adjunctive treatment of bovine endometritis. However, blocking EP2/4 receptors may also influence many other physiologic and pathologic events downstream from these two receptors, including ovulation and fertilization, bone formation, and closure of the ductus arteriosus (Sugimoto and Narumiya, 2007). Therefore, a specific signaling pathway downstream from EP2/4 receptors responsible for the endometrial inflammation of bovines caused by E. coli needs further investigation. Considering that EP4/adenylate cyclase/PKA has been shown to be involved in inflammation due to E. coli infection (Li et al., 2019a,b), other pathways related to inflammation, such as EP2/PKA/CREB response element binding protein (Fujino et al., 2002; Jiang and Dingledine, 2013) and EP4/phosphatidylinositol 3-kinase/ERK (Namba et al., 1993; Pozzi et al., 2004), need more focus.
In this study, we investigated the EP2/4 receptors of PGE2 that were possibly involved in E. coli–infected explants of the endometrium of dairy bovines. The results further suggested that accumulation of PGE2 might play an essential role in tissue damage of endometrial explants infected with E. coli via promoting proinflammatory factor IL-1β, IL-6, IL-8, TNF-α, HMGB-1, and HABP1 production. EP2/4 receptors are involved in a positive feedback loop for COX-2 and mPGES-1 expression, thus enhancing PGE2 production. Furthermore, these two receptors mediate the proinflammatory action of PGE2 by increasing proinflammatory factors and DAMP secretion and aggravating damage of explants.
In summary, the present study examined the receptors responsible for the proinflammatory action of PGE2 in bovine endometrial inflammation caused by E. coli. We concluded that EP2/4 receptors work together to mediate the pathologic accumulation of PGE2 and bovine endometrial inflammatory damage in E. coli–infected endometrial explants, thus providing the possibility of developing new NSAIDs against EP2/4 receptors for treatment of endometritis in dairy bovines.
Acknowledgments
The skillful technical assistance of Dr. Chenguang Du and colleagues with confocal laser scanning microscopy is greatly appreciated.
Authorship Contributions
Participated in research design: Zhang, Wang, Li, Liu, Cao.
Conducted experiments: Zhang, Wang, Li.
Contributed new reagents or analytic tools: Mao, Liu, Cao.
Performed data analysis: Zhang, Wang.
Writers contributed to the writing of the manuscript: Zhang, Wang, Li, Cao.
Footnotes
- Received September 5, 2019.
- Accepted November 11, 2019.
↵1 C.Z., L.W., and T.L. contributed equally to this work.
This work was supported by the National Natural Science Foundation of China [Grant 31672603].
Abbreviations
- cAMP
- cyclic 3,5-adenosine monophosphate
- COX-2
- cyclooxygenase-2
- DAMP
- damage-associated molecular pattern
- HABP1
- hyaluronan binding protein 1
- HMGB-1
- high mobility group box-1
- IL
- interleukin
- mPGES-1
- microsomal prostaglandin E synthase-1
- NF
- nuclear factor
- NSAID
- nonsteroidal anti-inflammatory drug
- PGE2
- prostaglandin E2
- PKA
- protein kinase A
- RT-PCR
- reverse-transcription polymerase chain reaction
- TNF-α
- tumor necrosis factor-α
- Copyright © 2020 by The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}