


Abstract
Multidrug resistance protein (MRP) 3 transports bile salts and conjugated xenobiotics from cells (hepatocytes and enterocytes) into the blood. Hepatic MRP3 expression is low under normal conditions but is markedly up-regulated during cholestasis. Since little is known about additional factors increasing human hepatic MRP3 expression, we investigated the variability of MRP3 expression in a large collection of human livers and factors contributing to variable MRP3 expression in liver and HepG2 cells. MRP3 was measured in 62 human livers from patients with and without omeprazole treatment and in HepG2 cells with and without omeprazole or β-naphthoflavone treatment. Livers of patients treated with omeprazole showed 4.8-fold (P< 0.0001) higher MRP3 protein expression compared with the remainder of the population. Accordingly, MRP3 mRNA and protein were induced 2.4- and 1.8-fold, respectively (P < 0.01 andP < 0.05), in HepG2 cells treated with omeprazole. Finally, MRP3 was induced in HepG2 cells by β-naphthoflavone. In summary, treatment with omeprazole and β-naphthoflavone is a determinant of variable human hepatic MRP3 expression.
It is increasingly recognized that active uptake and efflux processes are important determinants for disposition of endogenous compounds (e.g., bile salts) and drugs (e.g., cyclosporine and HIV protease inhibitors) (Kusuhara et al., 1998; Trauner et al., 1998; Borst et al., 2000;Fromm, 2000; Keppler and König, 2000). MRP3 is a member of the multidrug resistance protein (MRP) family (for review, see Borst et al., 2000; Keppler and König, 2000). These membrane proteins mediate ATP-dependent efflux of lipophilic substances conjugated to glucuronate, glutathione, and sulfate. Substrates of the basolateral export pump MRP3 are sulfated and nonsulfated bile salts, 17β-glucuronosyl estradiol, leukotriene C4, and several anticancer drugs (König et al., 1999a; Zeng et al., 1999; Hirohashi et al., 2000; Zelcer et al., 2001). MRP3 appears to be involved in bile salt homeostasis in several ways. First, expression of MRP3 in the basolateral membrane of enterocytes indicates that MRP3 could be involved in intestinal transport of organic anions from the cells into portal blood (Rost et al., 2002). Second, MRP3 expression was found in the basolateral membrane of human hepatocytes, with exceptionally high expression in livers from patients with Dubin-Johnson syndrome and a patient with primary biliary cirrhosis, in the lateral membrane of intrahepatic bile duct epithelial cells (cholangiocytes), and in the basolateral membrane of human gallbladder epithelia (König et al., 1999b; Kool et al., 1999; Gotoh et al., 2000; Rost et al., 2001). Since in animal models of cholestasis, Mrp2 is down-regulated whereas MRP3 expression is induced (Hirohashi et al., 1998; Ogawa et al., 2000;Donner and Keppler, 2001), it appears that MRP3 compensates for impaired MRP2 function and responds to bile salts at the transcriptional level (Schuetz et al., 2001). Accordingly, recent data indicate that intestinal MRP3 is also induced by bile acids (Inokuchi et al., 2001).
In addition to increased MRP3 expression in humans during cholestasis, there is little information on the variability of MRP3 expression in human livers and on factors contributing to MRP3 expression in liver, such as comedication (e.g., by the proton pump inhibitor omeprazole), age, gender, and smoking. We therefore investigated various aspects of MRP3 expression and regulation in 62 human livers and HepG2 cells.
Materials and Methods
Tissue Collection.
Human liver tissue (n = 62) was obtained as nontumorous tissue surrounding surgically removed liver tumors or metastases or material that was surgically resected for other reasons. Extensive documentation was obtained for each of the samples, including demographic data of the patient (age: mean ± S.D., 56.1 ± 16.5 years; range: 4–85 years; and sex: 31 male and 31 female), diagnosis, smoking status (11 smokers, 49 nonsmokers, and 2 unknown), and drug intake. The tissues were stored at −80°C until nuclear/membrane pellets were prepared according to the following standard procedure. One gram of tissue was homogenized in buffer containing 1 mM EDTA, 1 mM dithiothreitol, 10 mM HEPES pH 7.4, 0.2 mM Pefabloc (Roth, Karlsruhe, Germany), and 0.15 mM KCl and centrifuged at 15,000g (4°C for 30 min). The formed nuclear/membrane pellet was washed once with buffer, resuspended in an appropriate quantity of buffer containing 1 mM EDTA, 1 mM Pefabloc, 100 mM Tris, 1 μg/ml leupeptin, and 1 μg/ml pepstatin and stored at −80°C. Protein concentration was measured according to the method ofSmith et al. (1985). The study was approved by the ethics committees of the Medical Faculties of the Charité, Humboldt-University Berlin and the University of Tübingen, and written informed consent was obtained from each patient.
MRP3 mRNA in Human Liver.
High-quality RNA was obtained from a small piece of liver (maximal side length, 5 mm), which was transferred into a vial containing 1 ml of RNAlater (Ambion, Austin, TX). The samples were first kept at 4°C for 24 h to promote inactivation of RNases by the reagent and then stored at −80°C for maximally 2 months until RNA was prepared. Total RNA from tissue homogenates was isolated using RNeasy columns (QIAGEN, Hilden, Germany) according to the manufacturer's instructions. All RNAs were treated with DNase I using QIAGEN's on-column DNase digestion kit. The isolated RNA was reverse transcribed using the TaqMan Reverse Transcription Reagents (Applied Biosystems, Foster City, CA) according to the manufacturer's instructions.
Real-time quantitative PCR was performed with the ABI PRISM 7700 Sequence Detection System (Applied Biosystems) by applying the 5′-nuclease assay with TaqMan probes. PCR reactions were carried out in a 25-μl volume containing cDNA corresponding to 40 (human tissue) or 50 ng (HepG2 cells) of total RNA, 1× TaqMan Universal PCR Master Mix (Applied Biosystems), 300 nM forward primer, 300 nM reverse primer, and 200 nM TaqMan probe. The forward primer (5′-TGATTCCACTCAACGGAGCT-3′) was positioned in exon 11 of the MRP3 mRNA sequence from the GenBank database (accession no. AF085690) and the reverse primer (5′-TGATGCGCGAGTCCTTCA-3′) in exon 12. The TaqMan probe (5′-CGCGCCTTCCAGGTAAAGCAAATG-3′) was labeled with the reporter FAM (6-carboxyfluorescein) at the 5′-end and with the quencher TAMRA (6-carboxytetramethyl-rhodamine at the 3′-end and was designed to contain sequences of both exons (11 and 12). Cycling conditions were 2 min at 50°C followed by 10 min at 95°C, 40 cycles of 15 s at 95°C, and 1 min at 60°C. Standard curves for this assay were calculated by using serial dilutions of known amounts of linearized MRP3 plasmid cDNA (provided by Dr. R. B. Kim, Vanderbilt University, Nashville, TN). The results for MRP3 mRNA were normalized to β-actin as housekeeping gene. For β-actin detection, 1:50 dilutions of the cDNA liver samples were analyzed using the TaqMan human endogenous control reagents for human β-actin (Applied Biosystems) under the above-mentioned cycling conditions. Standard curves for β-actin were calculated by using serial dilutions of known amounts of total liver cDNA. Triplicate measurements were performed for all samples.
Histological Analysis of Liver Samples.
Frozen liver samples were thawed and fixed in 10% neutral buffered formalin solution (Sigma Chemie, Deisenhofen, Germany) and treated with paraffin in a Shandon Pathcentre according to the manufacturer's recommended procedure (Thermo Shandon GmbH, Darmstadt, Germany). Paraffin sections were hematoxylin-eosin or van Gieson (Lillie and Geer, 1965) stained and analyzed by standard procedures for histological classification of liver diseases on a Diaplan microscope (Leica, Wetzlar, Germany) with Leitz (Wetzlar, Germany) optic.
MRP3 Protein in Human Livers.
Fifty micrograms of total protein from nuclear/membrane pellet fractions were incubated in sample buffer at 37°C for 30 min before separation on 5% stacking and 7.5% separating SDS-polyacrylamide gels. Transfer of protein to nitrocellulose membranes was performed using a tank-blotting system (Bio-Rad, Munich, Germany). Nonspecific binding sites were saturated by incubation with 5% nonfat dry milk in 25 mM Tris/HCl, 137 mM NaCl, and 2.7 mM KCl, pH 7.4 (TBS) containing 0.1% (v/v) Tween 20 (TBS-T) for 1 h. Primary antibodies anti-MRP3 (human) Mab (M3II-9) (Alexis, Grünberg, Germany), as well as anti-β-actin clone AC-15 (Sigma-Aldrich, St. Louis, MO), were diluted according to manufacturer's instructions in 1% nonfat dry milk in TBS-T, and membranes were incubated for 1 h with this solution. After washing with TBS-T, membranes were incubated in 1% nonfat dry milk in TBS-T with peroxidase-conjugated goat anti-mouse IgG (Oncogene, Cambridge, MA) for 60 min and finally washed with TBS-T. Immunoreactive proteins were visualized by chemiluminescence detection solution (SuperSignal West Dura Extended Duration Substrate; Pierce, St. Augustin, Germany). Specificity of the MRP3 antibody was tested using MRP1, -2, and -3 overexpressing cells (provided by Professor P. Borst of The Netherlands Cancer Institute, Amsterdam, The Netherlands). Chemiluminescence of the two MRP3 bands described previously (König et al., 1999b) was measured with a charge-coupled device camera (Fuji LAS-1000; Raytest, Straubenhardt, Germany) and analyzed using AIDA 2.31 software (Raytest). Serial dilutions of known amounts of a total standard liver protein were coanalyzed on each gel as a calibration curve.
Effect of Omeprazole on MRP3 Expression in HepG2 Cells.
HepG2 cells were grown in Dulbecco's modified Eagle's medium containing 1% penicillin/streptomycin and 10% fetal calf serum. Cells were plated in six-well plates (500,000 cells/well). Twenty-four hours after seeding of cells, they were incubated with 10 or 100 μM omeprazole (kindly provided by AB Hässle, Mölndal, Sweden) in Dulbecco's modified Eagle's medium containing 0.1% DMSO (final concentration in medium), 1% penicillin/streptomycin, and 10% dextran-coated charcoal-treated fetal calf serum. Incubation was carried out for 48 h, and during this time, the incubation medium was changed once. After the incubation time, total RNA was isolated as described above. For the isolation of protein, cells were harvested and homogenized in lysis buffer (containing 10 mM Tris/HCl and 1 mM Pefabloc) by sonification.
Effect of β-Naphthoflavone on MRP3 Expression in HepG2 Cells.
For further investigation of a possible induction mechanism of MRP3 via the Ah receptor, we incubated HepG2 cells with the Ah receptor ligand β-naphthoflavone. Incubation studies were carried out as described above using β-naphthoflavone concentrations of 5 and 10 μM (final DMSO concentration, 1%). As a positive control, we determined in these cells the induction of CYP1A activity by β-naphthoflavone. Accordingly, cells were treated for 48 h with vehicle alone and 5 and 10 μM β-naphthoflavone. Thereafter, these cells were incubated with 25 μM of the CYP1A substrate ethoxyresorufine. Aliquots of the incubation medium were taken at 0, 1.5, and 3 h and analyzed for formation of the ethoxyresorufine metabolite resorufine. Concentrations of resorufine were measured by a high-performance liquid chromatography fluorescence detection system (extinction, 522 nm and emission, 586 nm).
Statistical Analysis.
Results are presented as mean ± S.D. Differences in MRP3 expression were tested for significance by unpaired t tests (e.g., with and without omeprazole treatment). Differences between mean values obtained for the different concentrations of omeprazole in in vitro incubation experiments were analyzed for statistical significance by analysis of variance with subsequent Student-Newman-Keuls tests.
Results
Histological Analysis of Human Livers.
Histological analysis revealed that the samples were noncirrhotic and nontumorous. Since it has been reported that cholestasis results in a pronounced up-regulation of MRP3 in liver, we particularly examined the samples for any signs of local cholestasis. However, none of the livers investigated revealed signs of local cholestasis.
MRP3 mRNA in Human Liver.
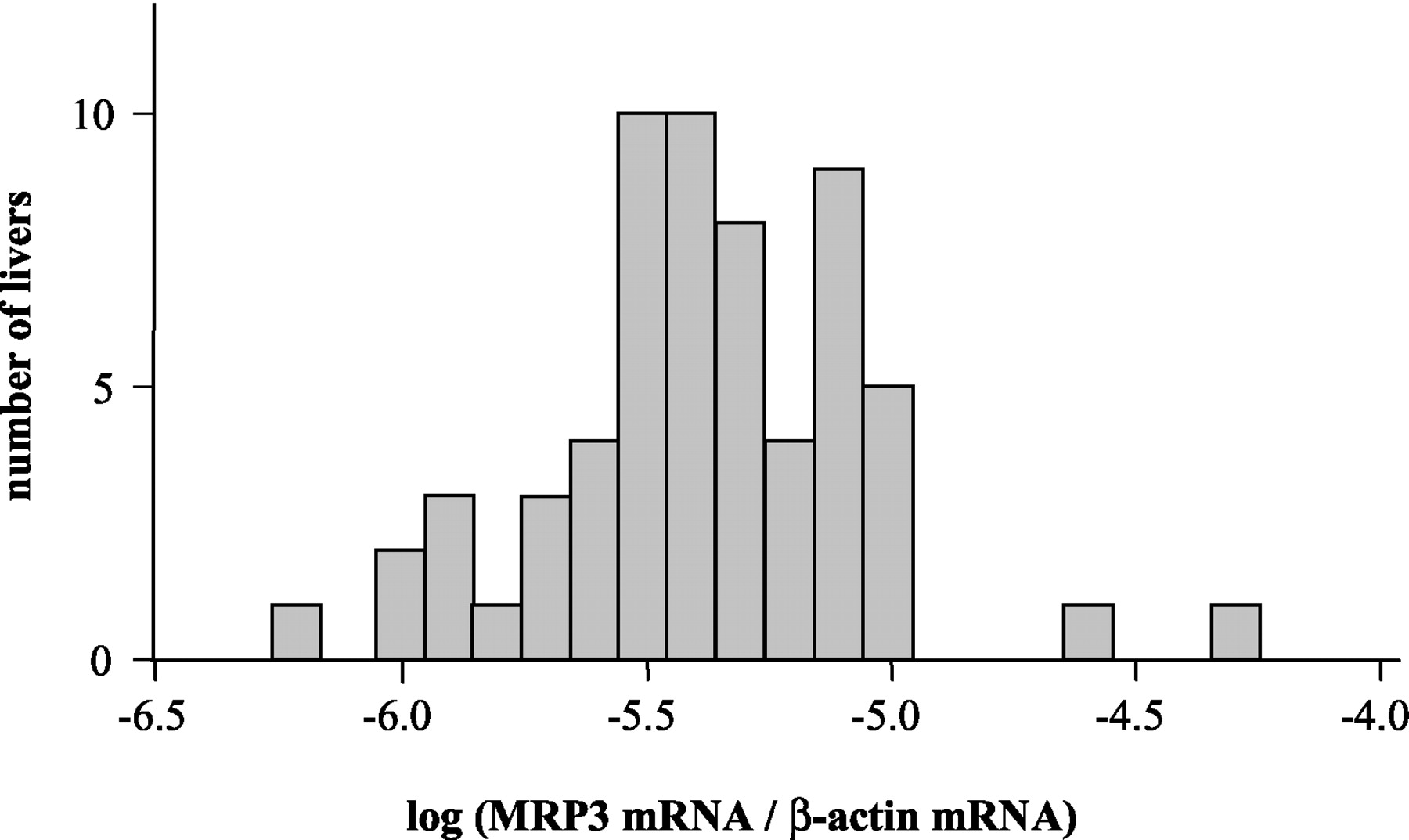
We analyzed MRP3 mRNA in 62 human livers by TaqMan real-time RT-PCR and found a MRP3 mRNA distribution as indicated in Fig. 1. The expression of MRP3 mRNA in human liver was highly variable and varied by a factor of 86. MRP3 mRNA was not significantly affected by age, gender, smoking, or drug treatment (e.g., omeprazole).

MRP3 mRNA expression in 62 human livers determined by real-time TaqMan RT-PCR and normalized for β-actin mRNA.
MRP3 Protein in Human Liver.
A representative Western blot for MRP3 in human livers is shown in Fig. 2. We observed the previously described pattern of two bands, a possible result of different levels of glycosylation (König et al., 1999b). Western blot analysis of MRP3 protein in 62 human livers showed a highly variable protein expression as indicated in Fig.3. All samples had constant β-actin protein levels (Fig. 2). No MRP3 protein was detectable in two livers (with log MRP3 mRNA/β-actin being −5.7 and −5.2; see Fig. 1). In the remainder of the population, MRP3 expression varied 193-fold. No correlation was found between expression of MRP3 protein and MRP3 mRNA levels in human liver. In addition, we analyzed whether medication has an influence on MRP3 protein levels. We found significant differences in MRP3 protein levels of subjects treated with omeprazole compared with untreated subjects (Fig. 3). Mean MRP3 protein expression of 10 subjects treated with omeprazole is 152.1 ± 117.0 (range, 0–413.1) compared with 31.8 ± 35.1 a.u./μg of total protein (P < 0.0001; range, 0–175.4) without omeprazole. Moreover, smokers had a (nonsignificant) trend toward higher MRP3 protein expression in comparison with nonsmokers [n = 7 versus 43 (omeprazole-treated patients were excluded) and 43.1 ± 61.5 versus 30.1 ± 30.4 a.u./μg of total protein, not significant]. Gender, age, and any other comedication (e.g., benzodiazepines) did not significantly affect MRP3 protein in human livers.

MRP3 protein expression in human livers. A representative Western blot shows variable MRP3 expression in seven nucleus/membrane fractions of human livers and constant β-actin levels. Fifty micrograms of protein from each sample were analyzed and quantified relative to a dilution series of a standard liver sample (75, 50, 25, and 10 μg of protein in lanes A, B, C, and D, respectively).

MRP3 protein expression in 62 human livers. Fifty micrograms of nucleus/membrane fractions of each human liver were analyzed by Western blot using a dilution series of a standard liver sample for quantification. Each arrow (in the insert, ∗) represents one subject treated with omeprazole. Two additional subjects are not shown in the log-transformed part of the figure since they did not have detectable MRP3 protein expression. One of these two individuals was also treated with omeprazole. The insert displays the distribution of the linear expression data.
Effect of Omeprazole on MRP3 Expression in HepG2 Cells.
To investigate the potential molecular basis for the increase in hepatic MRP3 in omeprazole-treated patients, we incubated HepG2 cells with omeprazole. MRP3 mRNA levels were 1.04 ± 0.36, 1.14 ± 0.61, and 2.46 ± 0.54 a.u. in control, 10, and 100 μM omeprazole-treated cells, respectively (control versus 100 μM omeprazole, P < 0.01; Fig.4). Moreover, MRP3 protein levels in control, 10, and 100 μM omeprazole-treated cells were 4.5 ± 0.7, 6.3 ± 2.3, and 8.2 ± 2.7 a.u./μg of total protein (Fig. 5; control versus 100 μM omeprazole, P < 0.05), respectively.

Induction of MRP3 mRNA by omeprazole in in vitro studies. HepG2 cells were incubated with vehicle (0.1% DMSO), 10, and 100 μM omeprazole for 48 h. MRP3 mRNA was analyzed by real-time TaqMan RT-PCR. Results obtained for MRP3 mRNA were normalized for β-actin mRNA (∗, P < 0.05; ∗∗,P < 0.01).

Induction of MRP3 protein by omeprazole in in vitro studies. After 48 h of incubation of HepG2 cells with vehicle (0.1% DMSO), 10, and 100 μM omeprazole, cells were homogenized and analyzed for MRP3 protein by Western blot. Fifty micrograms of cell homogenate were analyzed for MRP3 and quantified using a dilution series of a standard liver sample. A, MRP3 Western blot (7.5 μg of protein were loaded from the standard liver); B, mean MRP3 protein expression from duplicate measurements of three independent experiments (∗, P < 0.05).
To investigate the potential involvement of the Ah receptor pathway in MRP3 induction, we incubated HepG2 cells with 5 and 10 μM β-naphthoflavone, which is known to be a ligand for this receptor. MRP3 mRNA levels were 1.78 ± 0.86, 5.47 ± 0.94, and 5.81 ± 0.29 a.u. in control, 5, and 10 μM β-naphthoflavone-treated cells, respectively. Cells, which were treated with 5 and 10 μM β-naphthoflavone, showed on average a 10.5- and 8.5-fold higher resorufine formation in comparison to control cells, respectively (Fig. 6).

Induction of MRP3 mRNA and CYP1A activity in HepG2 cells by β-naphthoflavone. Cells were incubated with vehicle (1% DMSO) and 5 and 10 μM β-naphthoflavone for 48 h. After the incubation period, MRP3 mRNA was analyzed by real-time TaqMan RT-PCR and normalized to β-actin (A). CYP1A activity in these cells was characterized by the CYP1A substrate ethoxyresorufine (B). The metabolite resorufine was measured by a high-performance liquid chromatography fluorescence detection system after 1.5 and 3 h of incubation of HepG2 cells with 25 μM ethoxyresorufine.
Discussion
Our study in a large panel of human liver tissues revealed a considerable variability of MRP3 expression. Therapy with the proton pump inhibitor omeprazole was identified as an environmental factor causing increased hepatic MRP3 expression. Additional in vitro studies indicate activation of the Ah receptor pathway as a putative mechanism of hepatic MRP3 induction by omeprazole. The potential clinical relevance of these observations arises from the fact that MRP3 is involved in transport of endogenous compounds (e.g., bile salts) as well as elimination of multiple drugs or drug metabolites (e.g., anticancer agents and etoposide glucuronide) (Kusuhara et al., 1998;Trauner et al., 1998; Borst et al., 2000; Fromm, 2000; Keppler and König, 2000). Thus, interindividual differences in hepatic MRP3 expression are likely to influence bile salt homeostasis, drug elimination, and toxicity.
This link between transporter function and drug disposition and effects is now well understood for the MDR1 gene product P-glycoprotein (Fromm, 2000; Matheny et al., 2001). P-glycoprotein determines disposition of many commonly used drugs (e.g., cyclosporine, HIV protease inhibitors, and cardiac drugs). Moreover, induction (e.g., by rifampin) and inhibition of P-glycoprotein have been identified as mechanisms underlying drug interactions in humans (Fromm et al., 1999;Greiner et al., 1999; Westphal et al., 2000). In addition,MDR1 single nucleotide polymorphisms are associated with altered P-glycoprotein expression and function (Hoffmeyer et al., 2000;Hitzl et al., 2001; Kim et al., 2001) and determine the response of an individual to drug treatment (e.g., HIV) (Fellay et al., 2002). It is currently unknown whether genetic mutations contribute to variable MRP3 expression in humans, but for example, DNA from the only two patients with no detectable MRP3 might be very useful for mutation screening.
Using a large collection of human liver tissues, we identified omeprazole as an inducer of MRP3 expression. This effect could be reproduced with HepG2 hepatoma cells, which showed a concentration-dependent induction of MRP3 expression by omeprazole. Interestingly, we observed increased MRP3 mRNA and protein levels in HepG2 cells but only increased protein levels (without increase of MRP3 mRNA) in our collection of human livers. The underlying mechanism of these differences is unclear at the moment, but similar observations have been made during MRP3 induction studies in rats (Ogawa et al., 2000). It should be noted that we collected only nontumorous and nonpathologic human liver tissue for this study. We cannot, however, completely rule out that the vicinity of the collected tissue to a tumor or local inflammation affected MRP3 expression to some extent.
Although it was not the goal of this work to investigate the molecular mechanism of MRP3 induction, the following data indicate that activation of the Ah receptor pathway could be one mechanism involved in MRP3 induction by omeprazole. First, activation of the Ah receptor pathway mediates CYP1A1 induction by omeprazole (Daujat et al., 1992;Backlund et al., 1997; Dzeletovic et al., 1997). Second, dioxin-responsive elements to which the Ah receptor/Arnt heterodimer may bind have been identified in the 5′-flanking region of human MRP3 (Takada et al., 2000). Third, in vitro data indicate that MRP3 expression is also induced by β-naphthoflavone (unpublished data) and 2-acetylaminofluorene (Stöckel et al., 2000; Schrenk et al., 2001), which are known activators of the Ah receptor pathway. Finally, our own data indicate that smokers have on average 1.4-fold higher MRP3 protein levels in comparison to nonsmokers. Despite the low MRP3 expression in human liver, we identified a commonly used drug as inducer of hepatic MRP3 expression and provide evidence for involvement of the Ah receptor pathway in omeprazole-mediated MRP3 induction.
Taken together, using in vitro studies in combination with clinical samples, induction of an ABC transporter involved in homeostasis of endogenous compounds as well as in drug disposition was found. Moreover, evidence is provided for the putative mechanism of MRP3 induction. Further studies are required to determine the impact of highly variable MRP3 expression on disposition of endogenous compounds and drugs and the contribution of genetic polymorphisms to variable MRP3 expression.
Acknowledgments
We thank Dr. P. Borst (The Netherlands Cancer Institute) for kindly providing the MRP1, -2, and -3 overexpressing cells and Dr. L. Becquement for valuable suggestions.
Footnotes
-
Supported by the Deutsche Forschungsgemeinschaft (Bonn, FR 1298/2-1), Grant 01 GG 9846 from the German Federal Ministry of Education and Science, and the Robert Bosch Foundation (Stuttgart, Germany).
-
DOI: 10.1124/jpet.102.043547
- Abbreviations:
- HIV
- human immunodeficiency virus
- MRP
- multidrug resistance protein
- PCR
- polymerase chain reaction
- TBS
- Tris-buffered saline
- TBS-T
- Tris-buffered saline/Tween 20
- DMSO
- dimethyl sulfoxide
- RT-PCR
- reverse transcription polymerase chain reaction
- a.u.
- arbitrary units
- Received August 27, 2002.
- Accepted October 3, 2002.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}